
Abstract
Aims:
Sub-clinical inflammation during pre-diabetes is one of the predisposing factors that facilitates the progression of pre-diabetes to type 2 diabetes. The administration of oleanolic acid (OA) with or without dietary intervention ameliorates the metabolic and cardiovascular complications in diet-induced pre-diabetes animal models of pre-diabetes. This study aimed to investigate whether OA can also suppress immune activation and ameliorate pro-inflammatory markers.
Methods:
Pre-diabetes was induced by feeding Sprague Dawley rats a high-fat high carbohydrate diet for 20 weeks. The pre-diabetic rats were then treated with OA (80 mg/kg) or metformin (500 mg/kg) in the presence or absence of dietary interventions for a period of 12 weeks. At the end of the treatment period, the animals were euthanised and whole blood was used for platelet and immune cell count while plasma was used for fibrinogen, cluster differentiation 40 ligand and pro-inflammatory cytokine evaluation.
Results:
The results of this study revealed that OA, with or without dietary intervention, improved lipid metabolism by restoring high-density lipoprotein (HDL) and low-density lipoproteins (LDLs) as well as reducing platelets and immune cell counts. Furthermore, OA also decreased plasma proinflammatory cytokines, including tumour necrosis factor-α and -1β. Markers of immune activation such as C-reactive protein, fibrinogen, and CD40L were also decreased upon administration of OA with or without dietary intervention.
Conclusion:
The findings of this study suggest that OA may provide an alternative to prevent the progression of pre-diabetes to overt diabetes. This was evident by the reduction of differential white blood cell count and proinflammatory cytokines that exercebate insulin resistance. However, more studies are needed to elucidate the molecular mechanisms and to improve efficacy.
Keywords
Introduction
Innate immunity activation and chronic subclinical inflammation are some of the predisposing risk factors that precede the onset of diabetes in individuals that have been previously diagnosed with pre-diabetes. 1 According to the World Health Organisation, pre-diabetes is a long-lasting state of plasma glucose levels above normal but not high enough for the diagnosis of type 2 diabetes mellitus (T2DM). Sedentary lifestyles, genetic predisposition and chronic consumption of high caloric diets are among the causes of the development of pre-diabetes. 2 Obesity has also strongly correlated with the incidence of pre-diabetes and studies show that it is one of the risk factors for pre-diabetes. 3 Obesity is associated with metabolic complications including insulin resistance, hyperlipidaemia, hypercholesterolaemia and moderate hyperglycaemia that are implicated in innate immunity activation and low-grade inflammation. 4 The elevation of circulating lipid metabolites including triglycerides, free fatty acids, cholesterol and lipoprotein-cholesterols is implicated in innate immune system activation. 5 In addition, obesity and insulin resistance are also associated with the leakage of lipopolysaccharides (LPS) from the intestine into the circulation. 6 LPS is implicated in the activation of innate immunity through activation of pattern recognition receptors (PRR) such as Toll-like receptor 4 (TLR4) on the membrane surface of immune cells such as neutrophils and macrophages. 7 The binding of LPS to immune cells leads to their activation and secretion from these cells of pro-inflammatory cytokines such as interleukin-1β (IL-1β) and tumour necrosis factor-alpha (TNF-α). 8 Moreover, low-density lipoprotein (LDL) can also bind to the TLRs on the surface of resident macrophages resulting in the secretion of pro-inflammatory cytokines and lipid accumulation. Oxidised LDL can bind to CD36 on the platelet surface leading to platelet activation. 9 The activation of platelets has been reported to result in the formation of platelets–leukocytes aggregation with neutrophils and monocytes, as well as releasing cluster differentiation 40 ligand (CD40-L), which is another inflammatory mediator. 10 Moreover, Mzimela et al. reported that a diet-induced pre-diabetic rat model presents immuno-metabolic derangements including elevation of fibrinogen and lymphocytes, which worsen the progression from pre-diabetes to overt diabetes. 11 The pharmacological management of pre-diabetes includes the use of insulin-sensitising agents, e.g., biguanides such as metformin (MET). 12 These agents are associated with suppression of inflammation through the inhibition of oxidative stress but are said to be therapeutic only when used in conjunction with lifestyle modifications. 13 However, lifestyle modifications such as caloric restriction and increased physical activity are often neglected by patients. 14 Therefore, this calls for more research to establish strategies for preventing the development of low-grade inflammation while sensitising cells for insulin and subsequently preventing the progression of pre-diabetes to overt diabetes. 15 Plant-derived bioactive compounds, including triterpenes and flavonoids, have been shown to possess the anti-diabetic properties. 16 In our laboratory, oleanolic acid (OA), a plant-derived triterpene, has been reported to ameliorate the predisposing risk factors of the progression of pre-diabetes to diabetes, including impaired insulin transduction and hyperlipidaemia. 17 Furthermore, OA has been also found to ameliorate markers of cardiovascular disease and non-alcoholic fatty liver disease in a diet-induced pre-diabetes rat model.17,18 However, innate immunity activation, as well as chronic subclinical inflammation, is also associated with the progression of pre-diabetes to diabetes. Hence, this study sought to evaluate whether OA, with and without dietary intervention, can reduce the markers of innate immunity activation and chronic subclinical inflammation in a diet-induced pre-diabetic rat model.
Methods and materials
Drugs and chemicals
High-fat high-carbohydrate (HFHC) diet [AVI Products (Pty) Ltd., Waterfall, South Africa]; dimethyl sulphoxide (10% DMSO), phosphate-buffered saline (PBS), MET (Sigma-Aldrich, St Louis, MO, USA); ethanol, isofor, liquid nitrogen [Safeline Pharmaceuticals (Pty) Ltd., Roodeport, South Africa]; concentrated biotinylated detection antibody, biotinylated detection antibody detection diluent substrate reagent, stop solution, concentrate wash buffer, standards micro enzyme-linked immunosorbent assay (ELISA) plate, reference standard, concentrated avadin-horseradish peroxide (HRP), HRP diluent (Elabscience product purchased from Biocom Africa, Centurion, South Africa). All chemicals and reagents were sourced from standard pharmaceutical suppliers and were of analytical grade.
Extraction method
OA was extracted from Syzygium aromaticum [(Linnaeus) Merrill & Perry] [Myrcene] (cloves) using an established protocol from Khathi et al. 19
Animal studies
Male Sprague-Dawley rats (130–160 g) used in this study were bred and housed in the Biomedical Research Unit of the University of KwaZulu-Natal. The animals were maintained under standard laboratory conditions of constant temperature (22 ± 2°C), CO2 content (<5000 p.m.), relative humidity (55 ± 5%) and illumination (12 h light/dark cycle, lights on at 0700 hours). The noise level was maintained at less than 65 decibels. The animals were allowed access to food and fluids ad libitum. The Animal Research Ethics Committee approved all animal procedures and housing conditions of the University of KwaZulu-Natal (ethics no. AREC/035/016M). The animals acclimatised to their new environment for 1 week while consuming standard rat chow and tap water before exposure to a well-established experimental HFHC diet. 18
Experimental design
Experimental pre-diabetes was induced in male Sprague-Dawley rats (n = 36) using a previously described protocol. The pre-diabetic animals were further sub-divided into six groups, each group having six rats (n = 6). The groups were as follows: pre-diabetic control group (PC) were the pre-diabetic animals that continued with the experimental diet throughout the study period; metformin group (MET) were the pre-diabetic animals that continued with the experimental diet but received MET during treatment period; MET and dietary intervention group (MET+DI) were the pre-diabetic animals that changed to a standard diet and received MET during treatment period; oleanolic acid group (OA) were the pre-diabetic animals that continued with experimental diet but received OA during the treatment period as well as OA and dietary intervention group (OA+DI) were the pre-diabetic animals that changed to a standard diet and received OA during experimental period. Animals that served as non-pre-diabetic controls (NPC) were those that were fed standard rat diet and diagnosed as without pre-diabetes. The dietary intervention refers to changing animals from the HFHC diet to a standard diet that is provided by the UKZN animal unit. Figure 1 shows an illustration of animal groupings according to the treatments.

Animal groupings according to treatment
Treatment of pre-diabetic animals
The treatment period lasted for 12 weeks. The animals were treated every third day where the MET and MET+DI groups received MET (500 mg/kg p.o., with oral gavage) while the OA and OA+DI groups were given OA (80 mg/kg p.o., with oral gavage ) dissolved in DMSO and water (10% DMSO). Parameters, including total cholesterol and triglyceride concentrations, were measured every fourth week in all groups for the duration of the treatment period.
Blood collection and tissue harvesting
At the end of the 12-week treatment period, the animals were euthanised. For blood collection, all animals were anaesthetised with Isofor (100 mg/kg) [Safeline Pharmaceuticals (Pty) Ltd, Roodeport, South Africa] via a gas anaesthetic chamber (Biomedical Resource Unit, UKZN, Durban, South Africa) for 3 min. Blood was collected by cardiac puncture and injected into the pre-cooled heparinised containers. The blood was then centrifuged (Eppendorf centrifuge 5403, Eppendorf, Germany) at 4°C, 2000 × g for 15 min. The adipose tissue was also harvested for the analysis of cytokines. Plasma was collected and stored at −80°C in a Bio Ultra freezer (Snijers Scientific, Tilburg, the Netherlands) until ready for biochemical analysis.
Differential white blood cell count analysis
For analysis of differential white blood cell count, such as monocytes, neutrophils, eosinophils, basophils, total lymphocytes and platelets as well as mean platelet volume, blood was obtained using a tail prick method, where about 100 µl blood was transferred into tubes containing anti-coagulant, thereafter white blood cell count and mean platelet volume (MPV) were determined using a Coulter AcT diff Tainer haemocytometer (Beckman Coulter, Inc., Indianapolis, IN, USA).
Biochemical analysis
Oxidative stress was determined using a well-established malondialdehyde (MDA) assay protocol, whereas antioxidant enzymes, including superoxide dismutase (SOD) and glutathione peroxidase (GPx), were measured using Elabscience ELISA kits (Biocom Africa). Plasma IL-1β, CRP, TNF-α, fibrinogen and CD40L were also analysed using their respective rat ELISA kits (Elabscience, purchased from Biocom Africa) according to the manufacturer’s instructions.
Statistical analysis
The data was expressed as mean ± standard error of the mean (SEM). Statistical comparisons were performed with GraphPadInStat Software (version 5.00, Graph Pad Software, Inc., San Diego, CA, USA) using one-way analysis of variance (ANOVA) followed by Bonferroni post hoc comparison test. A value of p < 0.05 was considered statistically significant.
Results
Platelets and mean platelet volume
At the end of the treatment period (week 12), whole blood was collected from all the groups for full blood count using a haemocytometer. The results showed that PC had a significantly higher platelet count and MPV when compared with NPC (p < 0.05). However, OA treatment with or without dietary intervention showed a significant decrease in both platelets and MPV (Figure 2A,B; p < 0.005). Treatment with both MET and MET+DI followed the same trend as OA treatments in platelets count as well as MPV.

Bar graph showing the effects of OA and MET, with and without DI, on platelet count (A) and MPV (B). Values are presented as mean ± SEM (n = 6). = p < 0.05 denotes comparison with NPC; α = p < 0.05 denotes comparison with PC.
Plasma lipoproteins
The terminal plasma lipids were analysed in all experimental groups. The results showed that PC had a significantly higher low-density lipoprotein cholesterol (LDL-C) and high-density lipoprotein cholesterol (HDL-C) when compared with NPC (p < 0.05). The OA with dietary intervention treatment resulted in a significant decrease in HDL-C and LDL-C when compared with PC (Figure 3A, B; p < 0.05). MET had no significant difference in HDL-C in comparison with PC. However, MET+DI showed a significance decrease in HDL-C in comparison with PC (Figure 3A; p < 0.05).

Bar graph showing the effects of OA, and Met with and without DI on plasma HDL-C (A) and LDL-C (B) concentration. Values are presented as mean ± SEM (n = 6). * = p < 0.05 denotes comparison with NPC; α = p < 0.05 denotes comparison with PC.
Monocytes, neutrophil, lymphocytes and eosinophil count
Whole blood at the end of the treatment period (week 12) was collected and used to measure immune cells, that is, neutrophils, monocytes and lymphocytes counted using a haemocytometer. The monocyte count from PC showed a significant increase when compared with NPC. However, treatment with both MET and OA in the presence and absence of dietary intervention resulted in a significant decrease in comparison with PC (Figure 4A; p < 0.05). The neutrophil count from PC exhibited a significant decrease when compared with NPC. However, treatment with OA in the presence of dietary intervention restored plasma neutrophils, which also resembled MET and MET+DI (Figure 4B). The lymphocyte count from PC exhibited a significant increase in plasma lymphocytes in comparison with NPC; p < 0.05. However, OA and MET treatments both showed a significant decrease in lymphocyte count in comparison with PC (Figure 4C; p < 0.05). There were no significance differences between NPC and PC and OA, both in the presence and absence of dietary intervention, and no significant difference in comparison with PC and NPC (Figure 4D; p < 0.005).

>Bar graph showing the effects of OA, and Met with and without DI on plasma HDL-C (A) and LDL-C (B) concentration. Values are presented as mean ± SEM (n = 6). = p < 0.05 denotes comparison with NPC; α = p < 0.05 denotes comparison with PC.
Proinflammatory cytokines
At the end of the treatment period (week 12), plasma IL-1β and TNF-α concentrations were measured in plasma samples from all experimental groups. The results show that PC had high circulating IL-1β concentrations when compared with NPC; (p < 0.05). However, OA with and without dietary intervention treatment resulted in reduced plasma IL-1β in comparison with PC (p < 0.05). Plasma TNF-α concentrations in PC were significantly higher than in NPC (p < 0.05). In all groups where OA was administered (OA and OA+DI), there was a significant (p < 0.05) decrease in plasma and TNF-α concentration when compared with PC. The OA-treated group had no significant difference when compared with PC (p < 0.05). However, OA+DI showed a significant decrease in plasma TNF-α concentration when compared with PC (p < 0.05; Table 1). MET and MET+DI showed no statistical significance in IL-1β concentrations when compared with PC. However, both showed a significance decrease in TNF-α concentration in comparison with PC (p < 0.05; Table 1).
Effects of OA and MET with and without dietary intervention on adipose tissue IL-1β and TNF-α concentration.

DI, dietary intervention; IL, interleukin; Met+DI, metformin-treated with the DI; NC, non-pre-diabetic control; PC, pre-diabetes control; OA+DI, oleanolic acid-treated with the DI; TNF, tumour necrosis factor.
Values are presented as mean ± SEM, (n = 6) *= p < 0.05 denotes comparison with NPC; α = p < 0.05 denotes comparison with PC.
Plasma CD40L
Plasma CD40-L concentration was measured in all the experimental groups at the end of the treatment period (week 12). The results showed that plasma CD40-L concentration of PC was significantly higher than of NPC (p < 0.05). In all groups where OA was administered (OA and OA+DI), there was a significant decrease in CD40-L concentration when compared with PC (p < 0.05; Figure 5). MET and MET+DI also followed the same trend as OA and OA+DI (p < 0.05; Figure 5).
Bar graph showing the effects of OA and Met with and without DI on plasma CD40-L. Values are presented as mean ± SEM (n = 6). =p < 0.05 denotes comparison with NPC; α = p < 0.05 denotes comparison with PC.
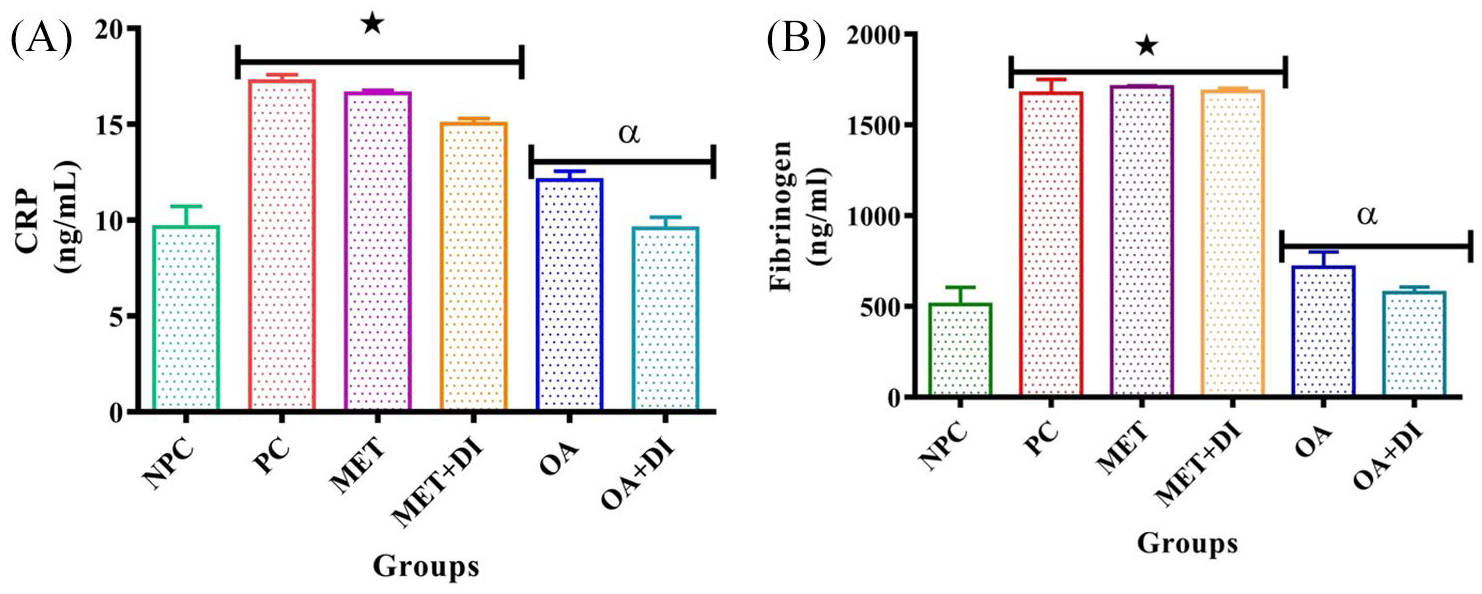
CRP and fibrinogen
Plasma CRP and fibrinogen concentrations were determined in all experimental groups at the end of the treatment period (week 12). The results showed that both CRP and fibrinogen concentration of PC was significantly higher than that of NPC (p < 0.05). OA treatment resulted in a significant decrease in both CRP and fibrinogen when compared with PC (p < 0.05; Figure 5). In CRP and fibrinogen concentrations both MET and MET+DI had no statistical significance in comparison with PC (p < 0.05; Figure 5A,B).
Discussion
Previous studies in our laboratory have alluded to the fact that plant-derived OA, in the presence and absence of dietary intervention, ameliorates markers associated with cardiovascular complications such as dyslipidaemia, production of pro-inflammatory cytokines, mean-arterial pressure and cardiac hypertrophy. 17 In addition, another study from our laboratory showed that OA also ameliorates markers of non-alcoholic fatty liver disease, including intrahepatic fat accumulation and hepatic inflammation. 18 The pathogenesis of the aforementioned diseases is linked closely to chronic sub-clinical inflammation and activated innate immunity. This is evidenced by the presence of chronic inflammation in diseases such as atherosclerosis and hepatosteatosis. 20 Hence, these previous findings led to the investigation of the effects of OA on pre-diabetes-induced innate immunity activation and the resulting sub-clinical chronic inflammation.
Activation of innate immunity and the inflammatory pathway during pre-diabetes is one of the contributing factors that facilitate the progression of pre-diabetes to overt T2DM. 21 Previous studies have reported that high energy diets and a sedentary lifestyle correlate with the prevalence of metabolic abnormalities with immune activation. 22 Diets that are rich in fats are implicated in the pathogenesis of complications such as subclinical inflammation, cardiovascular diseases and T2DM. 23 The pathogenesis of diet-induced subclinical inflammation may be attributed to the elevation of LDL and reduced HDL in circulation. In addition, increased intestinal permeability to microbiota, which leads to increased endotoxin such as LPS. 24 LPS in the circulation interacts directly with the PRR that are found on the surface of immune cells, resulting in their activation and release of pro-inflammatory cytokines. 7 Augmented pro-inflammatory cytokines exacerbate and sustain the systemic inflammation that results from hyperglycaemic-induced oxidative stress during pre-diabetes and T2DM. 25 Targeting inflammation in diabetes has the potential to provide an effective strategy in the management of diabetes. 26 Conventional treatments for T2DM, including MET, regulate glucose metabolism. However, patients continue to develop inflammatory-related complications such as steatohepatitis and atherosclerosis. 27 Therefore, there is a need for treatment that offers both glycaemic control and anti-inflammatory effect to prevent the progression to T2DM. 28 Previous studies in our laboratory have shown that the diet-induced pre-diabetes rat model is characterised with metabolic and cardiovascular complications including obesity, dyslipidaemia, oxidative stress, insulin resistance, impaired glucose tolerance and hypertension. 17 In these studies, OA treatment was shown to ameliorate metabolic derangements. 17 Hence, this study further investigated the effects of OA on selected markers of innate immunity activation and chronic low-grade inflammation, which also play a role in the progression of pre-diabetes to overt diabetes.
High-calorie diets such as high fat or high carbohydrate diets have both primary and secondary implications in the development of inflammatory diseases. 29 The primary implications may include the elevation of lipid metabolites such as LDL in the circulation, which may bind to circulating immune cells and lead to the production of pro-inflammatory cytokines. 30 Moreover, the level of circulating endotoxins such as LPS may arise secondarily to the over-indulgence of a high caloric diet, and HDL prevents the binding of LPS from binding to TLRs on the surface of immune cells, and, subsequently, the formation of inflammasome. 30 The current study found that diet-induced pre-diabetic animals had elevated plasma LDL-C and plasma HDL-C. However, treatment with OA combined with dietary intervention resulted in the reduction of HDL-C and LDL-C. These findings correlate with results from previous studies that reported that OA possesses anti-lipidaemia properties. 31 This is attributed to the downregulation of genes such as peroxisome proliferator activator receptor gamma (PPARγ) and adiponectin receptor-1 (ADPOR1), which are involved in the utilisation of fatty acids for energy production. 31
Diet-induced pre-diabetes has been previously reported to contribute to the activation of innate immunity cells including monocytes, neutrophils, eosinophils and lymphocytes. 32 This may be attributed to mechanisms such as insulin resistance induced glucotoxicity, lipotoxicity and oxidative stress. 11 Moreover, an increase in endotoxins such as LPS due to intestine permeability is also associated with increased innate immune cell count. 33 The present study found that untreated pre-diabetic animals had increased circulating monocytes, eosinophils and lymphocytes and decreased neutrophils. The decrease in circulating neutrophils correlated with findings from previous studies that reported a reduction in neutrophil count during pre-diabetes and early diabetes. 34 This may be accredited to intestine permeability and leakage of LPS from the gut as a result of over-indulgence in a HFHC diet and divergence of glucose to the polyol pathway. 33 However, the administration of OA with dietary intervention led to a reduction in these cells. This may be attributed to improved lipid and glucose metabolism following improved insulin sensitivity or decrease in calorie intake. 35 Furthermore, OA has been reported to reduce eosinophilic inflammation and suppress the signal transducer and activator of transcription 3 (STAT3) that sustains the activation of immune cells. 36 However, the switch from a high-fat diet to a standard diet may have lowered circulating endotoxins by reducing intestine permeability. 33 During pre-diabetes, monocytes infiltrate inflamed tissues such as adipose tissue where they differentiate to macrophages. 37 The accumulated neutrophils and macrophages in the inflamed tissues such as adipose tissues advance the inflammation by secreting proinflammatory cytokines. 38 Pre-diabetes is also associated with increased circulating proinflammatory mediators, including interleukin 1beta (IL-1β), interleukin-1 receptor agonist (IL-1RA), interleukin-6 (IL-6), interleukin-8 (IL-8) and monocyte chemotactic protein-1 (MCP-1). 39
Insulin resistance is further associated with adipose tissue hypertrophy, which is also implicated in the production of proinflammatory cytokines during pre-diabetes. 40 In this study, we observed that pre-diabetic animals had elevated pro-inflammatory cytokines including TNF-α, IL-6 and IL-1β. This could also be due to hypertrophy of adipose tissue due to the consumption of a HFHC diet, resulting in the secretion of inflammatory mediators from the adipocytes inflammation. 41 This could have led to adipose macrophage infiltration and activation of nuclear factor-kappa B (NF kB), as well as secretion of proinflammatory cytokines, hence low-grade inflammation. 41 The activation of NF-kB is linked directly with the activation of PRR on the surface of immune cells that release pro-inflammatory cytokine. 42 Elevation of pro-inflammatory cytokines is also associated with insulin resistance, and it precedes the onset of overt T2DM in pre-diabetic individuals. 43 Treatment with OA with and without dietary intervention resulted in a reduction of these pro-inflammatory cytokines. This may be attributed to activation of transforming growth factor-beta (TGF-β), which downregulates PRRs such as TLR4 and CD14, thus attenuating innate immune response that leads to the release of proinflammatory cytokines. 44 However, in vivo studies have also shown that dietary interventions such as low-fat diet ameliorate chronic inflammation. 45 Chronic inflammation is also driven by inflammatory mediators such as CRP. 46 CRP is one of the principal downstream inflammatory mediators that is synthesised by hepatocytes in response to IL-6. 47 Untreated pre-diabetic animals were found to have elevated levels of CRP. This may be attributed to the observed elevated plasma IL-6 concentrations. However, an increase in plasma lipids may also stimulate the secretion of CRP in cells, including adipocytes and hepatocytes. 48 The high levels of CRP are associated with elevated secretion of proinflammatory cytokines such as IL-6 which sustain inflammation. 48 Similarly, administration of OA with and without dietary intervention resulted in reduced levels of CRP. This suggests that IL-6 may be a causative factor in the elevated CRP concentrations. 49 However, these findings may also be attributed to the anti-adiposity activity of OA through the inactivation of hunger centres and reduced release of ghrelin hormone. 35 Increased circulating fibrinogen is one of the markers of a proinflammatory state. 50 Hyperglycaemic-induced platelet reactivity is associated with elevated hepatic fibrinogen production. 51 Moreover, insulin resistance-induced oxidative stress is associated with increased circulating thrombin, which stimulates hepatic fibrinogen production. 52 The role of fibrinogen in sustaining inflammation includes binding and activation of several immune cells such as neutrophil and monocytes that release pro-inflammatory chemokines as well as cytokines. 53 In the current study, administration of OA with and without dietary intervention reduced circulating fibrinogen. This may be attributed to the previously reported anti-hyperglycaemic and anti-oxidant effects of OA. 35 In addition, these findings may also be due to anti-hyperlipidaemic activities of OA and suppression of innate immune cell response. 54
Increased platelet counts and MPV are associated with glucose homeostasis dysregulation and immune activation. 55 The current study found that untreated pre-diabetic animals had increased platelet count and MPV, which is associated with the pathogenesis of cardiovascular diseases such as atherosclerosis and thrombosis. Augmented platelet count and platelet activation may be attributed to intraplatelet hyperglycaemia due to systemic insulin resistance. 56 Systemic insulin resistance affects insulin-dependent tissues such as skeletal muscle and adipose tissue. 57 This resistance leads to increased circulating glucose and glucose influx to cells that do not depend on insulin for glucose uptake. 58 Cells such as those found in the liver and kidney as well as platelets, use facilitated diffusion through glucose transporter 1 (GLUT 1) and glucose transporter 2 (GLUT2). 59 Interestingly, treatment with OA with and without dietary intervention reduced both platelet count and activation. This may be attributed to the anti-hyperglycaemic effects of OA through improved sensitivity of skeletal muscles to insulin and increased translocation of GLUT4, thus promoting glucose uptake. 60 However, the synergistic effects of dietary intervention may also contribute to the reduction of circulating glucose through the reduction of carbohydrate intake. 61 Moreover, these findings correlated with previous studies such as that of Osunsanmi et al., who reported that triterpenes have anti-platelet aggregation properties through modifying hydroxyl moiety at the carbon-3 position with the acetyl group, and prevent platelet activation. 62 The decrease in platelet activation may prevent the pathogenesis of cardiovascular complications in pre-diabetes, which is reported mostly in people with diabetes and metabolic syndrome. In addition, treatment with MET both in the presence and in the absence of dietary intervention yielded similar results, as expected. MET has been reported previously to have anti-thrombin properties by preventing platelet aggregation. 63 The present study also observed that pre-diabetes animals had elevated plasma CD40L concentrations, which may be attributed to platelet and monocyte activation due to hyperglycaemia. 64 CD40L is an inflammatory mediator that belongs to the TNF superfamily that is expressed by immune cells including monocytes, T-cells and activated platelets. 65 Interaction of the CD40L with mucosa T cells induces the production of chemokines that advance inflammation by promoting the production of inflammatory cytokines. 66 However, administration of OA with and without dietary intervention led to a reduction in plasma CD40L concentrations. Although the mechanism by which OA may lower plasma CD40L is not yet established, the suppression of platelet activation may result in the reduction of CD40L in circulation. 67
This study used MET as a positive control drug. The use of MET is recommended to be combined with dietary intervention. As expected, MET treatment in the presence of dietary intervention also ameliorated the proinflammatory cytokines and immune cell activation, which correlated with findings from previous studies reporting that MET reduces the production of autoantibodies and infiltration of inflammatory cells such as monocytes, neutrophils and lymphocytes into tissues. 63 Moreover, MET can inhibit CD40L, thereby suppressing platelet-activation-mediated inflammation. 63 However, in parameters such as plasma LDL-C, fibrinogen and CRP, OA was seen to be more potent than MET. This requires more studies comparing MET and OA in the pre-diabetes state.
Conclusion
The findings of this study suggest that administration of OA with and without dietary intervention has anti-inflammatory properties in pre-diabetes through attenuating innate immunity cell response and suppressing pro-inflammatory cytokine production. These findings may serve as a basis for further investigations of OA as a drug that can prevent the progression of progression of pre-diabetes to T2DM. However, more studies need to be done in order to elucidate the mechanism by which this treatment ameliorates inflammation during pre-diabetes, and the effects of OA in the permeability of the gastrointestinal tract.
Footnotes
Acknowledgements
The authors are grateful to the Biomedical Resource Unit, the University of KwaZulu-Natal for the supply of animals, and to Dennis Makhubela for his technical expertise.
Author contribution(s)
Conflict of interest statement
The authors declare that there is no conflict of interest.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: National Research Foundation (South Africa) for financial support (grant number 106041)
Novelty statement
The ingestion of high caloric diets results in subclinical inflammation that worsens the progression of pre-diabetes to overt type 2 diabetes (T2DM). Oleanolic acid (OA) has been found to have anti-hyperglycaemic properties. This study found that OA prevents the progression of pre-diabetes toward T2DM by reducing systemic inflammation in a pre-diabetic rat model. These results encourage further investigations into whether OA can be used to provide an alternative treatment for preventing the progression of pre-diabetes to T2DM.