
Abstract
Mesenchymal stem cells maintained in appropriate culture conditions are capable of producing robust cartilage tissue. However, gradients in nutrient availability that arise during three-dimensional culture can result in the development of spatially inhomogeneous cartilage tissues with core regions devoid of matrix. Previous attempts at developing dynamic culture systems to overcome these limitations have reported suppression of mesenchymal stem cell chondrogenesis compared to static conditions. We hypothesize that by modulating oxygen availability during bioreactor culture, it is possible to engineer cartilage tissues of scale. The objective of this study was to determine whether dynamic bioreactor culture, at defined oxygen conditions, could facilitate the development of large, spatially homogeneous cartilage tissues using mesenchymal stem cell laden hydrogels. A dynamic culture regime was directly compared to static conditions for its capacity to support chondrogenesis of mesenchymal stem cells in both small and large alginate hydrogels. The influence of external oxygen tension on the response to the dynamic culture conditions was explored by performing the experiment at 20% O2 and 3% O2. At 20% O2, dynamic culture significantly suppressed chondrogenesis in engineered tissues of all sizes. In contrast, at 3% O2 dynamic culture significantly enhanced the distribution and amount of cartilage matrix components (sulphated glycosaminoglycan and collagen II) in larger constructs compared to static conditions. Taken together, these results demonstrate that dynamic culture regimes that provide adequate nutrient availability and a low oxygen environment can be employed to engineer large homogeneous cartilage tissues. Such culture systems could facilitate the scaling up of cartilage tissue engineering strategies towards clinically relevant dimensions.
Introduction
Articular cartilage has a limited capacity for self-repair and as a result, cell-based strategies are being explored to repair or replace damaged cartilage tissues. Novel combinations of cells, scaffolds, growth factors and bioreactors are being developed to engineer functional cartilage tissue replacements in vitro.1–7 Mesenchymal stem cell (MSC) is a promising cell source to engineer these replacement tissues as they are readily expandable in vitro and can be differentiated along a chondrogenic pathway in three-dimensional (3D) culture.8,9 In theory, they are a more attractive cell source than chondrocytes due to difficulties in obtaining large numbers of healthy, functional chondrocytes in a clinical setting. However, when MSCs are cultured in identical conditions to chondrocytes, they have been shown to produce a cartilage matrix of inferior quality.5,10
Nutrient availability to cells throughout engineered 3D tissues in vitro occurs by diffusion and therefore depends on their distance from a free surface, diffusional constraints and cellular utilisation at the periphery. 11 These nutritional gradients lead to local differences in extracellular matrix (ECM) deposition within engineered tissues. It has been demonstrated that MSCs produce robust cartilage matrix in regions of optimal nutrient supply close to a free surface; however, inferior matrix accumulation occurs within core regions further from a free surface. 12 Importantly, within these peripheral regions, MSCs are capable of producing tissue that matches or even exceeds the quality of that produced by chondrocytes. 12 Diffusional limitations become exacerbated when attempting to engineer larger cartilage tissues using MSCs. For example, core regions devoid of cells and/or matrix were reported when MSCs were cultured in collagen meshes (8 × 2 mm) under chondrogenic conditions 13 or within high cell density hydrogels. 14 Strategies are therefore required to overcome these diffusional limitations to engineer functional cartilage tissue using MSCs.
Dynamic culture regimes involving fluid agitation have been shown to overcome diffusional limitations producing tissues of superior quality relative to free swelling conditions. Application of flow perfusion and simple spinner flask culture systems has been shown to enhance matrix accumulation by chondrocytes compared to static conditions.15–19 Forced convection of media through microchannelled hydrogels within a rotational bioreactor system has been shown to enhance matrix accumulation within large cartilage tissues engineered using chondrocytes. 20 However, interestingly, the same rotational culture system was shown to suppress chondrogenesis of MSCs. 21 In similar bioreactor systems, flow perfusion culture was shown to interfere with the chondrogenic differentiation of MSCs resulting in lower levels of sulphated glycosaminoglycan (sGAG) accumulation compared to statically cultured samples.22,23 However, there have been studies reporting improved chondrogenesis of MSCs under dynamic culture conditions.24,25 The effectiveness of dynamic culture regimes on enhancing chondrogenesis of MSCs is therefore unclear.
Dynamic bioreactor systems that enhance nutrient delivery to cells in 3D culture will also enhance local oxygen availability within such engineered tissues. Oxygen tension is a potent regulator of MSC fate with lower oxygen tensions known to support enhanced chondrogenic differentiation of MSCs.1,26–30 Dynamic culture has been shown to increase oxygen transport within central regions of MSC seeded fibrin hydrogels, which correlated with suppressed chondrogenesis compared to static conditions. 31 In static culture conditions, oxygen gradients develop, producing core hypoxic regions in which cartilage matrix synthesis by MSCs is increased. 32 We hypothesize that by dynamically culturing samples to overcome nutrient diffusion limitations, while simultaneously maintaining low oxygen conditions, that it will be possible to engineer homogenous cartilage tissues of scale using MSCs. Therefore, the first objective was to explore whether a dynamic culture system could enhance chondrogenesis of MSCs in both small (5 mm Ø × 3 mm) and large (10 mm Ø × 6 mm) channelled alginate hydrogels compared to static conditions. The second objective was to explore the role of oxygen tension in regulating the response of MSCs to dynamic culture regimes by engineering such cartilaginous tissues at both 20% O2 and 3% O2 conditions. The dynamic culture system was developed by engineering a bioreactor capable of translating 3D cell laden channelled alginate hydrogel constructs throughout a bath of cell culture media. Introducing channels into cell laden hydrogels is a common strategy to facilitate enhanced nutrient transfer while engineering large cartilage tissues.19,21,33–36 Alginate was used as the hydrogel material as it has been shown to support robust MSC chondrogenesis and supports a rounded chondrocyte-like cell morphology.37–39
Methods
Experimental design and bioreactor design
A custom-made bioreactor was developed to dynamically agitate and perfuse media through the channels of the MSC laden hydrogels throughout the culture period. A linear actuator was used to control the displacement of a porous stainless steel platform which housed cell strainers. MSC seeded hydrogels could then be placed in a cell strainer (Figure 1(c)) which was fixed in place within the porous platform (Figure 1(b)) and continuously translated through a bath of cell culture media (Figure 1(a)). This facilitated constant agitation of the cell culture media in the bath. The translation of the constructs in the cell strainer facilitated continuous perfusion of media relative to the peripheral and channelled free hydrogel surfaces. The whole bioreactor unit was placed in a cell culture incubator where the oxygen conditions could be controlled. The constructs were then continuously translated at a speed of 0.05 mm/s and an amplitude of 6 mm for the entire culture period. Small (5 mm Ø × 3 mm) and large (10 mm Ø × 6 mm) MSC laden alginate hydrogels containing 1 mm channels were placed in cell strainer. As a static, control gels were placed in an identical housing system except the actuator was not translated. Constructs were then cultured in static and dynamic conditions for 4 weeks at an external oxygen tension of 20% O2 and 3% O2. Media (400 mL per cell culture bath) was changed twice weekly.
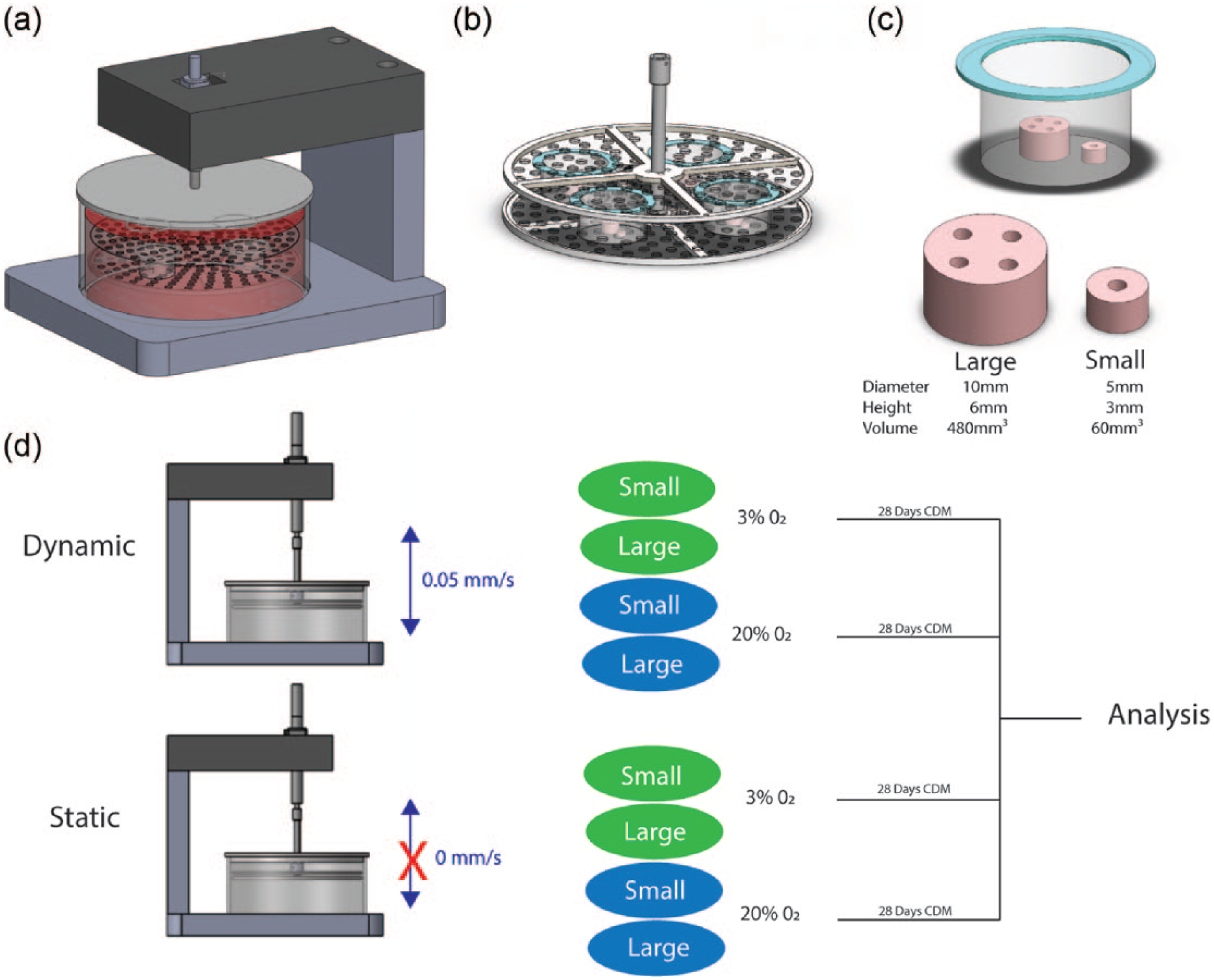
(a) Complete bioreactor design where a stainless steel platform was connected to a linear actuator. (b) Stainless steel platform containing fixed cell strainers housing MSC seeded alginate hydrogels. (c) Small and large alginate hydrogels placed within a cell strainer. (d) Experimental conditions; static versus dynamic culture for small and large construct in both 3% O2 and 20% O2.
Isolation and expansion of MSCs
Bone marrow–derived MSCs were isolated from the femoral shaft of 4-month-old pigs and expanded according to a modified method for human MSCs 26 in high-glucose Dulbecco’s modified Eagle’s medium (hgDMEM) GlutaMAX supplemented with 10% v/v fetal bovine serum (FBS), 100 U mL−1 penicillin/100 μg mL−1 streptomycin (all Gibco Biosciences, Dublin, Ireland) and 2.5 μg mL−1 amphotericin B (Sigma-Aldrich, Dublin, Ireland) at 20% pO2. Tri-potentiality was confirmed prior to use. Following colony formation, MSCs were trypsinised, counted, seeded at density of 5000 cells cm2 in 500 cm 2 triple flasks (Thermo Fisher Scientific), supplemented with hgDMEM, 10% v/v FBS, 100 U mL−1 penicillin/100 μg mL−1 streptomycin, 2.5 μg mL−1 amphotericin B and 5 ng mL−1 human fibroblastic growth factor-2 (FGF-2; ProSpec-Tany TechnoGene Ltd, Israel) and expanded to passage 2.
MSC laden alginate hydrogels
Alginate constructs were fabricated by placing MSC laden alginate (2% w/v ultrapure, low viscosity (20–200 MPa s) sodium alginate; Pronova, FMC Biopolymer, Norway) into custom designed solid 4% agarose/50 mM CaCl2 slab and allowing to gelate for 30 min after covering the alginate solution in the moulds with a solid 4% agarose/50 mM CaCl2 slab. MSCs were trypsinised, counted and re-suspended into a single-cell solution in the 2% alginate (2 × 107 MSCs/mL) prior to gelation. For fabrication of the small constructs, a 59 µL MSC/alginate suspension (2%, 2 × 107 MSCs/mL) was pipetted into a 5 mm diameter, 3 mm height agarose mould. For fabrication of the large constructs, a 471 µL MSC/alginate suspension (2%, 2 × 107 MSCs/mL) was pipetted into a 10 mm diameter, 6 mm height agarose mould. Channels were introduced into the alginate using a 1 mm diameter biopsy punch. The number of channels designated was based on the surface area of the construct. The number of channels was designated by keeping the ratio of construct area to total channel area constant between the large and small constructs. The parameters used for fabricating the groups are described in Table 1.
Description of construct parameters used for each group.

Chondrogenic culture conditions
The chondrogenic culture conditions applied in this study are defined as culture in a chondrogenic medium (CM) consisting of hgDMEM GlutaMAX supplemented with 100 U mL−1 penicillin/streptomycin (both Gibco), 100 μg mL−1 sodium pyruvate, 40 μg mL−1 L-proline, 50 μg mL−1
Biochemical analysis
The biochemical content of all hydrogels was analysed after 4 weeks of in vitro culture. Prior to biochemical analysis, constructs were washed in phosphate buffered saline (PBS), weighed and frozen for subsequent assessment. Each construct was digested with papain (125 μg mL−1) in 0.1 M sodium acetate, 5 mML-cysteine–HCl, 0.05 methylenediaminetetraacetic acid (EDTA), pH 6.0 (all from Sigma-Aldrich) at 60°C and 10 r/min for 18 h. DNA content was quantified using the Hoechst Bisbenzimide 33258 dye assay, with a calf thymus DNA standard. The amount of sGAG was quantified using the dimethyl methylene blue dye-binding assay (Blyscan, Biocolor Ltd, Northern Ireland), with a chondroitin sulphate standard. Total collagen content was determined by measuring the hydroxyproline content, using a hydroxyproline-to-collagen ratio of 1:7.69. 40 The other half was digested in 1 M hydrochloric acid at 60°C and 10 r/min for 18 h. The calcium content was determined using a Sentinel Calcium kit (Alpha Laboratories Ltd, UK). Three or four constructs per group were analysed by each biochemical assay.
Histological and immunohistochemical analysis
Constructs were fixed in 4% paraformaldehyde, dehydrated in a graded ethanol series, embedded in paraffin wax, sectioned at 8 µm and affixed to microscope slides. The sections were stained with aldehyde fuschin/alcian blue to assess sGAG content and Picrosirius red to assess collagen content. Collagen types II and X were evaluated using a standard immunohistochemical technique; briefly, sections were treated with peroxidase, followed by treatment with chondroitinase ABC (Sigma-Aldrich, Dublin, Ireland) in a humidified environment at 37°C to enhance permeability of the ECM. Sections were incubated with goat serum to block non-specific sites and collagen type II (ab3092, 1:100; 1 mg/mL) or collagen type X (ab49945, 1:200; 1.4 mg/mL) primary antibodies (mouse monoclonal, Abcam, Cambridge, UK) were applied for 1 h at room temperature. Next, the secondary antibody (anti-mouse (Immunoglobulin G) IgG biotin conjugate, 1:200; 2.1 mg/mL) (Sigma-Aldrich, Dublin, Ireland) was added for 1 h followed by incubation with ABC reagent (Vectastain PK-400; Vector Labs, Peterborough, UK) for 45 min. Finally, sections were developed with (3, 3’-diaminobenzidine) DAB peroxidase (Vector Labs) for 5 min. Positive and negative controls were included in the immunohistochemistry staining protocol for each batch.
Live/dead confocal microscopy
Cell viability was assessed after 24 h using a LIVE/DEAD™ viability/cytotoxicity assay kit (Invitrogen, Bioscience, Ireland). Briefly, constructs were cut in half, washed in PBS followed by incubation in PBS containing 2 µM calcein acetoxymethyl (AM) (green fluorescence of membrane for live cells) and 4 µM ethidium homodimer-1 (red fluorescence of DNA for dead cells; both from Cambridge Bioscience, UK). Sections were again washed in PBS, imaged at magnification 10× with an Olympus FV-1000 Point-Scanning Confocal Microscope (Southend-on-Sea, UK) at 515 and 615 nm channels and analysed using FV10-ASW 2.0 Viewer software. Live dead quantification was carried out using ImageJ.
Statistical analysis
Statistical analyses were performed using the software package MINITAB 15.1 (Minitab Ltd, Coventry, UK). Groups were analysed for significant differences using a general linear model for analysis of variance. Tukey’s test for multiple comparisons was used to compare conditions. Significance was accepted at a level of p ≤ 0.05. Numerical and graphical results are presented as mean ± standard deviation, with graphical results produced using GraphPad Prism (Version 6.02).
Results
Influence of dynamic culture and external oxygen tension on cell viability within constructs of different scales
Dynamic culture did not significantly enhance cell viability compared to static conditions under either oxygen condition for both small and large constructs; however, there was a trend towards higher cell viability with dynamic culture in larger constructs at both oxygen conditions. DNA content (ng/mg) was significantly higher in small dynamically cultured gels compared to larger gels (static and dynamic) at 3% O2 (Figure 2(a)). No significant differences were found between any of the groups at 20% O2 (Figure 2(a)). Live dead analysis revealed a significantly lower cell viability in larger gels compared to small gels cultured in static conditions at both oxygen tensions (Figure 2(b)). However, this decrease in viability was not significant when the large gels were dynamically cultured (Figure 2(b)).

Analysis of cell viability within channelled MSC laden alginate hydrogels under all conditions after 28 days of culture. Images are taken from central portion of gels. (a) DNA content normalised (ng/mg) (n = 4, p < 0.05, ANOVA, mean ± SD). (b) Quantification of cell viability from live dead images (n = 3, p < 0.05, ANOVA, mean ± SD, ‘a’ denotes significance between connected groups). (c) Representative live dead images.
Influence of dynamic culture and external oxygen tension on the expression of chondrogenic and hypertrophic markers within MSC laden hydrogels of different scales
Compared to static conditions, dynamic culture was found to either significantly increase or decrease sGAG accumulation within the engineered cartilage tissues depending on the external oxygen tension. At 3% O2, dynamic culture significantly enhanced sGAG accumulation in large samples, with no benefit of dynamic culture found in small samples (Figure 3(a)). At 20% O2, dynamic culture significantly decreased sGAG synthesis at both construct scales (Figure 3(a)). Histological staining revealed an inhomogeneous spatial distribution of sGAG across the construct cross sections, especially in static culture conditions (Figure 3(b)). Core regions of large statically cultured samples were devoid of sGAG at both oxygen tensions. In contrast, dynamic culture at 3% O2 supported a homogenous distribution of sGAG throughout the construct cross section (Figure 3(b)). Confirming the biochemical analysis, low levels of sGAG staining were found in constructs dynamically cultured at 20% O2 (Figure 3(b)).

Histological and biochemical analysis of sGAG and collagen content within channelled MSC laden alginate hydrogels under all conditions after 28 days of culture. (a) Biochemical sGAG and collagen content normalised (%ww) (n = 4, p < 0.05, ANOVA, mean ± SD, ‘a’ denotes significance between connected groups). (b) Histological staining for sGAG (aldehyde fuschin/alcian blue) and collagen (Picrosirius red), 2x scale 1 mm, representative images for n = 2.
Unlike sGAG synthesis, dynamic culture was not found to enhance collagen accumulation at 3% O2 compared to static conditions (Figure 3(a)). Significantly higher levels of collagen accumulation were found in gels cultured in static conditions compared to dynamic conditions at 20% O2 (Figure 3(a)). Histological staining again revealed significant spatial inhomogeneity in collagen accumulation across the construct cross sections (Figure 3(d)). The core regions of large, statically cultured constructs were again devoid of collagen matrix. In contrast, a more homogenous distribution of collagen matrix was observed in dynamically cultured tissues (Figure 3(d)). Confirming the biochemical data, low levels of collagen staining were found in constructs cultured dynamically at 20% O2 (Figure 3(d)).
Intense collagen type II staining was found in small-scale constructs cultured at 3% O2 in both static and dynamic conditions (Figure 4(a)). Spatial variations in collagen type II deposition were found within cross sections of scaled-up constructs. Core regions of large statically cultured samples were devoid of collagen type II, while a more homogenous distribution was found under dynamic conditions (Figure 4(a)). When the oxygen tension was switched to 20% O2, collagen type II was only found in statically cultured samples (Figure 4(a)). Negligible collagen type X staining was found under all conditions (Figure 4(b)).

Immunohistochemical staining for (a) collagen type II and (b) collagen type X within channelled MSC laden alginate hydrogels under all conditions after 28 days of culture, 2X scale 1 mm, representative images for n = 2.
Higher levels of calcium were deposited in samples cultured at 20% O2 compared to 3% O2 (Figure 5(a) and (b)). At 3% O2, negligible calcium was observed histologically, with low levels of calcium accumulation measured using biochemical analysis (Figure 5(a) and (b)). No significant differences in the levels of calcium content were produced between statically and dynamically cultured samples at either scale at 20% O2 (Figure 5(a)).

Histological and biochemical analysis of calcium content within channelled MSC laden alginate hydrogels under all conditions after 28 days of culture. (a) Biochemical calcium content normalised (%ww) (n = 4, p < 0.05, ANOVA, mean ± SD, ‘a’ denotes significance between connected groups). (b) Histological staining for calcium (Alizarin red), 2X scale 1 mm, representative images for n = 2.
Discussion
Engineering cartilage tissues of a sufficient scale to treat defects observed clinically is a major challenge in cartilage tissue engineering. This challenge amplifies for putative endochondral bone tissue engineering strategies where engineered cartilage is used to treat large bone defects.41–43 The objective of this study was to determine whether enhancing nutrient diffusion using dynamic bioreactor culture, while simultaneously maintaining a low oxygen tension, could make it possible to engineer larger homogenous cartilage tissues using MSCs. We demonstrate that a simple dynamic culture regime involving continuous translation of channelled MSC laden hydrogels through culture media at a low oxygen tension (3% O2) can enhance the distribution of cartilage matrix components within larger scale tissues compared to static conditions. No benefits of dynamic culture were found when engineering smaller cartilage tissues (5 mm Ø × 3 mm), suggesting that diffusional limitations that arise under static culture do not limit the capacity of MSCs to produce cartilage matrix components at this scale. However, when the constructs were scaled up (10 mm Ø × 6 mm), core regions devoid of cartilage ECM components were observed in static culture conditions. The bioreactor system developed in this study was designed to enhanced nutrient transport, resulting in tissues with a more homogenous distribution of matrix components. The study also further affirms the importance of controlling the external oxygen tension when designing chondrogenic culture protocols, as dynamic culture at 20% O2 was found to completely suppress chondrogenic differentiation of MSCs compared to static conditions. Taken together, these results indicate that dynamic culture regimes with a controlled external oxygen tension can be employed to engineer tissues of a sufficient scale to treat large cartilage defects or to enable their use in endochondral bone regeneration.
The core regions of large, statically cultured samples contained lower levels of cartilage ECM, highlighting the influence of construct scale on the spatial development of cartilage tissues engineered using MSCs. MSCs have been previously shown to produce cartilage matrix in regions of optimal nutrient supply (i.e. the periphery) and nutrient diffusion limitations become exacerbated when engineering larger cartilage tissues.12,13 The lack of matrix accumulation in core region is likely due to the reduced number of cells capable of producing cartilage matrix along with the remaining viable cells in the core lacking sufficient TGF-β3 stimulation 44 and/or insufficient nutrient supply (e.g. glucose) to produce robust cartilage matrix. Cell viability was significantly lower in central regions of large, statically cultured samples with ~60% of cells alive after 28 days of culture (Figure 2(b) and (c)). However, even with ~60% of the cells viable, histological analysis revealed limited matrix accumulation within these regions indicating that these remaining viable cells received insufficient nutrients and/or TGF-β3 stimulation to produce cartilage matrix. Core regions of statically cultured samples likely present a micro-environment of very low oxygen and nutrient availability which is known to lead to cell death and limit the capacity for differentiation with this cell source.11,45,46
A number of bioreactor culture systems such as spinner flask culture, perfusion culture and rotating wall vessels have been previously employed for cartilage tissue engineering. 47 Spinner flask offers a simple approach for enhancing nutrient transport for cartilage tissue engineering; however, the penetration depth of nutrient transport with these systems has been reported to be 1 mm in larger tissue engineered scaffolds. 48 Rotating wall vessels (RWVs) are generally preferred over spinner flasks and orbital shakers are they offer better medium exchange conditions which are more suitable for tissue engineering large constructs. However, in these systems, samples can be damaged when bouncing against the vessel walls since the constructs are unconstrained. 48 Although a number of studies have reported benefits of MSC chondrogenesis compared to static conditions with RWV, 49 a number of studies have also reported suppression compared to static conditions. 50 In our system, the constructs are translated through the cell culture media in a controlled manner which can facilitate constant agitation and replenishment of the cell culture media, as well as facilitating transport of media along the peripheral and channelled free surfaces. Although perfusion-based systems can ensure improved mass transport throughout the core of tissue engineered cartilage samples, 51 these systems are challenging to implement with hydrogel materials; instead, porous interconnected scaffold are required to facilitate media flow into the interior of the construct. In summary, our custom bioreactor system offers a simple yet effective method for dynamically culturing large cell laden hydrogels.
Our dynamic culture regime enhanced the accumulation of cartilage matrix components in larger samples at 3% O2, but suppressed chondrogenesis when the external oxygen level was increased to 20% O2. A low oxygen environment is known to be a potent promoter of the chondrogenic differentiation of MSCs.1,26–29 The dynamic culture regime utilised in this study will lead to enhanced oxygen transport throughout the constructs due to constant agitation of the cell culture media which is passing along peripheral and channelled free surfaces of the hydrogel. Therefore, an oxygen tension close to the external value (either 3% O2 or 20% O2) likely exists at all internal and external free surfaces of the dynamically cultured channelled hydrogels, while lower levels of oxygen develop within statically cultured constructs due to large oxygen transport distances and cellular oxygen consumption at the construct periphery. 14 The higher oxygen levels in dynamically cultured constructs maintained at 20% O2 therefore likely explain the suppression of chondrogenesis in this group, despite the fact that bioreactor culture will also lead to higher levels of glucose and other key regulatory factors within the core regions of engineered tissues. Consistent with the results presented here, a number of studies have reported the suppression of chondrogenic differentiation of MSCs in similar dynamic culture systems at 20% O2.21,22 In contrast, when the external oxygen tension is reduced to 3% O2, significantly higher and more homogenous distributions of sGAG are observed in larger constructs subject to dynamic culture compared to static controls. Here the engineered tissues benefit from enhanced nutrient transport as the local oxygen levels are maintained at the low levels necessary for robust chondrogenesis of MSCs. Our results point to the importance of careful regulation of oxygen availability in bioreactor systems designed to enhance nutrient transport in engineered cartilage tissues.
Although an adequately low oxygen micro-environment along with sufficient nutrient availability supported enhanced sGAG accumulation in large engineered tissues, no increases in collagen accumulation were observed. This indicates that the micro-environmental requirements for the accumulation of sGAG and collagen are not the same. Instead, higher collagen accumulation was observed in static samples cultured at 20% O2. A threshold oxygen tension of 8% O2 has been proposed for articular chondrocytes where an oxygen tension below this threshold supports mainly sGAG synthesis and above which favours collagen more synthesis. 52 The higher collagen accumulation in statically cultured samples at 20% O2 is likely due to the oxygen gradient created in the gels that creates an environment favourable for collagen synthesis. As articular cartilage is a spatially complex tissue, with variations in biochemical composition and organisation through its depth, it may be possible to engineer bioreactor systems that provide gradients in regulatory molecules to enable the development of more articular cartilage-like tissues. 14
Calcium accumulation was low in all groups cultured at 3% O2, with more calcium produced at 20% O2. It has been reported that differentiation at 20% O2 supports a more hypertrophic phenotype which likely explains the higher levels of calcium production observed here.26,53 The highest levels of calcium accumulation were observed in samples dynamically cultured at 20% O2, which histological analysis demonstrated was at the construct periphery. This may be due to the influence of fluid flow at these locations which has previously been shown to upregulate late hypertrophic and osteogenic markers during chondrogenic differentiation of MSCs. 22
In conclusion, this study demonstrates that dynamic culture regimes can be employed to engineer large cartilaginous tissues with a more homogenous distribution of matrix components compared to free swelling conditions. The problem of increased inhomogeneity in cartilage matrix distribution within larger engineered tissues is overcome using the system. The outcome of bioreactor culture is strongly dependent on the choice of the external oxygen tension, indicating that care must be taken when designing dynamic culture regimes for MSCs. Taken together, these results point to the benefits of dynamic culture systems for generating the large cartilage tissues. Such culture systems could facilitate the scaling up of cartilage tissue engineering strategies towards clinically relevant dimensions.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This publication has emanated from research supported by a research grant from Science Foundation Ireland (SFI) under grant number 12/IA/1554 and a European Research Council Starter grant (258463).