
Abstract
Dipeptidyl peptidase IV (DPP-4), an incretin glucagon-like peptide-1 (GLP-1) degrading enzyme, contains two forms and it can exert various physiological functions particular in controlling blood glucose through the action of GLP-1. In diabetic use, the DPP-4 inhibitor can block the DDP-4 to attenuate GLP-1 degradation and prolong GLP-1 its action and sensitize insulin activity for the purpose of lowering blood glucose. Nonetheless the adverse effects of DPP-4 inhibitors severely hinder their clinical applications, and notably there is a clinical demand for novel DPP-4 inhibitors from various sources including chemical synthesis, herbs, and plants with fewer side effects. In this review, we highlight various strategies, namely computational biology (in silico), in vitro enzymatic and cell assays, and in vivo animal tests, for seeking natural DPP-4 inhibitors from botanic sources including herbs and plants. The pros and cons of all approaches for new inhibitor candidates or hits will be under discussion.
Introduction
According to the statistics from International Diabetes Federation (IDF), there were 425 million diabetes mellitus (DM) patients in 2017 worldwide, and that number is expected to increase to 629 million by 2045. 1 Abnormally high blood glucose caused by insulin insufficiency or insensitivity can lead to severe complications such as chronic renal failure, microvascular complication, cerebrovascular accident, and infarction induced by high glycated serum and blood vessel proteins.2,3 Moreover, insufficient insulin signal leads to the decreased glucose uptake from the blood that, in turn, can result in ulcers, gangrene, diabetic retinopathy, and neuropathy. 2 Recent DM treatment is inclined to maintain the blood glucose level within normal limits by (e.g. nutritional therapy and physical management) and medication due to the incurable nature of DM.4,5 Diabetic medications can be characterized into five strategies based on their acting mechanisms: raising insulin secretion (e.g. sulfonylurea and meglitinide analogs), reducing intestinal glucose absorption (e.g. acarbose), triggering insulin-independent glucose uptake signaling (e.g. thiazolidinedione and biguanide), reducing urinal glucose reabsorption (e.g. gliflozins), and prolonging insulin sensitive [e.g. dipeptidyl peptidase 4 (DPP-4) inhibitor and glucagon-like peptide 1 (GLP-1) receptor agonists] [American Diabetes Association, 2019b]. These antidiabetic drugs help DM patients to maintain their blood glucose levels with various adverse effects (Table 1), such as urine-tract infection, lactoacidosis, hypoglycemia, and obesity. These drug-related adverse effects can deteriorate the quality of life of DM patients and create unsurmountable difficulties for proper dosing regimens in a clinical setting. It has been observed that DPP-4 inhibitors can exert a similar efficacy in reducing blood glucose levels without severe adverse effects such as hypoglycemia as compared with sulfonylurea. 6 Nevertheless, various adverse side effects associated with the current DPP-4 inhibitor can still be observed that, in turn, can severely limit their practical application. As such, there is a clinical demand for novel DPP-4 inhibitors from various sources including chemical synthesis and botanic sources containing herbs and plants with fewer side effects. In this article, we have reviewed various approaches including in silico, in vitro enzymatic and cell assays, and in vivo animal tests in the search for natural DPP-4 inhibitors for the treatment of type 2 diabetes.
Current hypoglycemic agents and their side effect. 7

Overview of DPP-4 and its biological function
Two forms of DPP-4
DPP-4, which is a 88 kDa serine protease, contains one region of cytoplasmic region (amino acids 1–6) coupled with transmembrane domain (amino acids 7–28) and extracellular region (amino acids 29–766) with the main catalytic domain. 14 There are two DPP-4 isoforms in the body: membrane-bound DPP-4 (mDPP-4) composed of full-length DPP-4 peptide; and soluble DPP-4 (sDPP-4), whose cytoplasmic and transmembrane regions are absent. 14 Both forms can exert various biological activities in regulation of physiology and pathology. 15
Biological function of soluble form DPP-4
sDPP-4 is secreted by lymphocytes, circulates in the blood, 16 and shows high concentration in kidney. 17 It has been observed that sDPP-4 plays various roles in improving skeleton muscle activity, immunocyte activation, chemotaxis, and homeostasis. sDPP-4 can secrete into serum via the response of skeletal muscle cells upon acute physical activities or feeding protein hydrolysate. 18 Secreted sDPP-4 can reduce vasoconstriction that is caused by neuropeptide Y (NPY) and subsequently increase the arteriolar diameter of skeletal muscle that provides a physiological explanation for raising training efficiency caused by sDPP-4.19,20 In addition to arteriolar diameter of skeletal muscle, secreted sDPP-4 acts as myokine, which stimulates inflammation in smooth muscles from blood vessel through activating protease-activated receptor 2 (PAR2)/ERK/NF-κB signaling pathway, increasing proinflammatory cytokine release and finally stimulating smooth muscle cell proliferation. 21 However, sDPP-4-induced smooth muscle inflammation is not always good to the body. For instance, Romacho et al. reported sDPP-4 might cause microvascular endothelial dysfunction, which is the cause of chronic kidney disease in elders, through the same signaling with smooth muscle inflammation.22,23 Thus, Dubé et al. illustrated that cardiovascular inflammation can be attenuated by DPP-4 inhibitor in the process of HIV treatment, and that leads to diminish cardiovascular morbidity of HIV treatment. 24 In T-cell activation, sDPP-4 can activate T-cell proliferation via co-stimulation with T-cell receptor (TCR) signaling and Toll-like receptor, whose activation is neither associated with its enzymatic activity nor with adenosine deaminase binding.25–28 On the other hand, sDPP-4 can upregulate the expressions of IL-6 and TNF-α in monocyte through caveolin-1/ERK/NF-κB/c-Fos signaling, which is involved in monocyte proliferation. 29 It is noteworthy that Tansi et al. proved the interaction between DPP-4 and HIV-1 transcription regulator Tat (HIV-1-Tat). 30 Markedably, the effect on DPP-4/HIV-1-Tat in viral infection and proliferation needs further investigative attention. The role of sDPP-4 in chemotaxis regulation is related to the degradation of chemokine. Hematopoietic stem cell homing is attracted by stromal cell-derived factor 1 (SDF-1/CXCL12) and colony-stimulating factors (CSFs), which are substrates of both sDPP-4 and mDPP4.15,31 Accordingly, sDPP-4 inhibition can improve success rate of transplantation after analyzing the relationship between sDPP-4 activity and hematopoietic stem cell transplantation. 32 The known DPP-4 (both soluble and membrane-bound forms) substrates include numerous homeostatic hormones such as GLP-1, NPY, glucagon, peptide Y, and secretin, which accurately regulate blood sugar homeostasis. 15 GLP-1 is involved in insulin sensitivity and secretion, food reward, and appetite through ghrelin and leptin, and cellular metabolism with adiponectin.15,33–35 NPY blocks melanocortin-4 receptor signaling and leads to the reduction of energy consumption and possibility of obesity.36,37 After secretion from intestinal L cell, GLP-1 is rapidly degraded by sDPP-4 into inactive GLP-1 amide, in which half-life is shorter than 2 min. 38 In dysglycemic patients, active GLP-1 content can be further decreased and cause more severe hyperglycemia. 39 Accordingly, inhibiting sDPP-4 activity can keep more active GLP-1 and NPY in serum and thus improve insulin efficiency. 40 Other biological activities including nonimmunodeficient virus infection are also explored. It has been demonstrated by the case study of Middle East respiratory syndrome coronavirus (MERS-CoV) infection that sDPP-4 can potentially function to block viral infection.41,42 Truncated C-X-C chemokine 10 (CXCL10) secretion in persistent infection of hepatitis C is essential, 43 implying that DPP-4 may play a role in HCV persistent infection as manifested by a case report, in which a DM patient complicated with HCV infection showed HCV replication reduction after sitagliptin treatment. 44 sDPP-4 is essential for maintaining immune activity and chemotaxis, especially in inflammatory regulation, based on all available information of its biological activities. Thus, the serum activity of sDPP-4 can be an indicator of physiological or immunological stages.
The effect of serum sDPP-4 activity can be classified into several categories: infection related damage, transplantation or autoimmune disease, respiratory disease, and response to diabetic medications and complications, as discussed in detail in the following. Serum sDPP-4 activity in HIV infection is referred to the HIV-induced intestinal damage that is caused by Th17 cell depletion.45,46 It has been observed that rheumatoid arthritis and multiple sclerosis patients have lower serum sDPP-4 activity than healthy people,47,48 whereas HIV patients have higher serum sDPP-4 activity. 49 Moreover, Leicht et al. analyzed sDPP-4 activity in end-stage renal disease patients before and after kidney transplantation, and sDPP-4 activities were found to decrease after kidney transplantation. 50 These results suggest that serum sDPP-4 activity can be a potential biomarker for monitoring the progress of autoimmune disease and the prognosis of organ transplantation. Interestingly, serum sDPP-4 activity can be associated with the progress of chronic obstructive pulmonary disease (COPD), 51 which is highly correlated with respiratory inflammation, obviously indicating the relationship between serum sDPP-4 activity and COPD progression. 52 However, the prognosis of malignant pleural mesothelioma (MPM), which is a rare pulmonary malignancy, can be predicted by sDPP-4 activity in pleural fluid. 53 The pathological correlation between sDPP-4 activity and MPM prognosis is still veiled. These reports provide interesting suggestions about sDPP-4 levels as the biomarkers of various diseases, which are not easy to monitor in serum or other body fluid.
Biological function of membrane-bound DPP-4
Remarkably, mDPP-4 can be found mainly in the kidney, gastrointestinal tract, T lymphocytes, and reproductive organs.54,55 Biological activities of mDPP-4 include the regulation of immune response and blood vessel function. 15 mDPP4, also named CD26, is a T-cell co-stimulator of T-cell receptor responding to antigen-presenting cells. 56 Thus, mDPP-4 recently has been considered as a potent target in treatment of transplantation and autoimmune disease. Dolanbay et al. reported an interesting study about the impact of mDPP-4 inhibition in early pregnancy that can be important in treating recurrent implantation failure. 57 In hematopoietic stem cell transplantation, graft-versus-host disease (GVHD) is a common complication, which is critical in survival rate after transplantation. 58 Zhang et al. proved the association between Th17 cells and GVHD and which Th17 cell can be regulated by mDPP-4 inhibition that indicates current DPP-4 inhibitors can help ameliorating the onset of GVHD. 59 The impact of mDPP-4 in immune regulation is also implicated in autoimmune disease and hypersensitivity. mDPP-4 expression levels in CD8+ T cells of Hashimoto’s thyroiditis patients are significantly lower than healthy subjects that is plausibly attributed to disease progression. 60 In contrast, mDPP-4 levels are almost 11-fold higher in psoriatic skin than in normal skin, asserting the involvement of mDPP-4 in psoriatic development. 61 mDPP-4 plays a positive role in asthma progress through promoting T-cell activation. 62 These reports have unequivocally demonstrated the positive role played by mDPP-4 in immune regulation. mDPP-4 acts on both endothelial and epithelial cells in regulation of blood vessel function,. In addition to endothelial inflammation caused by sDPP-4, the role of mDPP-4 in endothelial generally involves the endothelial migration, angiogenesis, and proliferation under hypoxia status, which can be found in the development of endometriosis. 63 Xu et al. have pointed out that DPP-4 inhibitors that can alleviate pulmonary artery remodeling and, finally, delay the development of pulmonary hypertension. 64 In the regulation of cardiovascular function, mDPP-4 inhibition can reverse diastolic left ventricular dysfunction via inhibiting mDPP-4/SDF-1α related angiogenesis. 65 mDPP-4 involves in epithelial-mesenchymal transition (EMT) for epithelial cell, 66 suggesting the potential implication of mDDP-4 in promoting cancer development. In fact, breast cancer metastasis can be triggered by DPP4 inhibition through CXCL12/CXCR4/mTOR pathway. 67 However, DPP-4 inhibitor shows the opposite activity in non-small cell lung cancer, which suppresses cancer cell growth via macrophage-mediated natural killer (NK) cell activation. 68 Collectively, these studies have furnished comprehensive descriptions of mDPP-4 biological functions in the whole body and further manifest that DPP-4 inhibition (sDPP-4 or mDPP-4) can produce unexpected side effects.
Role of DPP-4 in diabetes treatment
The endocrinological impact of DPP-4 is more prominent in the mediation of blood glucose. DPP-4 inhibition is a predominant approach for treating diabetes because of prolonged incretin half-lives within serum, especially in type 2 DM. 69 In addition, it has been suggested that sitagliptin can preserve pancreatic β-cell function and subsequently stabilize insulin secretion as shown by two 4-year clinical trials, in which sitagliptin was adopted to treat slowly progressive type 1 DM (SPTIDDM) and latent autoimmune diabetes adult (LADA).55,70 In addition to clinical treatment, DPP-4 levels can be used as a biomarker. For instance, high serum sDPP-4 levels can be referred to the elevated glycation end products, which subsequently evoke endothelial cell damage and diabetic nephropathy incidence.71,72 In addition, high serum sDPP-4 levels also indicate worse drug response to DPP-4 inhibitor and hyperglycemia, which are the indicators of poor glycemic control and advanced disease progress.73,74 The above information emphasizes the effect of DPP-4 inhibition and monitoring in DM treatment. The method of screening DPP-4 inhibitor and the recent known natural DPP-4 inhibitors are presented in the following.
Methods for screening novel DPP-4 inhibitors
In silico screening of DPP-4 inhibitors
Virtual screening has been seamlessly integrated into drug discovery and development 75 and its success significantly relies on compound library, 76 especially the structural diversity of compound library. 77 For instance, microalgal metabolites were screened for DDP-4 inhibitors. 78 Compared with the synthesized chemicals, natural compound libraries generally consist of more structurally diverse compounds than their synthetic counterparts, 79 providing a better screening resource. As such, numerous studies have adopted various natural compound libraries to find novel DDP-4 inhibitors as listed in Table 2.
Natural compound libraries adopted by various studies.

It is not uncommon to observe that docking studies were carried out based on a single DDP-4 crystal structure, despite the fact that a great number of DDP-4-inhibitor co-complex structures have been deposited in the Protein Data Bank (PDB). Deng et al. 88 for instance, docked a series of synthesized triazole-based uracil derivatives into the linagliptin-DDP4 co-complex structure (PDB code: 2RGU) using the standard precision (SP) Glide (Schrödinger, Inc.), which places internally generated ligand conformations with various positions and orientations into the binding pocket. Deng et al. 89 employed Gold (Cambridge Crystallographic Data Center), which is a genetic algorithm (GA)-based scheme to explore the conformational flexibility of ligand and the rotational flexibility of receptor, to dock synthesized pyrazolo inhibitors into the quinazolinone-DDP4 co-complex structure (PDB code: 2ONC). 90 It should be noted that both Glide and Gold are flexible docking algorithms. 91 Nevertheless, DDP-4 is unrestrained per se as manifested by the fact that DDP-4 consists of various binding subsites, namely S1, S’1, S2, S’2, and extensive S2, etc., to which the corresponding amino acids of the DDP4 substrate peptide designated by P1, P’1, P2, P’2, etc. from the nearest to the farthest cleavage point can bind as shown in Figure 1, 92 and S2, which is composed of various hydrophobic residues, namely GLU205 and GLU206 dyad and ARG125, is highly plastic. 93 Furthermore, Nabeno et al. categorized inhibitors into three different classes (Figure 1) based on the interactions between inhibitor and DDP-4 subsites as listed in Table 3, from which it can be observed that inhibitors of different classes bind to different DDP-4 subsites, and S1 and S2 are the common binding subsites. 92 In fact, the root means square deviation (RMSD) between alogliptin-bound protein conformation (PDB: 3G0B) and teneligliptin-bound one (PDB: 3VJK) is 1.07 Å, denoting the promiscuous nature of DDP-4 that can be further demonstrated by Figure 2, in which the six DDP4 co-complex structures (PDB codes are 3W2T, 1X70, 3BJM, 2RGU, 3VJK, and 3G0B) are superimposed. Moreover, the diverse structures of inhibitors can also manifest the plasticity of DPP4. Lee et al. for instance, analyzed the chemical structures of those launched DDP-4 inhibitors and divided them into different classes 94 that, in turn, can be further noticeable the promiscuity of target protein. 95 As such, the promiscuity of DDP-4 cannot be fully taken into account unless more sophisticated structure-based ensemble docking schemes such as SVM-Pose/SVM-Score combinatorial ensemble docking 96 or analog-based pharmacophore ensemble schemes such as pharmacophore ensemble/support vector machine 97 can be adopted. It can be argued that molecular dynamics (MD) can be used to address the flexibility of DDP4 as illustrated by the study of Liu et al. 98 Nevertheless, the lengthy MD calculation will substantially increase computational time and expense, making it impractical to be carried out in a high-throughput fashion, let alone the more resource-demanded quantum mechanical (QM)/molecular mechanics (MM) algorithm. 99

Nomenclature of substrate inhibitor residues and their corresponding subsites in the binding pocket of enzyme and the concept of three classes of inhibitors based on their subsites.
Three classes of inhibitors based on their subsites, their corresponding Protein Data Bank (PDB) entry, and references.

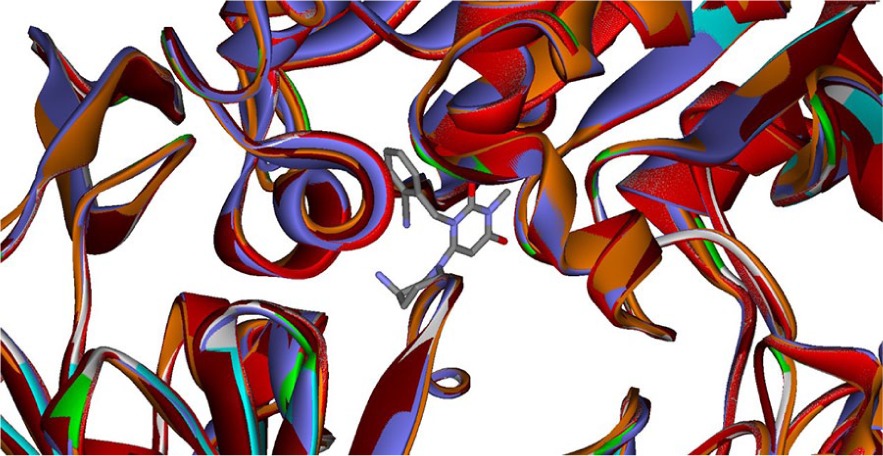
The superposition of six DDP4 co-complex structures, whose PDB codes are 3W2T (color-coded in green), 1X70 (gray), 3BJM (purple), 2RGU (orange), 3VJK (brown), and 3G0B (red). The ligand alogliptin is shown in colors.
Direct test of compounds against DPP-4
There are four types of assay methods for screening DPP-4 inhibitors for direct testing: direct enzymatic assay, in vitro cell assay, ex vivo assay, and in vivo animal tests. DPP-4 and tested compounds are mixed in the direct enzymatic assay, followed by adding specific substrate peptides such as gly-pro-p-nitroanilide. The chemical p-nitroanilide will be released from peptides and the amount is determined by optical absorption at 405 nm at noninhibition state. 105 This method is fast for analysis and can be used to evaluate the inhibition pattern from calculated Ki values. However, the minimum changes within direct enzymatic assay cannot be directly translated into the actual bioactivity in cells and animals. 106 Ex vivo assay can simulate the biological interaction within body, whereas it needs fresh serum or tissue sample as the source of DPP-4.107,108 Moreover, previous studies reported that mucosal DPP-4 inhibition can be possibly related to the onset of coeliac disease, which is an autoimmune disorder due to the immune response to gluten.109,110 However, a modern version of DPP4 activity assay needs to homogenize the whole intestinal biopsy that, in turn, can lead to mucosal DPP-4 inhibition. 108 Yazbeck et al. have derived a new DPP4 substrate with 13C isotope that can be released upon reaction with DPP-4. 111 As such, the requirement to homogenize the biopsy is completely exonerated, leading to higher correlation as compared with its conventional counterparts. 111
Myocytes and pancreatic cells are often used in the cell-based assays to discover DPP-4 inhibitors. Because β-cells in pancreatic islet is an important GLP-1 target, the downstream signaling of GLP-1 in pancreatic cells can be an indicator or biomarker of DPP-4 activity. 112 In addition, GLP-1 attenuates lipopolysaccharide (LPS)-induced cardiomyocyte inflammation. The variations of inflammatory signaling including NF-κB, ERK, and TNF-α within LPS-induced cardiomyocyte can indirectly gauge DPP-4 activity. 113 Nevertheless, the results of cell-based assay can be an authentic representative of the realistic situation mainly due to the fact that they do not consider in vivo pharmacodynamic and pharmacokinetic factors. Nevertheless, direct action upon target cells can be very helpful in detailing intracellular dynamics prior to clinical or animal tests.
DPP-4 inhibitors have been highlighted as potential regimen for autoimmune-disease based on the characteristics in T-cell activation and inflammation. 55 Notably, autoimmune animal model becomes a platform for testing in vivo efficacy of DPP-4 inhibitors in long-term administration. 114 Alternatively, in vivo assay of DPP-4 inhibitory efficacy can be verified by diabetic animal model despite the fact that DPP-4 can degrade GLP-1, leading to insulin desensitization and secretion decrease. 112 The most unvanquished limitation of in vivo test is that only end-point effect can be observed in pre-testing drug candidate despite the fact that it is more related to clinical situations. The data retrieved from in vitro and direct enzymatic assay can be synergistically essential for understanding the conceivable hypoglycemic mechanism.
Natural compounds as novel DPP-4 inhibitors
The effects of DPP-4 studies were mainly focused on immune, endocrine, and neuron system from the end of 1990s to early of 2000s.115–117 A study reported in 2006 that incretin was the molecular target of DPP-4, suggesting the implication of DPP-4 inhibitor in diabetes treatment. Consequently, the diabetic research has turned into a new paradigm for searching for antidiabetic DPP-4 inhibitors. 118 To date, only very limited natural DPP-4 inhibitors from various sources/origins have been reported (Table 4). Natural DPP-4 inhibitors from different origins using different approaches for screening the compound to reach the target are summarized as follows. In addition to plant source, DPP-4 inhibitors from animals and microbes are single-subclasses, in which DPP-4 inhibitors from animals and microbes are peptides and macrolides, respectively. Interestingly, the most predominant subclasses of DPP-4 inhibitors are terpenoids, peptides, phenolics, and flavonoids. These findings have implicated that alkaloids are not suitable as DPP-4 inhibitors or their applications in DDP4 inhibition have not been well explored. In addition to pure compounds, some crude extracts of natural materials or protein hydrolysates can exert DPP-4 inhibition as well. For instance, the DPP-4 inhibition activities of methanol extracts of Ficus benghalensis, Syzigium cumini, Ocimum sanctum, and Eucalyptus sp. have been demonstrated.119,120 The hypoglycemic efficacy of traditional Chinese antidiabetic medicines decoction of Schizandra chinensis Baill., Coptis chinensis, Psidium guajava L., and Morus alba L. has been verified by DPP-4 inhibition by in vivo test. 121 The protein hydrolysates from whey, barbel, and yam can reduce the DPP-4 activity in enzymatic measurements.122–125 Through partition by molecular sieve, the highest inhibition peptide sequences of DPP-4 such as Ala-Pro, Leu-Pro-Val-Pro-Gln, Trp-Ser-Gly, and Phe-Ser-Asp have been found.126–128 Nevertheless, these results from in vitro enzymatic assays cannot guarantee a promising future since physiological regulation of DPP-4 is far more complicated than bench-top experiments. Therefore, the results from direct enzymatic assay for new candidates or hits require further validation such as in vivo investigations to confirm their actual therapeutic values when compared with clinical medicines.
Natural DPP-4 inhibitors from different origins.

FSD, Phe-Ser-Asp; IPM, Ile-Pro-Met; LPVPQ, Lys-Pro-Val-Pro-Gln; WSG, Trp-Ser-Gly.
Caution of using DPP-4 inhibitors
Previous sections have described the biological functions, assay methods, and known natural DPP-4 inhibitors, which are purported to treat DM. However, DPP-4 inhibition can possibly cause unexpected sequela owing to its entanglement with immune response and endothelial functions. In fact, the roles of DPP-4 in tumorigenesis and progression, respectively, have been reviewed recently.144,145 In lung cancer and pancreatic cancer, DPP-4 inhibitor can assuredly reduce cancer progression and promote the overall survival. 146 However, in breast cancer, prostate cancer, and endometrial carcinoma, DPP-4 inhibition would cause the opposite consequence which promotes cancer progression.67,147,148 The role of DPP-4 inhibition in cancer treatment is inconclusive, but it is certain that DPP-4 inhibition in tumorigenesis and tumor development in site-specific tumor should be considered.144,145 In addition to cancer development, opportunistic infection is another issue of DPP-4 inhibition. Anno et al. reported a 69-year-old DM patient, who developed fever after taking vildagliptin for 1 week owing to hypercytokinemia. 149 It is obvious that numerous chemokines such as CXCL3, CXCL4, CXCL 5, and CXCL10 can also function as DPP-4 substrates. 56 Chen et al. have found that DM patients with short-term DPP-4 inhibitor treatment are at higher risk of herpes zoster infection as compared with non-DPP-4 treatment patients after surveying the Longitudinal Health Insurance Database 2000. 150 The development of Hashimoto’s thyroiditis and celiac disease are inversely correlated with DPP4 levels (vide supra), suggesting that DPP-4 inhibition can promote disease progression. Inflammatory bowel disease (IBD), which is a general term for Crohn’s disease and ulcerative colitis, is caused by opportunistic infection or immune cell infiltration.151,152 A meta-analysis published by Radel et al. has indicated that DPP-4 inhibition can increase the risk of Crohn’s disease. 153 In conclusion, DPP-4 inhibitors can be used to treat DM, whereas their complications with other immune disease or cancer should be seriously considered.
Perspectives and future research
The effect of virtual screening via computational biology or informatics further combined with in vitro enzymatic and cell assay, and in vivo animal tests offer a promising approach to discover candidates or hits for expediting the preclinical development process (Figure 3). Nevertheless, poor or ill drug absorption, distribution, metabolism, excretion, and toxicity (ADME/Tox) properties make substantial contributions to drug attritions, 154 and little effort has been dedicated to profiling ADME/Tox properties of DDP-4 inhibitors. As such, it is necessary to predict ADME/Tox parameters in the process of virtual screening, which should be carried by adopting schemes that can consider the unstructured nature of DDP-4, to minimize the late-stage failures.

Effect of natural products in DPP-4 inhibition and the screening methods.
Footnotes
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Conflict of interest statement
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.