
Abstract
Biomimetic microenvironments are key components to successful cell culture and tissue engineering in vitro. One of the most accurate biomimetic microenvironments is that made by the cells themselves. Cell-made microenvironments are most similar to the in vivo state as they are cell-specific and produced by the actual cells which reside in that specific microenvironment. However, cell-made microenvironments have been challenging to re-create in vitro due to the lack of extracellular matrix composition, volume and complexity which are required. By applying macromolecular crowding to current cell culture protocols, cell-made microenvironments, or cell-derived matrices, can be generated at significant rates in vitro. In this review, we will examine the causes and effects of macromolecular crowding and how it has been applied in several in vitro systems including tissue engineering.
Introduction
Macromolecular crowding is a biophysical phenomenon wherein molecules of defined size and charge exist within a milieu, such as the cytoplasm or cell culture media, resulting in the excluded volume effect. 1 The excluded volume effect causes steric hindrance due to a molecule’s inherent property of impenetrability. In a cellular context, cells are naturally crowded compartments while also being surrounded extracellularly by a crowded environment of macromolecules. It was estimated that the cytosolic compartment of Escherichia coli contained approximately 300–400 mg/mL of macromolecules, making it inherently crowded. 2 This property of ‘crowdedness’ has been evolutionarily conserved across species, from bacteria to humans.3,4 Therefore, given the innate crowdedness of a cell 5 and the historically crowded nature of its environment, it would seem relevant to incorporate this cellular property into scientific experiments, and control for this property, similar to that of temperature and pH.
However, macromolecular crowding, while obvious, is underappreciated. 6 Since its first observation in the 1950s, studies on macromolecular crowding or incorporating macromolecular crowding into in vitro experiments have been few and far between due to the scepticism of adding ‘unknown’ variables (macromolecular crowders) to experiments and the general notion of ‘experiments have worked without the macromolecular crowders, so we do not really need it’. In the 1980s, less than 10 reports of macromolecular crowing were published. However, in the 1990s, there was a five-fold increase in macromolecular crowding-centered manuscripts and this pattern was similarly seen in the 2000s. Recently and in the 2010s, a similar trend is being observed, with approximately 450 reports on macromolecular crowding between 2010 and 2017. This exponential increase in reports on macromolecular crowding and its potential applications reflects the growing appreciation and importance of this biophysical phenomenon.
Macromolecular crowding, in general, causes microviscosity and anamolous diffusion. 7 As such, it results in a spectrum of effects, including effects on biochemical reaction rates, protein folding and binding,8,9 oligomerization and micro-compartmentalization. Figure 1 depicts the effect of macromolecular crowding in solution and illustrates how macromolecular crowding causes the excluded volume effect. In the top panel of Figure 1, when a milieu is uncrowded, or devoid of macromolecular crowders, test molecules are able to diffuse randomly. However, in a crowded milieu, in the presence of macromolecular crowders, test molecules are unable to diffuse freely. Of note, macromolecular crowding (or volume exclusion) is different from volume reduction as it was observed that a gradual reduction in cell culture media volume resulted eventually in cell death. 10 To emulate macromolecular crowding in vitro, several inert macromolecular crowders have been successfully studied and their results are summarized in Table 1. Macromolecular crowders could either be charged or neutral, have varying sizes and hydrodynamic radii. For example, Ficoll is neutral while dextran is negatively charged. Furthermore, Ficoll could be either 70 or 400 kDa. Therefore, a suitable macromolecular crowder has to be selected and optimized in experiments.
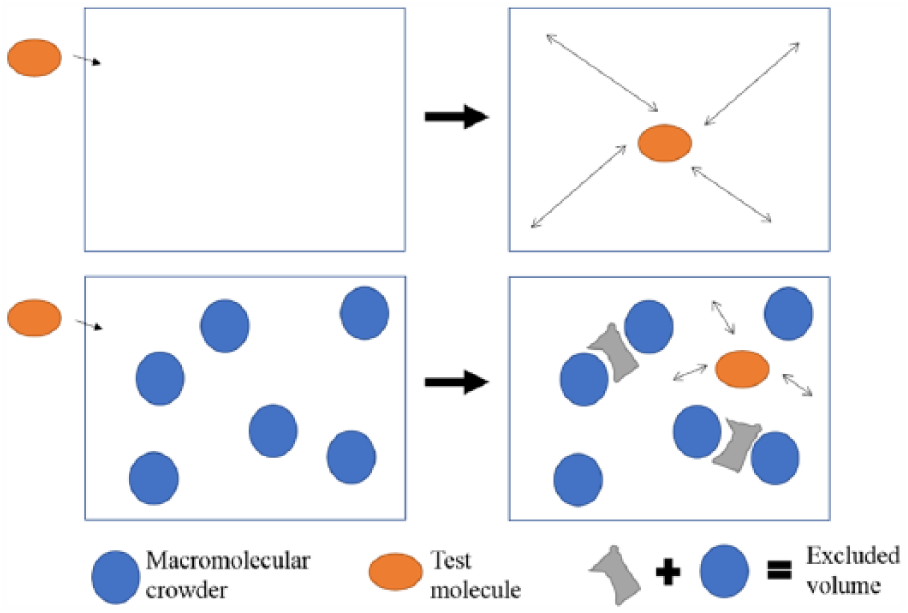
Macromolecular crowding causes the excluded volume effect. In an uncrowded environment, devoid of macromolecular crowders, test molecules freely diffuse in the environment. However, in a crowded environment, where macromolecular crowders are present, test molecules are unable to diffuse freely. The blue circles (macromolecular crowders) plus the grey irregular shape both contribute to the excluded volume effect which is unavailable to the test molecule. Adapted and modified from Minton. 1
List of commonly used macromolecular crowders for in vitro cell culture.

BSA: bovine serum albumin; PEG: polyethylene glycol; PSS: polystyrene sulphonate.
Incorporating macromolecular crowding into typical in vitro experiments would provide a better mimic of the in vivo state, cellular events, interactions and biological processes. 23 For example, macromolecular crowding has been known to cause protein aggregation. In the cell, proteins exist in different states at different times. The evolutionary development of molecular chaperones resulted from the need to maintain flexibility in protein conformation in complex organisms. Molecular chaperones were described to be able to bind temporarily to hydrophobic regions of protein surfaces to prevent protein aggregation. Molecular chaperones were thus attributed to the innate crowdedness of the cell and therefore help proteins maintain its conformation and stability even within the crowded cellular environment. 24 Therefore, without taking into account the effect of macromolecular crowding, a key cause and function of chaperones may not have been identified. Extending this concept to other cellular processes, macromolecular crowding can be viewed as a ‘discovery tool’ to identify processes or functions which may otherwise have previously been overlooked. Banerjee et al. 25 discovered a hidden conformation of α-synuclein, while Shirai and Kikuchi 26 reported the ability of macromolecular crowding to promote α-synuclein fibril formation, a key component in Lewy bodies which are hallmark of Parkinson’s disease. In the first report, Banerjee et al. used macromolecular crowding to study the different conformations α-synuclein could adopt, expanding on the current literature which did not take macromolecular crowding into account. In the second report, Shirai et al. found that α-synuclein fibril formation was enhanced with macromolecular crowding and also with increased α-synuclein concentration. Both reports contribute to the understanding of the cellular mechanisms underlying Parkinson’s disease progression, through α-synuclein formation and accumulation. Zinchenko 27 also highlighted the importance of macromolecular crowding in understanding DNA packaging in cells. The state of crowdedness within the nucleus is therefore highly related to the gene expression profile of a cell. In enzyme kinetics, 28 macromolecular crowding impacts on reaction rates, for example, macromolecular crowding has been reported to increase the reaction rates of the RNA enzyme, ribozyme, 29 which has immense implications in RNA evolutionary biology. In addition, macromolecular crowding has recently been reported to affect biological reactions occurring on cell surfaces. Chapanian et al. 30 used a system of blood type conversion of red blood cells where selective glycosidases remove antigen carbohydrates, located on the cell surface, to convert red blood cells from blood type A or B to universal type O. This enzymatic reaction was magnified 400-fold in the presence of macromolecular crowders.
Macromolecular crowding plays key roles in cell signalling, actomyosin kinetics, 31 calcineurin behaviour 32 and as an activator of heat-shock-protein-90. 33 In protein dynamics, macromolecular crowding has been reported to affect protein assembly and disassembly. 34 Luo et al. 35 discussed the increase in β2-microglobulin fibril formation by macromolecular crowding as it favours the nucleation step while inhibiting fibril disassembly. Rosin et al. 36 reported the increase, in the order of magnitude, in actin polymerization when macromolecular crowders such as dextran were added to solutes.
Macromolecular crowding has been reported to have effects on the DNA, RNA and protein level. On the DNA level, Golkaram et al. 37 and others38–40 explained that macromolecular crowding regulated gene expression by limiting diffusion. By measuring the kurtosis of mRNA diffusion, it was observed that macromolecular crowding reduced ‘noise’ in a cell population by limiting the diffusion of transcription factors. Transcription factors remained on promoters longer thereby removing unstable intermediate states. Furthermore, the size of the macromolecular crowder had an impact on the ability to reduce ‘noise’. Larger sized macromolecules were more effective as compared to smaller sized macromolecules. In another study, Harve et al. 41 discussed how macromolecular crowding stabilized hydrogen bonding between nucleotides thereby stabilizing DNA/DNA and DNA/RNA hybrids. Even applicable to virus studies, Smith et al. 42 have applied macromolecular crowding to simulate virus capsid assembly in vitro. In prokaryotic systems, Nolan et al. 43 showed that with macromolecular crowding, E.coli β-galactosidase acquired a relaxed conformation and higher thermal stability resulting in lower catalytic efficiency. In addition, Tyrrell et al. 44 reported that polyethylene glycol (PEG)-induced macromolecular crowding, in particular high-molecular mass PEG, increased ligand affinity and favoured compact RNA states, thus moving scientific observations towards more in vivo-like conditions.
Applications of macromolecular crowding
Enhancing extracellular matrix deposition in vitro
In this review, particular focus is on the extracellular matrix, which is the structural element holding cells together to form tissues and represents a unique and specific microenvironment for cells. 45 The extracellular matrix is composed primarily of collagen and to a lesser extent of fibronectin, laminin, glycosaminoglycans, proteoglycans and is organ-specific.46,47 Collagen exists in over 20 different subtypes, with Collagen I being the most common. Collagen I is the primary structural component of the extracellular matrix of a cell and is frequently used as scaffolding material or as a coating to improve cell culture in vitro. However, collagen production and deposition in vitro is notoriously slow due to the enzymatically rate-limited reaction of pro-peptide cleavage of procollagen, resulting in minimal non-detectable amounts of functional collagen in vitro. With macro-molecular crowding, Lareu et al. 48 reported an increase in collagen deposition when fibroblasts were grown under crowded conditions as compared to control cultures. This was shown to be due to an increase in propeptide-C-proteinase enhancer protein under macromolecular crowding which explains the increase in collagen deposition.
While macromolecular crowding has been useful in understanding cell biology, it has also been useful in improving on current in vitro experiments. Smaller amounts of reagents are needed for the same experiment and time-frames were condensed in experimental set-ups. Lareu et al. 48 showed that the incubation time for collagen deposition was reduced from 4 weeks to 2 days using 100 µg/mL of macromolecular crowder dextran sulphate on the same number of seeded cells. In key experiments, such as the polymerase chain reaction, the addition of a small amount of macromolecular crowders to reactions was sufficient to enhance polymerase processivity, promote higher specific amplicon yield, primer annealing, as well as DNA polymerase thermal stability. 49 Incorporating macromolecular crowding into cell culture resulted in cost-effective experiments. This effect was also mirrored in industrial processes, for example, wherein the addition of macromolecular crowding to dummy templates allowed for faster purification of punicalagin from pomegranate husk extract. 50 In addition, the authors reported the possibility of minimizing the amount of template material used in preparations of molecularly imprinted monolith. In instances where the template material is rare, using the polystyrene-based macromolecular crowder proved beneficial. Macromolecular crowding has also been reported to be used in the fabrication of liquid crystalline imprinter polymers. 51 In food chemistry, Perusko et al. 52 applied macromolecular crowding, using PEG, to enhance oxidation and glycation of whey proteins which resulted in the shortening of processing time, milder conditions, while preserving the structure of proteins as well as reducing the formation of protein aggregates.
In the field of bioengineering and tissue regeneration, macromolecular crowding has been shown to successfully promote the cell-specific production and deposition of extracellular matrix in vitro. In two-dimensional (2D) monolayer models, several groups have reported enhanced extracellular matrix deposition as compared to uncrowded controls, including reports in human corneal fibroblast cultures,53,54 skin cell cultures, 55 human dermal fibroblast culture,56,57 bone marrow stroma, 58 vocal fold lamina propria tissue, 59 mesenchymal stem cell culture 60 and cartilage. 61 Of note, the extracellular matrix produced under macromolecular crowding is distinctly cell-specific. Macromolecular crowding maintains the specific complexity and composition associated with the cell type.
Extrapolating the observations seen in 2D monolayer cultures, several groups endeavoured to apply macromolecular crowding to three-dimensional (3D) organotypic cultures, which is a more accurate representation of the in vivo environment. The effort to obtain more clinically relevant tissues, rich in extracellular matrix, produced by the cells themselves, was successfully shown in several tissue types, including corneal stroma supramolecular assemblies 42 and skin organotypic co-cultures. 62 These tissues could subsequently be used as 3D in vitro models or as tissue grafts. In addition, Dewavrin et al. 63 reported that in 3D models, macromolecular crowding was able to tune 3D architectures of collagen hydrogels in terms of collagen nucleation and fibre diameter, organization and growth. The gels generated under these ‘nature-inspired’ conditions were more resistant to mechanical stress as compared to those generated without macromolecular crowders. Enhancing extracellular matrix in vitro allows, for instance, the rapid production of collagen. Large amounts of collagen, produced by cells themselves and in a short time-frame, provide a screening platform for therapeutics. For example, Chen and colleagues18,64 developed the Scar-In-A-Jar model which allows for the rapid screening of anti-fibrotic agents. This screening platform was so named because scar tissue consists primarily of collagen. Therefore, a large amount of collagen deposited in a culture dish was akin to a scar formed in a dish, or a scar formed in a ‘jar’. In addition, Chen et al. 65 succinctly summarized the broad applications of macromolecular crowding for cell-based therapies.
Making microenvironments
Generation of cell-specific microenvironments
The benefits of rapidly producing large amounts of extracellular matrix are multi-fold. In addition to generating screening platforms for drug and toxicity assays, macromolecular crowding also allows for the generation of cell-specific microenvironments. Microenvironments play a key role in cell expansion in vitro, especially for stem cell culture. Current in vitro microenvironments for stem cell culture comprise usually of feeder cell layers or a single extracellular matrix coating, such as collagen or fibronectin on either glass or plastic surfaces. Only a few coatings comprise mixtures of extracellular matrix components; however, these tend to be of xenogeneic origin. A current challenge is to create xenogeneic-free cell-specific microenvironments for improved in vitro cell culture systems.
A major field and focus in tissue engineering is making matrices and biomimetic microenvironments. The current state of the art is either to coat surfaces with a commercially available extracellular matrix, or to construct matrices in vitro. Matrices could either be cell-free or still contain cells, which could either be living or devitalized. Cell-derived matrices, a term given to decellularized extracellular matrix, has been gaining importance as it is an exclusively cell-made matrix. Decellularization essentially refers to the removal of the cells, thus leaving behind the extracellular matrix constructed by those cells. Several decellularization methods can be used to obtain a cell-derived matrix, including chemical agents such as ammonium hydroxide, 66 Triton-X, trypsin-Ethylenediaminetetraacetic acid (EDTA), (CHAPS), sodium dodecyl sulphate, 67 desiccation or lyophilization, 68 biological detergents such as sodium deoxycholate,47,69 phospholipase A2 70 and snap freezing. 71 In addition, these methods can be applied to 2D monolayers or 3D tissues, though for 3D constructs, permeation of specific tissues has to be optimized first.
In 2D monolayer cultures, few protocols incorporate macromolecular crowding into the initial construction process of the extracellular matrix, thus resulting in already low amounts of deposited extracellular matrix and after subsequent decellularization, there is only a very thin layer of extracellular matrix remaining. Applying macromolecular crowding to the initial construction and deposition phase of extracellular matrix produces a large volume of cell-made extracellular matrix, which is still significant after the decellularization process. A variety of macromolecular crowders have been identified which have been shown to promote deposition of extracellular matrix by the cells themselves, including dextrans, ficolls, PEG, polystyrene and polyvinylpyrrolidone (PVP). 72
Biomimetic microenvironments generated with macromolecular crowding
Incorporating macromolecular crowding into this research effort has produced several cell-specific microenvironments with an array of functions. Ang et al. 73 reported the ability of macromolecular crowding to hasten adipogenic induction protocols to differentiate bone marrow–derived mesenchymal stem cells into adipocytes. Furthermore, extracellular matrix generated from differentiated cells (under macromolecular crowding; adipo-Extracellular Matrix [ECM]) maintained adipogenic cues and contained adipocyte lineage inducing properties. Mesenchymal stem cells subsequently seeded on adipo-ECM differentiated into the adipogenic lineage significantly faster, as compared to tissue culture plastic. This was evidenced by significant upregulation of early and late adipogenic markers expressed by differentiated mesenchymal stem cells seeded on adipo-ECM. In addition, these cells also had a 60% higher lipid content. Microenvironments generated under macromolecular crowding were also able to promote cell signalling. Lee et al. 74 discussed the ability of extracellular matrices generated under macromolecular crowding to encapsulate maturing adipocytes within a collagen IV-enriched cocoon and which was observed to interact with paxillin-positive focal adhesions. This matrix-cell signalling is apparent with the enhanced phosphorylation of activating transcription factor 2 (ATF2) which is a major transcription factor in uncoupling protein 1 (UCP1) regulation, thus illustrating the importance and capability of microenvironments in directing cell fate and cell differentiation.
Biomimetic microenvironments are also important in cell population expansion. Peng et al. 75 reported the successful expansion of H9 human embryonic stem cells for up to 20 passages using extracellular matrices generated by fibroblasts cultured together with macromolecular crowders as compared to matrigel. The expanded cells maintained its differentiation capacity and teratoma formation and were suitable for clinical applications. Rashid et al. 76 reported an increase in proliferation rates of fibroblasts and bone marrow–derived mesenchymal stem cells using a novel macromolecular crowder, PVP. PVP 360 kDa was able to attain fractional volume occupancy of full serum, which is 54%, without changing the viscosity of the solution. Therefore, a panel of macromolecular crowders exist which could tune the viscosity of solutions to mimic cellular or extracellular compartments which would be organ-specific. In addition, Dewavrin et al. 77 noted a 70% boost in stem cell proliferation when cells were grown on tuned soft hydrogels generated with macromolecular crowding. This opens up an untapped avenue for stem cell culture and expansion as current challenges in stem cell culture are largely due to the spontaneous differentiation and senescence of stem cells when grown in vitro.
Interestingly, the culture of cells, for example, mesenchymal stem cells, in the presence of macromolecular crowders was already sufficient to affect cell proliferation, adhesion and migration due to the unique supramolecular assembly and alignment of the extracellular matrix generated with macromolecular crowders. Zeiger et al. 78 explained that the increase in cell proliferation and decrease in cell motility were due to the enhanced extracellular matrix deposition and how it in turn had an effect on the intracellular actin cytoskeleton which thus affected cell proliferation and migration. This highlights the importance of cell-matrix reciprocity and the importance of biomaterial characteristics and microenvironments.
Conclusion
Macromolecular crowding is an important cell property which should be accounted for in in-vitro models to more accurately mimic the in vivo state, resulting in translationally relevant results. While debates exist on the ‘friend or foe’ nature of macromolecular crowding, 79 the benefit of applying macromolecular crowding to scientific and technological endeavours is immense, extending from discoveries of cellular biological functions, clinical applications of primary cell expansion in vitro, to improving on cost-effective protocols in industrial processes, and the implications and possibilities of applying macromolecular crowding are still being unravelled.
In constructing biomimetic microenvironments in vitro, incorporating macromolecular crowding into experiments takes into account the crowded nature of cells and cellular compartments and provides more accurate and efficient in vitro models. The success of stem cell culture in vitro hinges on a fine balance of growth factors, media composition, signalling cues and structural properties of the extracellular matrix. It is evident that incorporating macromolecular crowding into current cell culture protocols provides a beneficial enhancement to current procedures. Applying macromolecular crowding to improve the cells’ innate ability to produce and deposit extracellular matrix allows for the generation of biomimetic matrices which can be used as a stem cell niche, as well as for other tissue engineering purposes.
While there exists several widely used macromolecular crowders, including neutral and negatively charged carbohydrates, studies are on-going in search of even more macromolecules which can function as crowders. Sharp 80 and other groups 81 have reported the size dependence of macromolecular crowding, concluding that generally ‘smaller is better’. In addition, other groups are developing sensors to more accurately quantify macromolecular crowding in living cells.82,83 Of interest, certain groups have used macromolecular crowding to create heterogeneous microenvironments in picoliter droplets to study specific mechanisms of gene expression in vitro. 84 Currently, in silico models provide foresight and predictions to even more functions, causes, effects and uses of macromolecular crowding, including DNA structures, 85 protein structures, 86 reaction rates 87 and the very basics of biochemical reactions which govern cells, moving towards the creation and study of artificial cells. 26 These studies are highly relevant in the continuous quest of scientific discoveries, technological improvement and medical advances.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by Swiss Biotechnet.