
Abstract
Objective
Angiogenesis plays a crucial role in osteoarthritis (OA) by promoting inflammatory cell invasion, supporting neo-innervation and joint tissue fibrosis, and contributing to structural damage and pain. Thrombospondin-3 (THBS-3) is highly expressed in OA cartilage. However, the mechanisms responsible for upregulation of THBS-3 in OA are unclear.
Design
OA chondrocytes and a collagen-induced osteoarthritis (CIOA) mouse model were used as in vitro and in vivo models, respectively. THBS-3 was used to treat chondrocytes in vitro and in vivo. To explore the mechanism of THBS-3 in chondrocytes treatment, we pretreated chondrocytes with a THBS-3 inhibitor and assessed cartilage metabolic function and then analyzed related indicators of vascularization and chondrofibrosis.
Results
Proteomics revealed higher THBS-3 expression in the cartilage of CIOA mice than in that of normal mice. Compared with those from healthy individuals, chondrocytes from OA patients presented significantly increased protein expression of THBS-3. In both in vivo and in vitro experiments, THBS-3 promoted matrix metalloproteinase-13 and disintegrin and metalloprotease with thrombospondin-5, suppressed aggrecan, and promoted the vascularization and chondrofibrosis in dysfunctional chondrocytes from osteoarthritic chondrocytes. THBS-3 activated the transforming growth factor-beta (TGF-β) signaling pathway. The pretreatment of OA chondrocytes with a TGF-β inhibitor before THBS-3 exposure reversed these changes.
Conclusion
THBS-3 promotes the angiogenesis and fibrosis of chondrocytes by activating the TGF-β/Smad2/3 signaling pathway.
Introduction
Osteoarthritis (OA) is a highly prevalent form of arthritis associated with chronic joint pain and physical disability, resulting in a major financial burden and extensive impairments in quality of life. 1 An increasing number of studies have revealed that neo-innervation and joint tissue fibrosis play a role in the development of OA.2,3
The composition of secreted extracellular matrix (ECM) proteins by chondrocytes is dynamically adjusted in response to joint dysfunctional homeostatic environments. Healthy articular hyaline chondrocytes, found in normal cartilage, are known to express high levels of aggrecan and collagen type II. 4 However, in cartilage lesions, the de-differentiation of chondrocytes compromises the quality of hyaline cartilage, resulting in reduced production of essential components and increased expression of fibrotic expression. 5 Fibrotic chondrocytes secrete fibrocartilage-like proteins, resulting in tissue that is stiffer and mechanically inferior to the original hyaline cartilage, while bone morphogenetic protein (BMP-2) acts as a marker of fibrosis and ECM remodeling prior to osteogenesis and ECM calcification. Angiogenesis and fibrosis are closely intertwined, with the blood supply playing a crucial role in tissue fibrosis progression, while fibroblast growth factor-2 (FGF-2), which is involved in fibrosis, also drives the formation of new capillaries.6,7
Thrombospondin-3 (THBS-3) belongs to the thrombospondin protein family, which is a part of the chondrocyte ECM protein family. THBS constitutes a family of five secreted, multimeric, modular matricellular glycoproteins. 8 These proteins exhibit specific spatiotemporal expression patterns in both embryonic and adult tissues and are involved in a wide range of essential processes, including angiogenesis, wound healing, cell proliferation, migration, and the organization of connective tissues. 9 THBS-3 can also regulate postnatal skeletal maturation, influence endochondral ossification within cartilage, and participate in bone formation. 10 However, the exact relationships between THBS-3 and OA are still unclear and further research is needed to explore the role of THBS-3 in OA and its implications for this disease. Therefore, we hypothesized that THBS-3 may play a crucial role in regulating cartilage angiogenesis and fibrosis. To further investigate the hypothesis, we investigated the effects of THBS-3 in human chondrocytes derived from OA samples, C28/I2 immortalized cells lines and a mouse model of collagenase-induced OA (CIOA). The confirmation of this hypothesis will provide a new theoretical basis for the use of THBS-3 in the treatment of OA.
Materials and Methods
Isolation and Culture of Chondrocytes From OA Patients
Articular cartilage was obtained from the knee joints of OA patients (n = 10) who underwent total knee replacement surgery at the First Affiliated Hospital of Harbin Medical University. Cartilage samples were washed three times with phosphate-buffered saline (PBS), cut into 1 mm2 pieces, and then digested in 2 mg/ml collagenase type II (Sigma, St. Louis, MO, USA) at 37°C overnight. Isolated chondrocytes were filtered through a cell strainer and resuspended in fresh medium before centrifugation at 1,000 rpm/min for 5 minutes. Chondrocytes were cultured in DMEM/F-12 (Hyclone; Cytiva) supplemented with 10% fetal bovine serum (FBS) (Hyclone; Cytiva) and 0.5% penicillin‒streptomycin (Beijing Solarbio Science & Technology Co., Ltd.) at 37°C and 5% CO2. Only chondrocytes from the P0-P1 passages were used in subsequent experiments.
The procedures of this study were approved by the Institutional Ethics Committee of the First Affiliated Hospital of Harbin Medical University.
Culture of Immortalized Human Chondrocytes
Immortalized human chondrocytes C28/I2 (Otwo Biotech, Inc., Cat# HTX2308; China) were cultured in Dulbecco’s Modified Eagle Medium/Ham’s F-12 Nutrient Mixture (DMEM/F-12) (Hyclone; Cytiva) supplemented with 10% FBS (Hyclone; Cytiva) and 0.5% penicillin-streptomycin (Beijing Solarbio Science & Technology Co., Ltd.) at 37°C and 5% CO2. Adherent cells were used in subsequent experiments. The culture medium was replaced every 48 hours, and the cells were passaged every 2-3 days.
Culture of Human Umbilical Vein Endothelial Cells
Migration and tube formation experiments were conducted to assess the migratory and tube-forming abilities of human umbilical vein endothelial cells (HUVECs) treated with THBS-3. HUVECs (OTWO, Inc., Cat# HTX1922; China) were cultured in endothelial cell medium (ECM) (ScienCell, Cat# 1001; USA) supplemented with 25 ml of FBS (ScienCell, Cat# 0025; USA), 5 ml of endothelial cell growth supplement (ECGS) (ScienCell, Cat# 1052; USA), and 5 ml of penicillin-streptomycin (ScienCell, Cat# 0503; USA) and used for tube formation and migration experiments at 37°C and 5% CO2. The culture medium was replaced every 48 hours, and the cells were passaged every 2-3 days.
Small Interfering RNA and Transfection Experiments
Small interfering RNAs (SiRNAs) and transfection reagents were purchased from RiboBio Co., Ltd. (China) according to the manufacturer’s instructions. Sequences of THBS-3 siRNA are listed as follows:
S1 sense: 5′-GGACAATTGCCCTAAAGTA-3′;
S2 sense: 5′-GGACCAGTGTGATGACGAT-3′; and
S3 sense GGACTGTATGGAAGTGTAT-3′.
Cell Lysis and Protein Extraction
After three washes with PBS, the cells were subjected to protein extraction. The cells were lysed in radio-immunoprecipitation assay (RIPA) (P0013B, Beyotime) with 1% PMSF (phenylmethylsulfonyl fluoride) (ST506, Beyotime) on ice for 5 minutes and removed with a scraper followed by centrifugation at 12,000 rpm/min for 10 minutes at 4°C. The samples were diluted with SDS-PAGE (sodium dodecyl sulfate polyacrylamide gel electrophoresis) sample loading buffer (Beyotime) and kept at 100°C for 10 minutes.
Protein Assay
The protein concentration was measured at 562 nm by a BCA kit (Boster, China) using microplate reader (Bio-Rad, Richmond, CA, USA).
Western Blotting
Proteins were separated on 10% or 12% SDS‒PAGE mini-gels. Proteins were subsequently transferred onto polyvinylidene difluoride membranes (Thermo Fisher, Hampton, NH) and probed with primary antibodies against THBS-3 (Proteintech, Cat# 19727-1-AP; USA) (1:1,000), aggrecan (Abbkine, Cat# ABP54013; China) (1:1,000), disintegrin and metalloprotease with thrombospondin-5 (ADAMTS-5) (Abcam, Cat# AB41037; USA) (1:1,000), matrix metalloproteinase-13 (MMP-13) (Bioword Technology, Cat# BS1231; China) (1:1000), BMP-2 (Abbkine, Cat# ABP0179; China) (1:1000), FGF-2 (Abbkine, Cat# ABP53280; China) (1:1,000), ANG-2 (Abcam, Cat# AB155106; USA) (1:1,000), VEGF-A (Abcam, Cat# AB214424; USA) (1:1,000), PDGF-B (Abcam, Cat# AB178409; USA) (1:1,000), transforming growth factor-beta 1 (TGF-β1) (Abcam, Cat# AB179695; USA) (1:1,000), p-Smad2/3 (Abbkine, Cat# ABP0048; China) (1:1,000) and GAPDH (ABclonal, Cat# A19056; China) (1:10,000) overnight at 4°C. The blots were then incubated with a secondary antibody (LI-COR, Cat# 926-32211; USA) (1:10,000) at room temperature for 1 hour. Visualization was achieved via an LI-COR imaging system (Biosciences, USA), and subsequent quantification was carried out with Image-Pro Plus software.
Quantitative Real-Time Polymerase Chain Reaction
Total RNA from chondrocytes was extracted with TRIzol reagent (Invitrogen, Cat# R1100; USA) according to the manufacturer’s instructions. Total RNA concentration was determined by measuring the absorbance at 260/280 nm using a spectrophotometer (BioTeke Corporation). cDNA was then synthesized from 1 µg of total RNA via the 4*DN Master Mix with gDNA Remover and 5*RT Master Mix II (TOYOBO, Cat# FSQ-301; Japan), and 25 ng of the resulting cDNA was used for quantitative real-time polymerase chain reaction (qRT-PCR). The qRT-PCR mixture contained 5 μl of iTaq Universal (Bio-Rad, Cat# 1725124; USA), 0.5 μl of each primer, and 1 μl of cDNA and nuclease-free H2O in a total volume of 10 μl. The samples were amplified for 40 cycles via the Applied Biosystems® 7500 Real-Time PCR System (Thermo Fisher Scientific). The 2^-ΔΔCT method was employed for calculating relative gene expression levels, with normalization against the housekeeping gene GAPDH. Sequences of THBS-3 and GAPDH are listed as follows:
THBS-3 sense: 5′-GTGTCCTCTTTGGCCTCTATTC-3′; antisense: 5′- CTGGTATCGCACCAGTACTTT-3′.
GAPDH sense: 5′-GGTGTGAACCATGAGAAGTATGA-3′; antisense: 5′-GAGTCCTTCCACGATACCAAAG-3′.
Tube Formation Assay
Endothelial cells were seeded at a density of 2 × 104 cells per well in μ-Slide Angiogenesis (ibidi, Cat# 81506; GER) precoated with 50 μl of Matrigel in each well (Corning, Cat# 354230; USA). The endothelial cells in the plates were subsequently incubated at 37°C with 50, 100, or 200 nM recombinant human THBS-3. After 6 hours of incubation, the HUVECs were imaged using an inverted microscope (Leica) and analyzed via Image-Pro Plus software.
Cell Migration Assay
A wound scratch assay was used to assess the migration of HUVECs treated with 50, 100, or 200 nM human recombinant THBS-3 protein. In brief, 5.0 × 105 cells were seeded per well in a six-well culture plate and incubated for 24 hours at 37°C until they reached confluence. A scratch was subsequently created via a sterile p200 pipette tip, and the resulting cell debris was washed three times with PBS. Next, the cells were incubated in basic ECM supplemented with various concentrations of human recombinant THBS-3 proteins. Images of the wounds were captured immediately after scratching and 24 hours later. ImageJ software was used to measure the area change in the scratched areas. The migration rate was calculated as follows: migration area (%) = (A0–An)/A0 × 100, where An and A0 represent the remaining area and initial area of the wound, respectively.
Mouse Model of Type VII CIOA
For the CIOA mouse model, 12-week-old male C57BL/6 (Animal Center of Harbin Medical University) mice were anesthetized, and then, type VII collagenase was injected into the left knee. 2’OMe+5’Chol-modified THBS-3 siRNAs labeled with 5FAM were purchased from Ribo Company (RiboBio, China). The mice (n = 18) were randomly divided into the following groups: the control (n = 6), CIOA (n = 6), and CIOA+siTHBS-3 (n = 6) groups. At 7 days post-injection, 5 nM siTHBS-3 (OA + siTHBS-3 group) was injected intra-articularly once a week into the left knee joints of the mice. The mice in the control group were not injected. The mice were sacrificed at week 7 for bone and cartilage analysis. All the animals were kept in a controlled environment that ensured that they were free from specific pathogens. The animals were subjected to a 12-hour light and 12-hour dark cycle and were provided with a standard chow diet. All in vivo experiments were approved by the Harbin Medical University Committee on the Use and Care of Animals (No. 2023JS57) in accordance with the ARRIVE guidelines 2.0 https://arriveguidelines.org
Micro–Computed Tomography
Subchondral bone explants from the mouse joint specimens were initially fixed overnight in a 10% formalin solution. These explants were subsequently examined using micro–computed tomography (micro-CT) imaging (Quantum GX, Perkin Elmer, Waltham, USA).
Histology
Knee joints were fixed in 4% paraformaldehyde for 24 hours and then decalcified over an 8-week period using a 10% EDTA decalcifying solution at pH 7.4. The samples were embedded in optimal cutting temperature (OCT) compound (Sakura Finetek) or paraffin. The medial portion of each sample was longitudinally oriented and sectioned into 4 μm slices. The medial compartment of the knee joint was subjected to staining procedures, including haematoxylin-eosin (H&E) staining, Safranin O and Fast Green staining and immunohistochemistry. Images were captured via a light microscope (BX53; Olympus Corporation). Tissue histological scoring was performed according to the Osteoarthritis Research Society International (OARSI) grading system. 11 Pathological grading was defined by the depth of OA progression into the cartilage layer, whereas staging was defined by the extent of cartilage involvement on one side of the joint cavity. The scoring was an amalgamation of OA grading and staging, calculated as the product of grading and staging. Histological scoring was conducted by two independent observers in a blinded manner.
Immunohistochemistry
Immunohistochemical analysis was performed using the PK-4001 histochemistry kit (Vector), with endogenous peroxidase activity quenched by 3% H2O2, antigen retrieval achieved through 0.1% trypsin, and blocking carried out using normal goat serum for 20 minutes. Knee joints sections were incubated overnight at 4°C with primary antibodies targeting the following proteins: aggrecan (1:100; Abcam), MMP-13 (1:100; Abcam), ADAMTS-5 (1:100; Abcam), ANG-2 (1:100; Abcam), BMP-2 (1:100; Abcam), and FGF-2 (1:100; Abcam). After incubation, sections were rinsed with PBS and incubated with the appropriate biotinylated secondary antibodies, followed by horseradish peroxidase-conjugated streptavidin-biotin. Immunoreactions were developed using the DAB (3,3'-diaminobenzidine) substrate chromogen system (Dako), and nuclei were counterstained with hematoxylin, differentiated with saturated lithium carbonate to yield a blue color. The number of positive cells in the central areas of the articular cartilage was quantified using ImageJ software (Media Cybernetics).
Proteomics
Mouse knee joint cartilage was dissected, snap-frozen in liquid nitrogen, and stored at −80°C. Tissues were lysed in 50 mM ammonium bicarbonate with protease/phosphatase inhibitors, denatured at 95°C, and digested with trypsin (1:50, w/w) at 37°C overnight. Peptides were desalted on C18 columns, dried, and reconstituted in 0.1% formic acid for analysis. LC-MS/MS was performed on a Q Exactive HF-X instrument coupled to an EASY-nLC 1200; peptides were separated on a C18 analytical column using a 150-minute gradient and analyzed in data-dependent acquisition mode. Raw data were processed with Firmiana/Mascot against the Mus musculus reference proteome; identifications were filtered to a < 1% FDR (false discovery rate) at peptide-to-spectrum match (PSM) and protein levels. Label-free quantification was performed using normalized spectral intensities. Differentially expressed proteins were identified using combined fold change and statistical testing.
Statistical Analysis
The data are presented as the mean ± standard error of the mean (SEM). For comparisons between two groups, a two-tailed Student’s t test was employed. For multiple group comparisons, one-way analysis of variance (ANOVA) was used. Initially, the homogeneity of variance was assessed, followed by post hoc multiple comparisons to evaluate differences between groups. Statistical significance was defined as P < 0.05.
A schematic depicting the experimental design and downstream assays from in vitro and in vivo models (
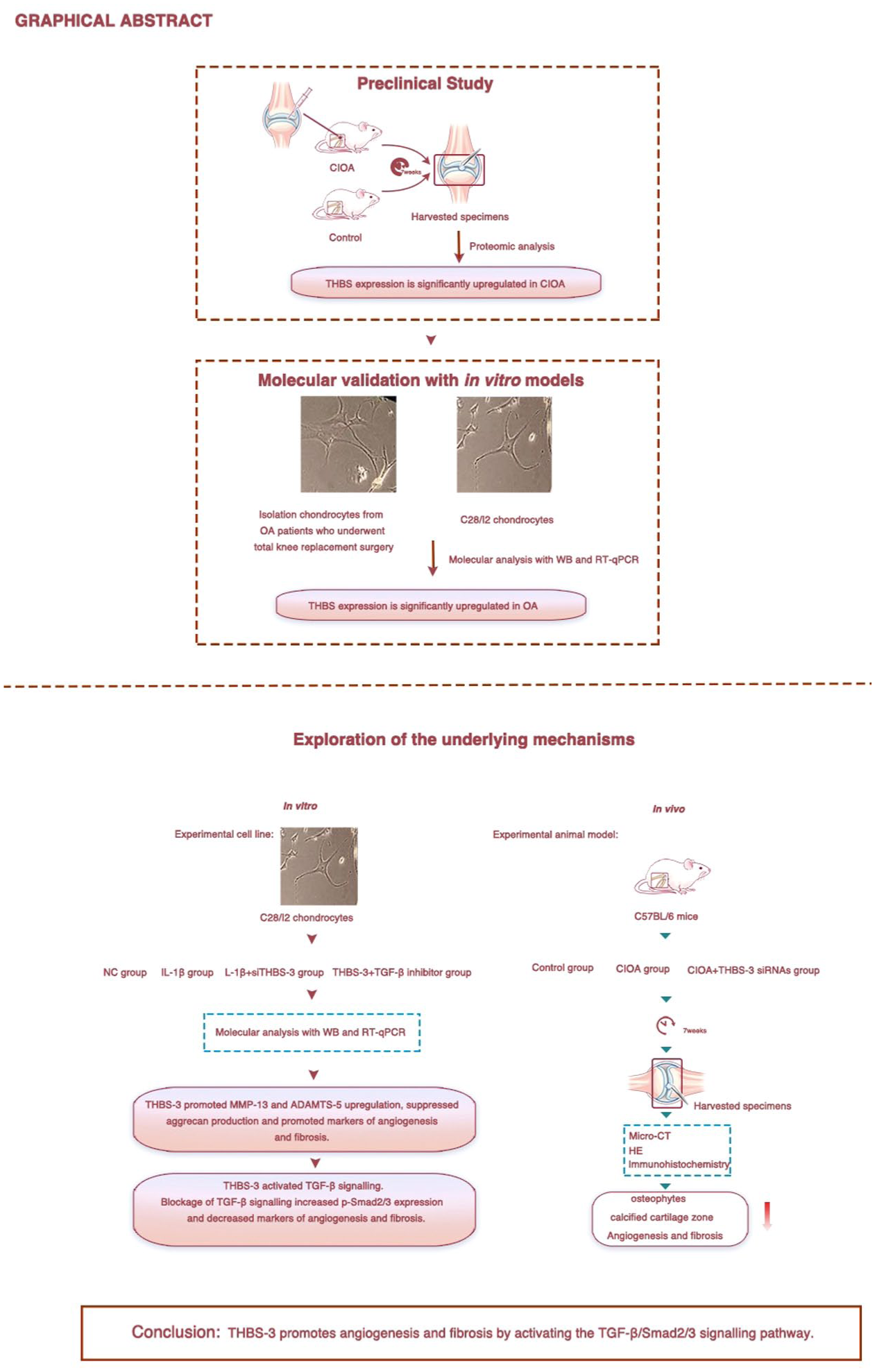
A schematic depicting the experimental design and downstream assays from in vitro and in vivo models.
Results
THBS-3 Expression Is Elevated in OA Cartilage, Reducing Expression of Aggrecan, and Increasing Expression of MMP-13 and ADAMTS-5
Mouse studies: Proteomic analysis revealed a significant difference in THBS-3 levels in CIOA and control cartilage samples. Compared with the control group, the CIOA group presented upregulation of 234 proteins and downregulation of 96 proteins. A volcano plot was created to illustrate the overall distribution of differentially expressed proteins between the control and CIOA groups. The ratio of THBS-3 expression in the CIOA group to that in the control group was 1.614, indicating significant upregulation (
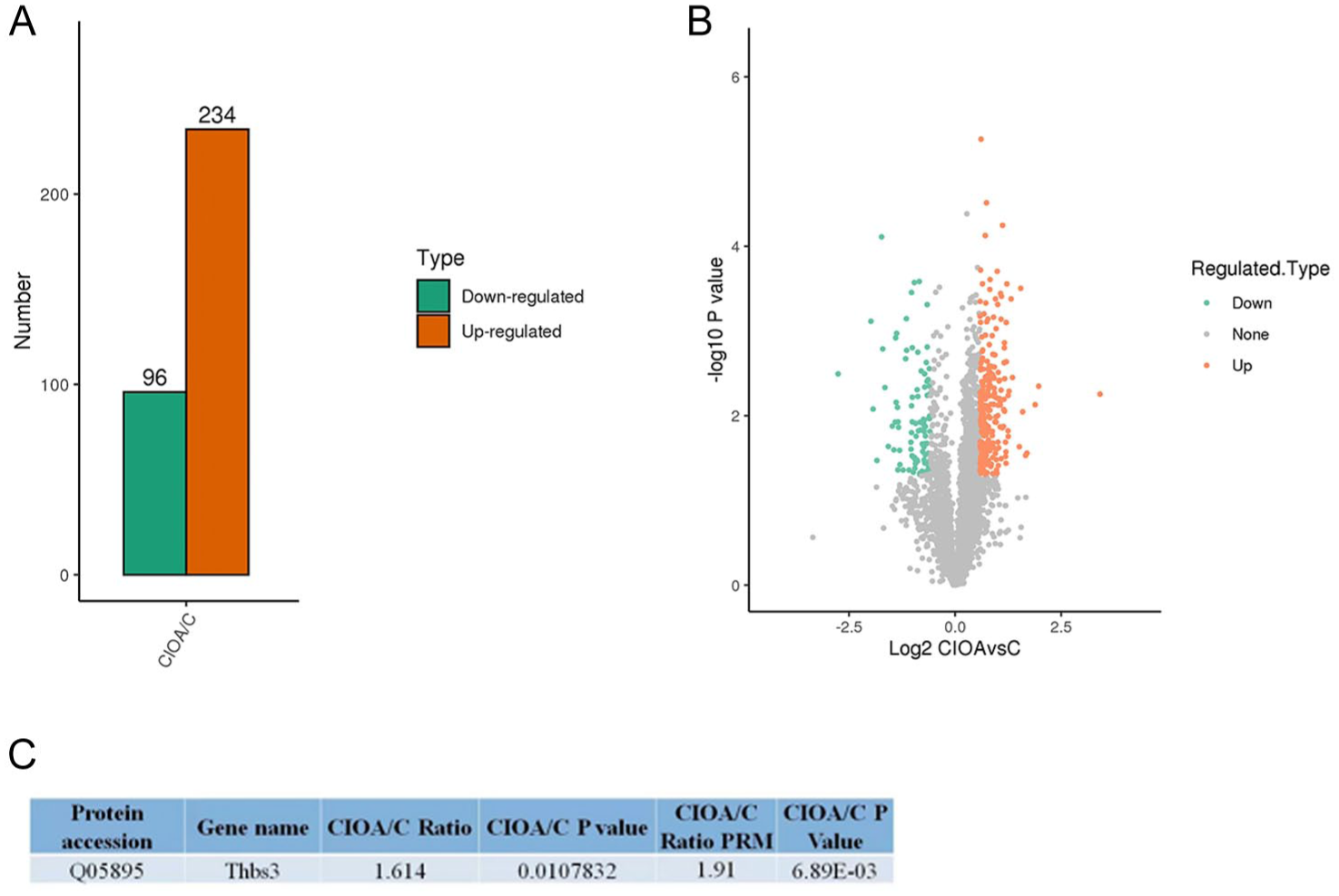
Proteomic analysis of knee joint tissues in normal mice and collagen-induced osteoarthritis (CIOA) model mice. (
Human studies: To investigate the involvement of THBS-3 in human OA, we analyzed THBS-3 expression in human cartilage samples. In OA samples (n = 10) the expression level of THBS-3 was significantly increased (P = 0.0236) (
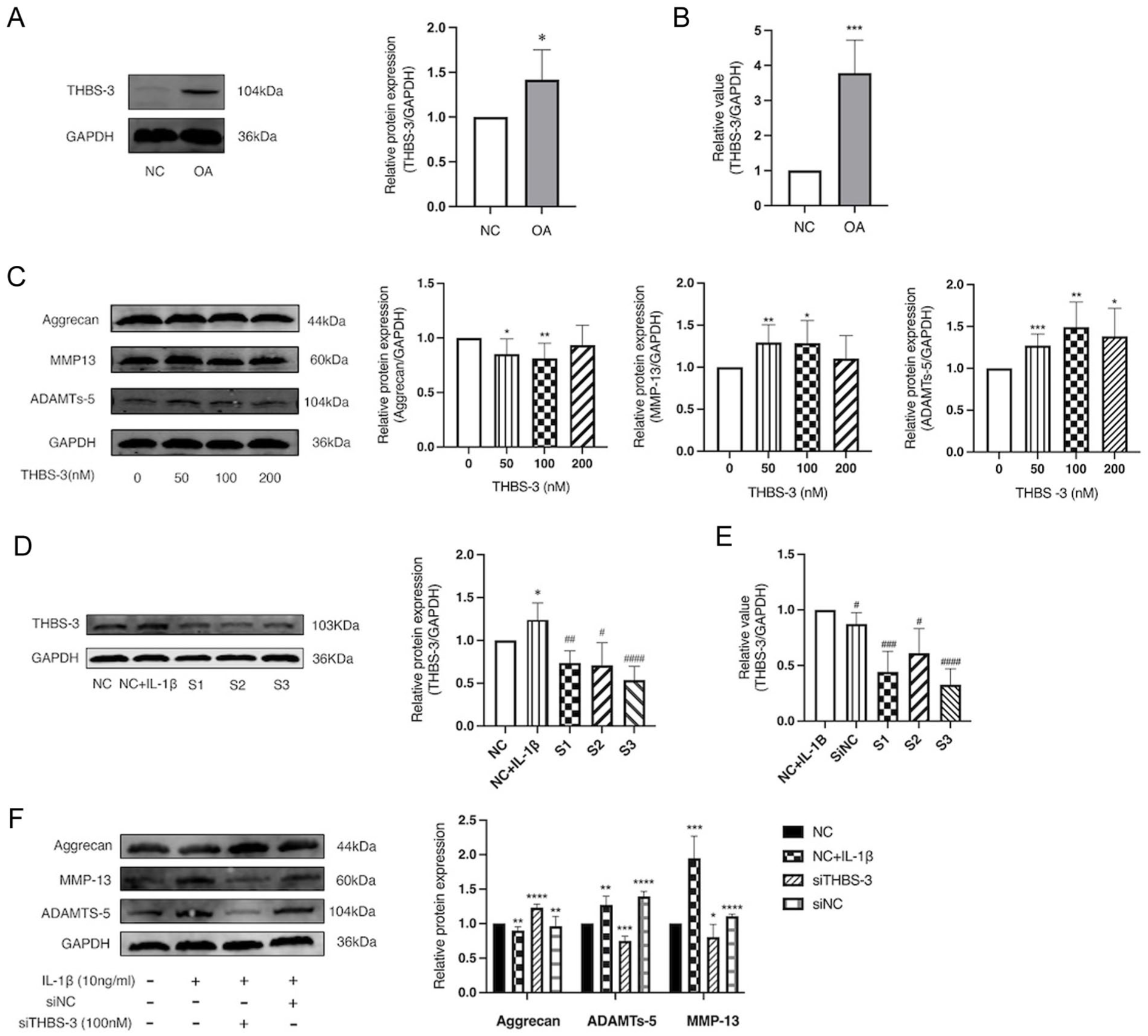
THBS-3 expression is elevated in OA cartilage, reducing expression of aggrecan and increasing expression of MMP-13 and ADAMTS-5. (
To further explore the influence of THBS-3 on catabolic and anabolic activity in chondrocytes, we exposed C28/I2 chondrocytes to 50, 100, or 200 nM human recombinant THBS-3 protein for a 24-hour period. We assessed the expression of MMP-13, ADAMTS-5 and aggrecan. Our findings indicated that THBS-3 actively promoted the expression of MMP-13 and ADAMTS-5 while simultaneously suppressing the expression of aggrecan (
In many published in vitro studies, stimulation of chondrocytes with IL-1β is used as a cellular model of OA. 3 To investigate whether IL-1β affects THBS-3 expression, we examined the levels of THBS-3 expression in C28/I2 chondrocytes after 24 hours of IL-1β (10 ng/ml) treatment.
For siRNA studies, we designed three THBS-3 siRNAs (S1-S3) and selected the most effective siRNA through WB and RT-qPCR analysis for subsequent experiments. The results indicated a significant increase in THBS-3 expression in C28/I2 chondrocytes in response to IL-1β (P = 0.0138). In both the WB (P < 0.0001) and RT-PCR (P < 0.0001) experiments, the expression of THBS-3 was lowest in the S3 group (
For a more in-depth examination of the influence of THBS-3 on IL-1β-induced ECM degradation, C28/I2 chondrocytes were subjected to transfection with THBS-3 siRNA or a scrambled siRNA control. THBS-3 siRNA resulted in a substantial reduction in the IL-1β-induced upregulation of MMP-13 and ADAMTS-5 in chondrocytes. In contrast, THBS-3 siRNA led to increased expression of aggrecan compared with that in the normal chondrocytes treated with IL-1β alone (
THBS-3 Increases Expression of Markers of Vascularization and Fibrosis in Chondrocytes C28/I2
To further investigate the role of THBS-3 in chondrocytes, we stimulated C28/I2 chondrocytes with 50, 100, or 200 nM recombinant human THBS-3 protein and conducted WB analysis to examine markers associated with angiogenesis (ANG-2, VEGF-A, and PDGF-B) and fibrosis (BMP-2 and FGF-2). THBS-3 promoted ANG-2, VEGF-A, PDGF-B, BMP-2, and FGF-2 protein expression (
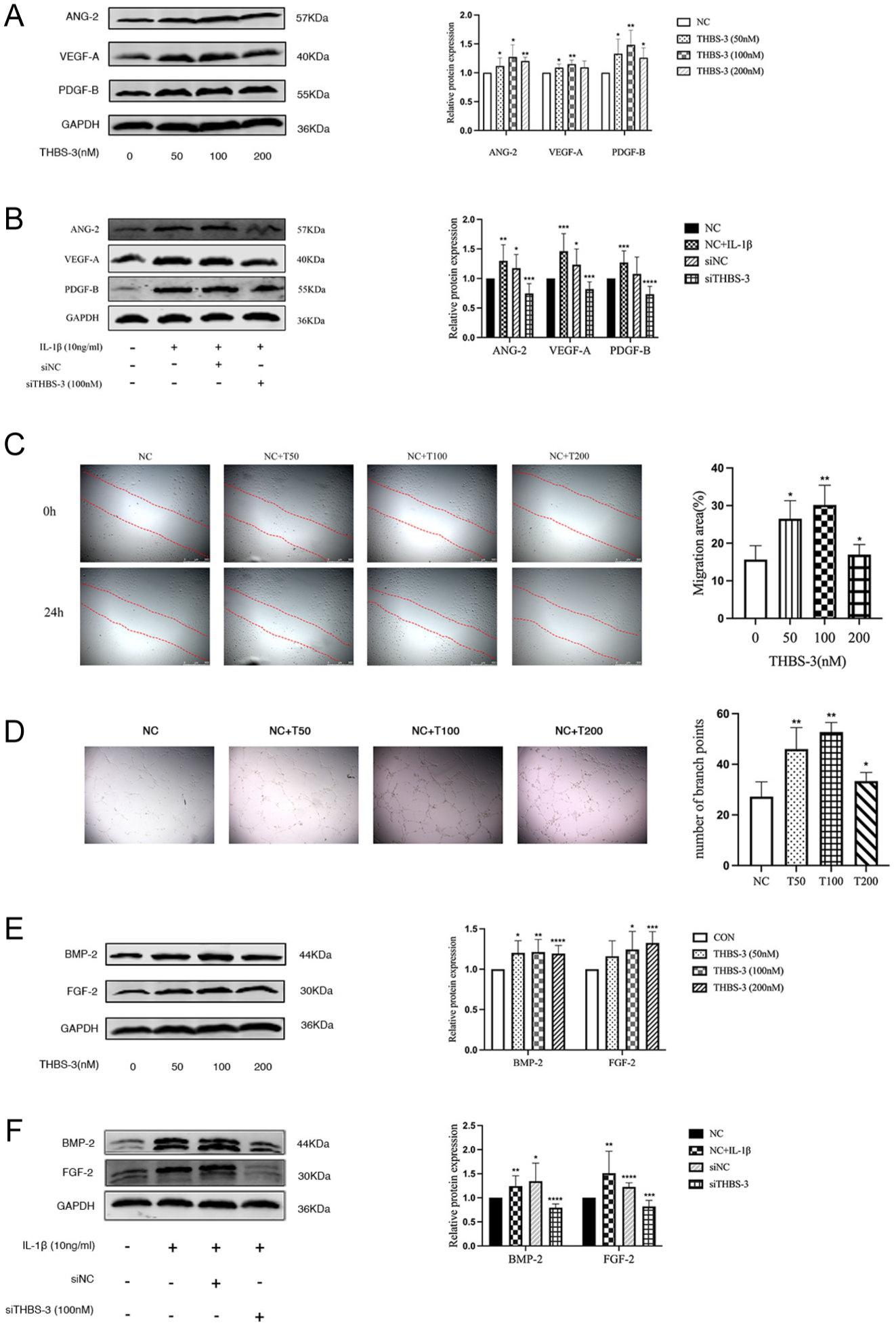
THBS-3 increases expression of markers of vascularization and fibrosis in chondrocytes C28/I2. (
THBS-3 Promotes Migration of HUVECs
The migration of endothelial cells is a pivotal process in angiogenesis and facilitates the extension of blood vessels. We prepared HUVECs that were treated with different concentrations of THBS-3 (0, 50, 100, or 200 nM), and we assessed the impact on angiogenesis through analysis of endothelial cell migration and tube formation. THBS-3 significantly increased the migration of HUVECs compared with that of the control group. With increasing THBS-3 concentration, the migration area expanded proportionally, reaching its most pronounced extent at a concentration of 100 nM (P = 0.0040) (
THBS-3 siRNA Application Effectively Inhibits Bone Spur Formation and Mitigates Pathological Changes in the CIOA Model
The in vivo data described justified further investigation of the therapeutic efficacy of THBS-3 inhibition using siRNA in the CIOA model. Intra-articular injections of THBS-3 siRNA (5 nM) were administered once a week for 7 weeks. H&E staining revealed that the tidemark, which had moved upward in CIOA mice, was normalized by THBS-3 siRNA treatment (

THBS-3 siRNA application effectively inhibits bone spur formation and mitigates pathological changes in the cartilage tissue of the mice with CIOA. (
THBS-3 Promotes Expression of Markers of Vascularization and Fibrosis in C28/I2 Chondrocytes Through the TGF-β/Smad2/3 Signaling Pathway
TGF-β1 is a crucial growth factor for developing, maintaining, and repairing articular cartilage.
12
To investigate whether THBS-3 plays a role in OA through the TGF-β /Smad2/3 pathway, we made an intriguing discovery. These results showed that THBS-3 could activate TGF-β pathway, which peaked at 6 hours (P = 0.0353) (
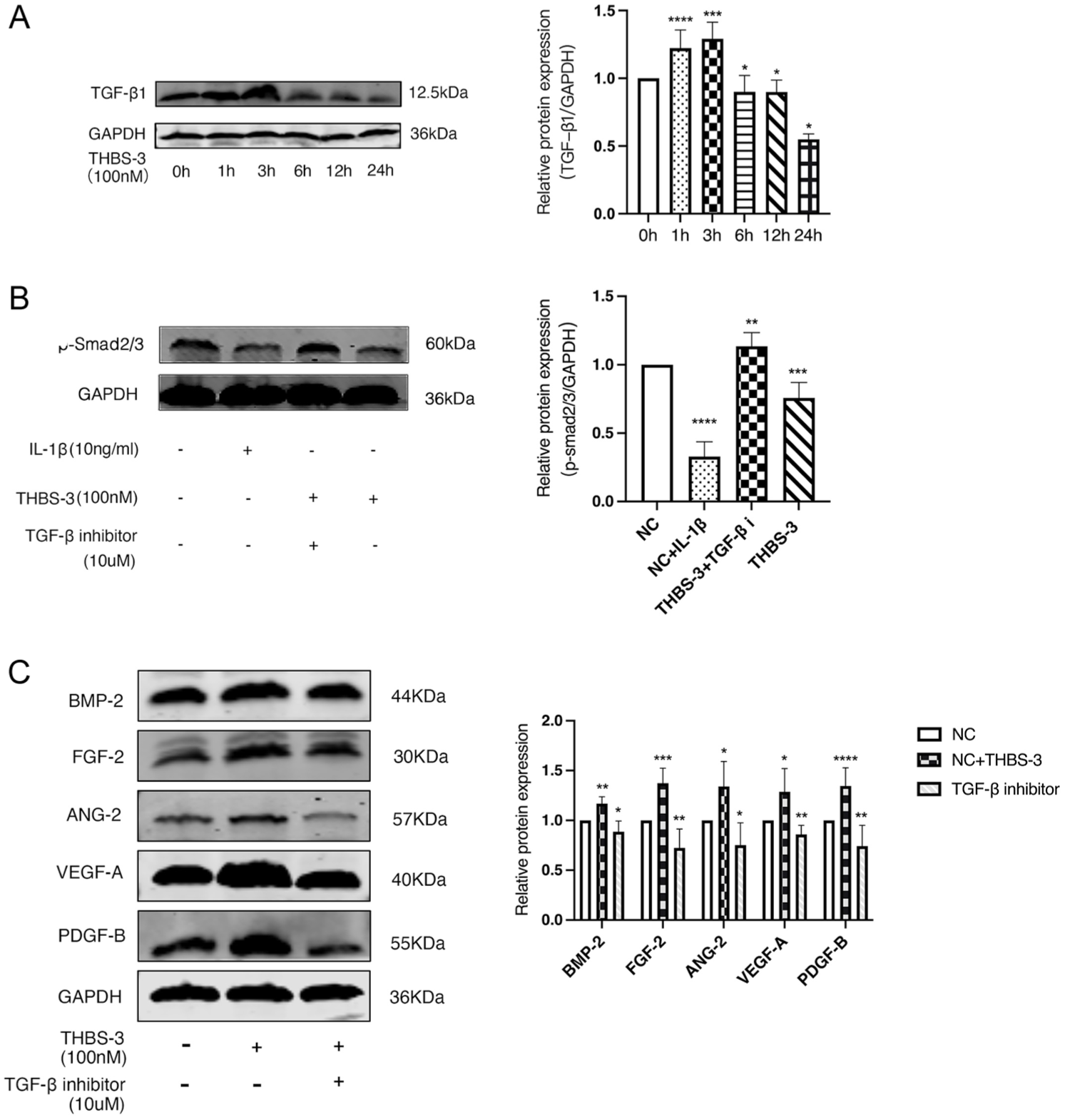
THBS-3 promotes expression of markers of vascularization and fibrosis in C28/I2 chondrocytes through the TGF-β/Smad2/3 signaling pathway. (
Discussion
The aim of this study was to explore the mechanistic role of THBS-3 in OA and the results can be summarized as follows:
First, we confirmed that OA resulted in upregulated expression of THBS-3. THBS-3 actively promoted catabolism while simultaneously suppressing anabolic activity. Second, THBS-3 promotes expression of angiogenic and fibrotic markers in chondrocytes, which may drive these processes in OA cartilage. Third, we demonstrated that THBS-3 in chondrocytes promotes angiogenic and chondrofibrotic markers in vitro by activating the TGF-β/Smad2/3 signaling pathway in OA.
A published meta-analysis of gene expression in OA and non-OA chondrocytes has revealed a significant increase in the expression of THBS-3 and COMP in OA. 13 Our findings support the previously published observations in gene expression and further highlight a notable increase in the expression of THBS-3 in OA. We used the C28/I2 cell line for all the molecular investigations because of the variability and heterogeneity of the primary chondrocytes isolated from human clinical specimens. A published in vivo study has shown that mice lacking the ECM proteins THBS-1, THBS-3, THBS-5, and type IX collagen exhibit skeletal abnormalities. 14 THBS-3, THBS-5, and type IX collagen all play direct roles in regulating linear growth in growth plate tissue. Another in vivo study has revealed that THBS-3 regulates postnatal bone maturation, influencing endochondral ossification. In cases of THBS-3 deficiency, accelerated ossification of the calcified cartilage in the femoral head occurs. 10 However, interestingly, the effects of THBS-3 on various cells and tissues are not the same. Therefore, further research was needed to explore the role of THBS-3 in OA, prompting us to perform these studies.
An imbalance between anabolic and catabolic processes in chondrocytes is believed to be an important driver for the development of OA. We observed that THBS-3 inhibits biosynthetic activities in chondrocytes. Furthermore, research indicates that angiogenesis and fibrosis are involved in the progression of OA. Studies have found that PDGFR-β in endothelial cells promotes angiogenesis through the formation of the PDGFR-β/talin1/FAK complex. In PDGFR-β knockout mouse OA models, H-type blood vessels were reduced, subchondral bone degeneration is improved, and cartilage degeneration is alleviated. 15 Other research has shown that in OA mouse model cartilage tissue, the number of CD31-positive newly formed blood vessels increased with the duration of modeling, and the expression of VEGF-A, VEGFR-2, COX-2, and iNOS was correlated with the severity of OA. 16 Angiogenesis and fibrosis are closely interconnected, with blood supply playing a crucial role in the process of inflammation and fibrosis. Various studies have confirmed the presence of fibrosis in the OA infrapatellar fat pad (IFP). 17 The IFP forms numerous blood vessels and fibrous tissues, including the synovial membrane. 18 It has been reported that blood vessel formation in the IFP accelerates the development of fibrosis in the knee joint following injury. 19 Accumulating evidence demonstrates that angiogenesis in OA is a multifocal event, taking place across distinct joint tissues, such as the articular cartilage, subchondral bone, and IFP, thereby underscoring its central role in OA pathophysiology. Nevertheless, whether THBS-3 drives the pathogenesis of OA via this mechanism has yet to be elucidated. In a study exploring the influence of THBS-3 on the biological behavior and survival of osteosarcoma patients, the THBS-3 gene exhibited significant differential expression in osteosarcoma. Increased levels of THBS-3, which functions as a potent stimulant for tumor progression, were found to actively stimulate angiogenesis. 20 . BMP-2 is a marker of fibrosis and ECM remodeling before osteogenesis and ECM calcification occur. Numerous studies provide evidence for FGF-2 as a promoter of fibrosis. The role of FGF-2 as a pro-angiogenic protein is also well established. 21 VEGF, ANG-2, PDGF, and FGF-2 promote angiogenesis by interacting with various receptors on the surface of endothelial cells. In addition, FGF-2 regulates the expression of cadherins, integrins, and several ECM components, thereby facilitating the maturation of newly formed blood vessels and modulating endothelial cell adhesion to the ECM. 22 Preliminary results obtained by our research group showed that the expression level of THBS-3 was significantly greater in OA cartilage cells than in normal cells. We applied immunohistochemical staining to detect the expression levels of the joint cartilage anabolism marker aggrecan and joint cartilage catabolism markers MMP-13 and ADAMTS-5. Exogenous THBS-3 has been shown to promote cartilage collagen catabolism and inhibit synthetic metabolism in CIOA model mice. In this study, we conducted a series of experiments using exogenous THBS-3 to treat normal chondrocytes. The discovery that exogenous THBS-3 can decrease the protein expression level of aggrecan and increase the protein expression levels of MMP-13 and ADAMTS-5 in OA chondrocytes further confirms the destructive effect of THBS-3 against OA and that THBS-3 can significantly alleviate pathological changes in OA model mice, confirming the therapeutic effect of siTHBS-3 application on OA in vivo and in vitro. To visually evaluate the expression of markers of vascularization and fibrosis in chondrocytes, we used WB and immunohistochemical analysis to detect ANG-2, VEGF-A, PDGF-B, BMP-2, and FGF-2. Exogenous siTHBS-3 application can significantly ameliorate the expression of vascularization and fibrosis in OA chondrocytes.
TGF-β1 has been extensively investigated in OA in the context of its crucial role the development, maintenance, and repair of articular cartilage. 12 TGF-β1 is also a key modulator of fibrosis since it induces fibrogenesis through the modulation of Smad molecules and other Smad independent pathways. Previous studies have shown that in in vitro culture of endothelial cells and in vivo vascular generation model experiments, THBS-4 promotes angiogenesis through TGF-β1 regulation. 23 Studies have reported elevated expression of THBS-1 in synovial tissues of individuals with rheumatoid arthritis, and TGF-β1 significantly increased the expression of THBS-1 at both the mRNA and protein levels in the synovium. 24 THBS-1 may be involved in the pathogenesis of rheumatoid arthritis through the TGF-β1/THBS-1 pathway. THBS-1 inhibits the osteogenic differentiation of human mesenchymal stem cells by activating TGF-β. 25 Furthermore, our findings indicate that the expression of TGF-β1 in chondrocytes stimulated with THBS-3 (100 nM) progressively increases over time, with peak expression observed at 6 hours. These observations reinforce the notion that properly controlled endogenous production of TGF-β1 is essential for maintaining chondrocyte homeostasis, whereas excessive or inadequate activation of TGF-β1 can be detrimental to joint health. 26 The elevated TGF-β1 levels prompted us to further explore the regulatory mechanisms involved. Given the observed alterations in the expression levels of TGF-β signaling receptors in the context of OA, it is reasonable to expect changes in the activity of their signal mediators, Smad2/3.
Research has provided compelling evidence of reduced Smad2 phosphorylation levels in cartilage during OA progression, which is evident in both spontaneous (STR/Ort) and mouse models of CIOA.
27
Furthermore, Smad2 phosphorylation was reportedly lower in the cartilage of older mice than in that of their younger counterparts.
28
While Smad3 phosphorylation has not been specifically examined in these models, a recent study reported decreased Smad3 phosphorylation levels in Smurf-2 transgenic mice that spontaneously develop an OA-like phenotype.
29
These collective findings strongly suggest that OA is associated with a compromised TGF-β/Smad2/3 signaling pathway. Our findings are consistent with those of previous studies. In the OA model, a reduction in p-Smad2/3 expression was observed. For more detailed analysis of the pathways associated with THBS-3 in chondrocytes were stimulated with THBS-3, which inhibited p-Smad2/3 expression (
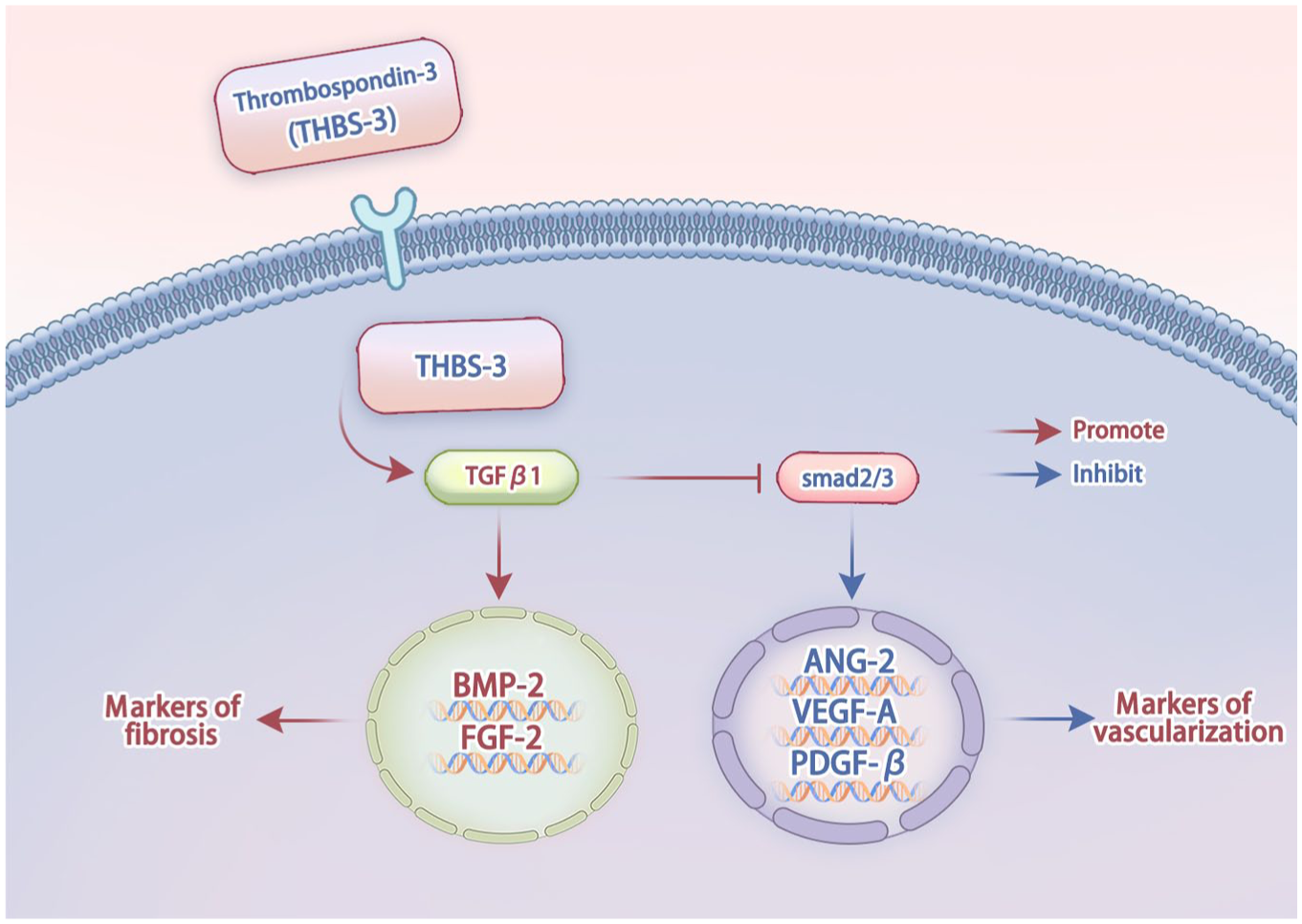
Schematic diagram of the mechanism of THBS-3 on promotes cartilage angiogenesis and fibrosis.
This study provides the first direct evidence that THBS-3 acts as a pivotal upstream regulator of cartilage angiogenesis and fibrosis through modulation of the TGF-β/Smad2/3 pathway in OA. Our findings elucidate the link between THBS-3 and pathological remodeling at both molecular and tissue levels, offering novel insights into OA pathogenesis. Furthermore, these results identify THBS-3 as a promising therapeutic target, laying a foundation for the development of precise, mechanism-based interventions in OA management.
This study has certain limitations. Owing to the formidable challenges associated with procuring cartilage tissue from healthy control donors, our only viable option was to acquire chondrocytes from normal individuals through commercial channels. In addition, the stability of exogenously administered siTHBS-3 in the animal model is limited; future studies could explore the use of advanced biomaterials to enhance in vivo stability and sustained efficacy, which is crucial for developing targeted OA therapies.
Conclusion
The results presented in this study lead to the conclusion that THBS-3 plays an important role in the regulation of angiogenesis and fibrosis via the TGF-β/Smad2/3 pathway in OA. Furthermore, we propose THBS-3 as a novel therapeutic target for treating OA.
Footnotes
List of abbreviations
Angiotensin-2 ANG-2
Bone morphogenetic protein BMP-2
Disintegrin and metalloprotease with thrombospondin ADAMTS-5
Extracellular matrix ECM
Fibroblast growth factor FGF-2
Haematoxylin-eosin H&E
Human umbilical vein endothelial cells HUVECs
Matrix metalloproteinase MMP-13
Microcomputed tomography Micro-CT
Microcomputed tomography Micro-CT
Mouse model of type VII collagenase-induced OA CIOAO
Osteoarthritis Research Society International OARSI
Platelet-derived growth factor-B PDGF-B
Small interfering RNA siRNA
Thrombospondin-3 THBS-3
Transforming growth factor-beta TGF-β
Vascular endothelial growth factor-A VEGF-A
Ethical Approval
The procedures of this study were approved by the Institutional Ethics Committee of the First Affiliated Hospital of Harbin Medical University (No. 2023JS57 Date: December 28, 2023).
Author Contributions
JY.Y. and SY.W. contributed to the conception and design of the study. HY.L., XY.Z., and YL.W. performed the experiments and contributed to the analysis and interpretation of data. YP. Z, A.M., and ZY.Z. contributed to reviewing the manuscript. All approved the submitted manuscript.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants from the National Natural Science Foundation of China to Zhiyi Zhang (NSFC 82271826) and Shuya Wang (NSFC 82202020), and partially by the First Affiliated Hospital of HMU Merit Review Frontiers grant to Shuya Wang (HYD2020YQ0008).
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
All data generated or analyzed during this study are included in this published article.