
Abstract
The larynx sometimes requires repair and reconstruction due to cancer resection, trauma, stenosis, or developmental disruptions. Bioengineering has provided some scaffolding materials and initial attempts at tissue engineering, especially of the trachea, have been made. The critical issues of providing protection, maintaining a patent airway, and controlling swallowing and phonation, require that the regenerated laryngotracheal cartilages must have mechanical and material properties that closely mimic native tissue. These properties are determined by the cellular and proteomic characteristics of these tissues. However, little is known of these properties for these specific cartilages. This review considers what is known and what issues need to be addressed.
Keywords
Introduction
The larynx represents arguably the most intricate cartilaginous complex in the human body, in which a variety of extrinsic and intrinsic muscles interact with cartilage to control aspects of breathing, swallowing, coughing, and phonation. Cartilage is a critical source of rigidity that permits the maintenance of anatomical form and function; ensuring that tubular structures remain patent and providing sites for muscle attachment. If the larynx is impaired by congenital defects, cancer or trauma, reconstruction of various structures within the larynx, including the cartilage, could improve functionality and quality of life for the patient. Airway reconstruction is at the vanguard of clinically translatable tissue engineering; however, the majority of investigation to date has focused on the mechanical properties and replacement of the trachea, potentially with the inclusion of the cricoid.1,2 Primary laryngeal reconstruction could range from a single cartilage, such as the thyroid cartilage, to the entire larynx; thus knowledge about each laryngeal cartilaginous component is required to enable the development of suitable scaffolding materials to engineer these cartilages. There have been some initial attempts to tissue engineer the hemilarynx, using either decellularized matrix 3 or synthetic materials1,2,4-8 but there have been issues with incomplete cellularization and inadequate mechanical properties. Because of the roles of these cartilages in keeping the airway patent, it is critical that the cartilaginous structures, especially the cricoid and thyroid cartilages, are functional immediately upon placement into the patient.
In order to commence tissue engineering of the laryngeal cartilage, it is necessary to know not just the specific shapes of these components, but their biochemical and material properties.4,9-14 Furthermore, many of the properties of cartilage are determined by their cellular composition and density, and the proteomic/glycomic composition of the extracellular matrix (ECM).15,16 In this review, the literature of the mechanical, cellular, and proteomic properties of the laryngotracheal cartilages will be examined in order to provide a baseline of knowledge and illustrate gaps in the literature. As outlined, regenerative medicine of the airway has been trachea focused, and the mechanical and protein compositions of tracheal cartilage have been investigated. The foundations of cartilage proteomics will be described and the information elicited from investigation of cartilage from extra-airway sources will be discussed to provide insight for future laryngological research. The current status of laryngotracheal tissue engineering clinical translation is outside of the scope of this review. Recently published, comprehensive reviews on these topics are suggested to readers.1,2,6
Gross Structure and Biomechanics of Laryngotracheal Cartilages
Within the larynx, there are 3 unpaired cartilages: the thyroid cartilage, the cricoid cartilage, and the epiglottis; and 3 paired cartilages: the arytenoid, corniculate, and cuneiform cartilages. The hyoid is the sole bony structure local to the larynx, placed superiorly, providing insertion sites for several muscles that effect laryngeal movement. Although the cartilaginous tracheal rings are not part of the larynx, reconstructive surgery often includes one or more tracheal rings.
Thyroid Cartilage
The thyroid cartilage functions to protect the airway and provide support to the anterior end of the vocal folds, associated muscles, and membranes. The laminae are fused at the anterior midline, with gender differences in the size, prominence, and angle between the laminae.17,18 Each lamina has a superior and inferior horn (cornu). On the posterior aspect, the thyroarytenoid, thyroepiglottic, and vocalis muscles have insertions. The collagen fibrils (primarily collagens I, II, and III) of the vocal ligament tendon insert directly into the thyroid cartilage proper as there is no perichondrium in this area. 19 There are also extrinsic muscles that attach to the abluminal wall of the thyroid cartilage. The cricothyroid muscle originates on the cricoid cartilage and inserts on the inferior border of the thyroid cartilage to move the thyroid cartilage forward. Key muscles imparting tensile stress on the larynx are the thyrohyoid, sternothyroid, and the thyropharyngeal part of the inferior constrictor muscle of the pharynx. 17 These muscles are primarily for swallowing and speech functions.
The thyroid cartilage is composed of hyaline cartilage.20,21 Distinct gender differences in ossification of the thyroid cartilage are seen. Males have an earlier age to initiate ossification and a more extensive area of ossification.20,22 These tissue level differences may be due to decreases in the levels of chondroitin-4- and -6-sulphates in a rim of interterritorial matrix around chondrocytes in males.22,23 The anterior half of the female thyroid cartilage does not ossify, and does not demonstrate these changes in the levels of these matrix components.22,23 These findings may explain the gender differences in thyroid cartilage mineralization. Other studies suggest that there are changes in chondrocyte shape through the thyroid cartilage with centrally located chondrocytes having an appearance similar to hypertrophic chondrocytes from growth plates, and that these chondrocytes may play a role in changes in the fibers, leading to vascularization and the formation of cartilage canals. These canals are mineralized, and then undergo a process of endochondral ossification. 24 Figure 1 shows picosirius red staining of thyroid cartilage sections from a young adult male. Picosirius red staining detects collagen I and III and may be birefringent into green/yellow areas depending on fiber thickness. Note the change in cell shape and density ( Fig. 1A ), as well as gradations in collagen I and III density, especially near areas of vascularization and possible ossification ( Fig. 1B ).
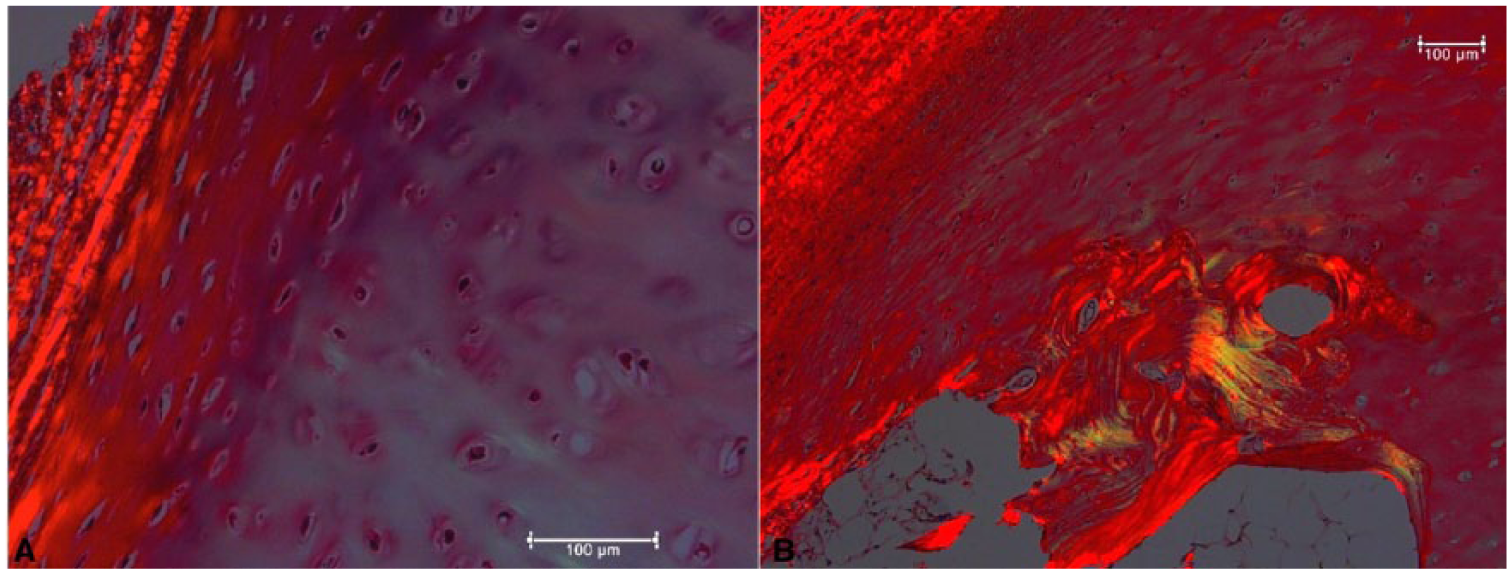
Picosirius red staining of the thyroid cartilage of a young male.
The compressive, tensile, and bending stresses applied to the thyroid cartilage are not well described. One study examined both the bending properties and hardness of individual lamina, and found that sites of ossifications increased the stiffness 23 but the heterogeneity of the structures suggests that more detailed mapping should be undertaken, including across the midline. It is not clear how areas of ossification will affect the mechanical properties of the thyroid cartilage, especially around the sites of insertion for the vocal ligament tendon. The shape of the chondrocytes and the arrangement of collagen fibrils have not been examined in thyroid cartilages, as these also contribute to mechanical properties.
Each inferior horn of the thyroid cartilage forms a synovial joint with the posterolateral aspect of the cricoid cartilage—the cricothyroid joint. These joints allow the thyroid and cricoid cartilages to approximate or separate at their anterior aspects, with posterior-tilting lengthening the vocal folds. There are 3 morphological categories of cricothyroid joint: type A joints are well-defined with a clearly identifiable cartilaginous protuberance, type B joints are poorly defined with only a small cartilaginous joint facet, and type C joints have a completely flat cricoid with no identifiable joint surface. 25 Type A joints enable a significantly greater range of movement than type B or C joints, and engineered cricothyroid joints should mimic this morphology.25,26 It is not known if the cartilage at this joint is similar to cartilage at larger articular joints.
Cricoid Cartilage
The cricoid cartilage is hyaline cartilage and forms a complete ring around the airway. It possesses 2 sets of paired synovial joints: the aforementioned cricothyroid joints, and a second pair of joints on the lateral surface of the superior cricoid lamina for articulation with the arytenoid cartilages.
In rabbits it has been shown that injury to the cricoid results in a straightening of the cartilage. This suggests that the cricoid as a whole has a balanced system of forces, but injury results in loss of force equilibrium and the resultant tension distorts the cartilage. 24 A study using three isolated human cricoids determined that an average radial expansile force of 97.25 N was required to fracture the cricoid cartilage. 27 The histology of the cricoid has not been examined in detail, but it is known to possess a thick perichondrial membrane. 28 Figure 2 shows a hemotoxylin and eosin–stained ( Fig. 2A ) and picosirius red–stained ( Fig. 2B ) sections from the cricoid cartilage of a young adult male. There are flattened chondrocytes parallel to the surface and round chondrocytes in the center of this section. Picosirius red staining detects areas with higher levels of collagens near areas of vascularization and possibly ossification. The cellular arrangement, as well as collagen fibril orientation and the presence of specific proteoglycans (PGs), could be key in tissue engineering a cricoid cartilage with this balanced system of forces.
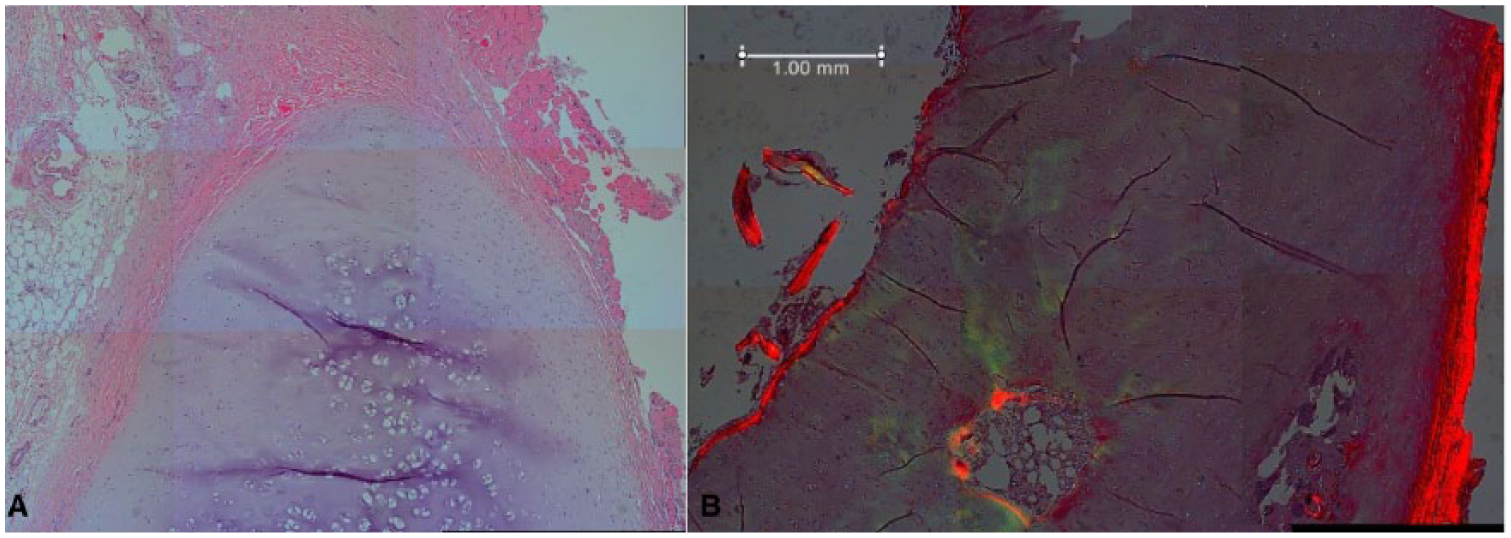
Section of cricoid cartilage stained with (A) hematoxylin and eosin or (B) picosirius red to show the intense staining of the collagen.
Biomechanical characterization of the equine cricoid cartilage has been initiated, although the shape is slightly straighter in the endoluminal region than in other species. 29 Three regions just below the sagittal ridge of the cricoid were measured using a confined compression assay. The mean aggregate modulus, a measure of stiffness, was 2.29 ± 0.09 MPa, which increased with age, and the rostral samples were 20% stiffer than the caudal samples. 29 The pooled cricoid permeability was measured to be 0.136 ± 0.174 × 10−14 m4/N·s. 29 These measurements suggest that the cricoid is not homogenous in its biomechanical factors, and thus there are regional differences in cellular and proteomic parameters.
Arytenoid Cartilage
The paired pyramidal arytenoids, sitting superolaterally to the cricoid laminae, contain two types of cartilage—hyaline and elastic.30,31 The base is formed of hyaline cartilage, primarily collagen II with high levels of PGs. The elastic cartilage is found superomedially, from the tip of the vocal process to the arytenoid apex, and contains collagen and PGs32,33 and an interspersing of elastic and collagenous fibers. 31 The transition between these zones is gradual. 30 This unique composition plays a critical role in the physiology of the arytenoids. The vocal ligament attaches to the vocal process, and during abduction or adduction of the true vocal fold, the elastic vocal process bends; furthermore, the entire superomedial section that contacts the other arytenoid is elastic cartilage.30,31 This arrangement allows for greater movement of the vocal process, protection of cartilage from potentially damaging vibrations, and cushioning of the arytenoids when they contact.30,34 The hyaline cartilage of the arytenoids also ossifies; however, investigation of the initiation and modulation of the mineralization process, as well as the roles of gender, are not well known. 23 Conversely, the elastic cartilage of the arytenoids normally does not ossify.
The saddle-like synovial cricoarytenoid joints allow the arytenoids to move in several directions, with a primary mechanism of switching the glottis from the respiratory (abducted) to the phonatory position (adducted) in a complex triplanar movement.34,35 A recent evaluation of the structural support of the cricoarytenoid found it lacks strong ligaments, with synovial folds the limit of fibrous support. Local musculature is the primary supportive mechanism, providing dynamic stabilization to protect against large vibratory forces.34,35
Despite the significant complexity of the arytenoids and the mechanics of their movement, they have not been studied in great depth.29-35 In the horse, the bulk aggregate modulus from the center of the cartilage has been measured using confined compression tests to be 0.42 ± 0.11 MPa and a permeability of 1.303 ± 0.191 mm4/N·s. 29 The elastic modulus of the elastic portion of the arytenoids is not known, although it is understood that this structure, along with the vocal ligaments and elastic nodules, must provide gradual changes in elasticity in order to allow the vocal folds to vibrate properly in the human. Knowledge of the cellular arrangements, collagen fibril and elastin orientation, and other proteins of the ECM are critical pieces of information for tissue engineering an arytenoid cartilage and its attachment to the dynamic vocal fold.
Corniculate and Cuneiform Cartilages
The corniculates and cuneiforms are small, paired elastic cartilages within the aryeptiglottic (AE) fold. Each corniculate cartilage is situated posterosuperiorly to the apex of the arytenoid cartilage, with which it interacts via a syndesmosis or diarthrosis joint, and is surrounded by a thin perichondrium. It variably attaches an arycorniculate muscle. 36
The rod-shape cuneiform cartilage is situated lateral and anterosuperior to the arytenoid cartilage, near the mucosa of the AE fold. 36 It may act to support the AE fold. Contemporary scientific evaluation of the structure of these cartilages is lacking in the literature, and the potential significance of their presence or absence in a tissue engineered larynx has yet to be investigated.
Epiglottic Cartilage
The epiglottic cartilage is a thin leaf-shaped sheet of elastic cartilage covered by a mucosa. Its stalk (petiolus) is attached to the luminal surface of the thyroid cartilage (in the midline below the superior thyroid notch) by the thyroepiglottic ligament, and it is bound to the hyoid by the hyo-epiglottic ligament, with a free edge superiorly. It projects obliquely upward to cover the laryngeal inlet. During deglutition the epiglottis flips posteroinferiorly, protecting the airway, although its presence is not obligatory for adequate swallowing. 37
The epiglottis itself may be described by the angle formed between the aryepiglottic folds and the midline. 37 The epiglottis is covered by a perichondrium,33,38 with flattened chondrocytes near the surface and round chondrocytes toward the center. 39 The number of chondrocytes and elastic fibers does not vary significantly, but the amounts of acid mucopolysaccharides may decrease in some epiglotti. 37 Immunohistochemical staining detected Collagen II and V in both the interterritorial matrix and pericellular regions of the cartilage, collagen I and X in the pericellular matrix, and elastin in the interterritorial matrix. 38 Quantitative staining detected lower amounts of total glycosaminoglycans (GAGs) than in other types of cartilage. 38
Several material parameters have been measured in the epiglottis. Cartilage samples from several anatomical sites of young rabbits were tested using a biphasic indentation assay. The epiglottis had an aggregate modulus of 0.290 ± 0.174 MPa, a Poisson’s ratio of 0.250 ± 0.115 (a measure of compressibility), and a permeability of 8.01 ± 5.16 × 10−16 m4/N·s. 4 The aggregate modulus of the epiglottis varied significantly from that of the auricular elastic cartilages and nasal hyaline cartilage. 4 There were no significant differences in the other parameters between these cartilage types. In order to determine if their tissue engineered construct matched the native tissue, some labs have examined a number of variables, including collagen content, GAG content, elastin content,4,33 water content, 4 and the aggregate modulus (386.62 kPa). 4 Furthermore, the contributions of the mucus glands and deposits of adipose tissue at the base of the epiglottis contribute to the properties of the entire epiglottis, and must be considered when designing a tissue engineered construct. 33
Tracheal Cartilage
Each tracheal cartilage is an incomplete ring of hyaline cartilage with the trachealis muscle connecting the posterior ends, allowing expansion during respiration and swallowing. The first tracheal cartilage is the broadest, occasionally bifurcating at the dorsal ends, and may fuse with the cricoid. 40 Tracheal rings have a greater amount of collagen at the surface, and the collagen is arranged in fibrils parallel to the surface with layers running circumferentially and axially.41,42 These layers are detected on both the luminal and abluminal surfaces. This layering of the collagen fibers is less evident by scanning electron microscopy in the deeper zones of the cartilage and the fibrils may run perpendicular to the surface. 41 The central core of the tracheal cartilage contains a higher proportion of PGs. 41 Water content and hydroxyproline content (as a measure of collagen presence) decrease with age but PG amount shows no consistent changes with age. 41
The tensile strength of tracheal cartilage has been measured in the circumferential direction (the direction of the plane of stress produced by contraction of the trachealis muscle or by dilation). At the abluminal surface, the average tensile modulus (Young’s modulus) is 13.6 ± 1.5 MPa. The modulus decreases as the depth from the surface increases, and at approximately 600 microns from the surface, the average tensile modulus is 4.6 ± 1.7 MPa. 41 Using the central portion of the tracheal cartilage (2.1 mm thickness, 6.6 mm length, 6 mm width), a tensile modulus of 3.33 MPa exists. 42 An average value has been suggested at 4.4 MPa. 43 There are increases in the stiffness of tracheal cartilage with age. 44
Variations in the tensile moduli and the coefficient of viscosity between tracheal cartilages have been found to impact subglottal acoustic input impedance, which can be observed empirically in subglottal acoustic spectra. 15 Therefore, tissue engineered laryngotracheal cartilage should be similar to native tracheal cartilage in these features.
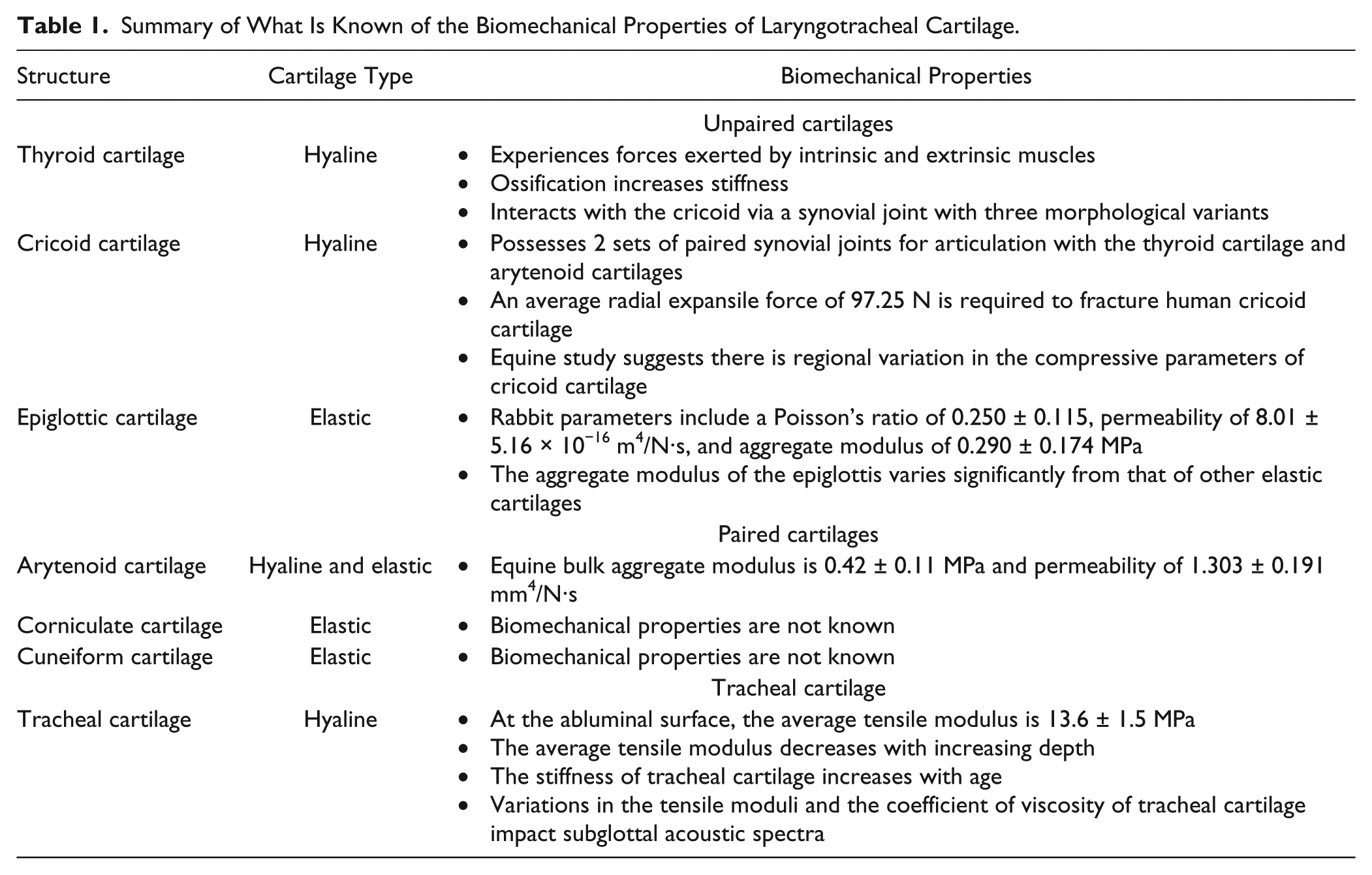
Table 1 summarizes what is known of the biomechanical properties of laryngotracheal cartilage.
Summary of What Is Known of the Biomechanical Properties of Laryngotracheal Cartilage.
Cartilage Proteomics
The proteomic composition of the laryngeal cartilages has not yet been elucidated. However, knowledge of the proteomic composition of various other cartilages entails information that may be crucial to both developing better scaffold biomaterials for tissue engineering of laryngeal cartilage and detailing benchmarks to determine if a tissue engineered construct has the proteomic composition of native tissue.3,5,45-47 Mass spectroscopy and proteomics provide an efficient method to monitor the complete profile of proteins in a tissue.
Although proteomic studies have been performed on a wide number of normal human tissues, cartilage and bone present some technical difficulties because of the highly cross-linked ECM, which may not be completely solubilized.16,48,49 One study on cartilage found differences during the development of the murine femoral cartilage ECM between proliferative chondrocytes and older, hypertrophic cartilage. 50 Changes in cell shape were demonstrated, as well as differential expression of 146 proteins, including several collagens, several protein components of PGs, and other ECM proteins. 51
In a second study, a comparison of the proteome from eight cartilage sources (hyaline—femoral head, humeral head, tibial plateau, costal, and tracheal; fibrocartilage—meniscus, disc annulus fibrosus, and disc nucleus pulposus) isolated in toto from adult humans has been performed. 16 This study identified more than 300 different proteins with 120 to 198 proteins detected in any one tissue. There were significant variations in the protein composition of the ECM of cartilage from different anatomical sources, such as collagens I and III between the tracheal and articular cartilage. Other proteins with significant differences between these 2 cartilages include fibronectin, decorin, lumican, cartilage oligomeric protein (COMP), and lysozyme C. Comparison of rib and tracheal cartilage, although much more similar to each other, still had significant differences in COMP and lysozyme C. 16 Thus even hyaline cartilage has differences in its proteomic composition with anatomical location, possibly because of differences in the various stresses experienced by each cartilage.
In another study, normal human articular cartilage was obtained from the medial and lateral tibial plateaus and sectioned to facilitate analysis at varying cartilage depths. 48 This study discovered 277 quantifiable proteins, of which 70 ECM proteins demonstrated unequal distribution between the different articular cartilage layers in several different patterns. Further studies using hip and knee articular cartilage samples were able to fractionate the ECM from the chondrocytes and then used trypsin digestion to disperse the insoluble collagen network. 49 These studies detected 425 proteins, with differences in the proteins present in different zones of the articular cartilage and between normal and osteoarthritic tissue. 49 These findings contribute to the concept that there are spatial patterns to protein expression in articular cartilage that correlate with the functional roles of that region of the cartilage, whether it is lubrication and resistance to shear stress at the superficial layer, a shock-absorbing layer with elastic properties, or to develop connections to subchondral bone or cartilages with different stressors.16,47
Table 2 summarizes knowledge of the proteomic composition of cartilage.
Summary of What Is Known of the Proteomic Composition of Cartilage.

Future Work
The concepts described have important translational implications for laryngotracheal cartilage tissue engineering. There is evidence that the structure of tracheal cartilage ECM significantly differs to that of articular cartilage, therefore detailed and specific laryngotracheal cartilage research will be required to adequately understand the functional implications of its microstructure. Furthermore, the protein composition of the ECM of various laryngeal cartilages may contain significant differences in a few key proteins, creating the specific biomechanical properties that are utilized in normal function. Knowledge of the proteome should provide information on which proteins are especially critical in building a scaffold for tissue engineered cartilage that provides a hospitable environment for chondrocytes to remodel. The ECM within a single cartilage may also vary greatly across its gross structure, including the orientation and density of collagen and elastin fibrils, arrangement and density of chondrocytes, and concentration of GAGs through the depth of the cartilage. Proteomic and spatial localization information will help define the roles of various proteins in determining the material properties of the tissue. There are also the joint structures that must be considered and the areas of muscle insertion, which may be different from articular sites of insertion. Furthermore, knowledge of the regions that ossify and the factors that modulate this process may be of interest in aging studies. Proteomics may also contribute information about biomarkers to measure the maturation of tissue engineered cartilages during in vitro culture before implantation.
Mechanically, very little is known of these cartilages. Tissue stiffness in both tensile and compressive modes is a critical parameter since the stiffness of both the thyroid and arytenoid cartilages may affect vocal performance, especially in older tissues where ossification will affect the stiffness. The cricoid and tracheal cartilages must have the proper stiffness to provide airway patency. The compressive and tensile properties of the human cricoid, tracheal, arytenoid, and epiglottic cartilages are unknown. Bending modulus and joint lubrication are other material properties that are critical parameters of cartilage function. The bending modulus at several points on the epiglottis is critical to its function while joint lubrication is important to the movement of the cricoarytenoid and cricoidthyroid joints. Testing of these cartilages must be correlated with cell density, GAG concentration, distribution of critical proteins, collagen and elastin networks, and sites of ossification. This knowledge will contribute significantly to the ability of personnel in regenerative medicine to reconstruct the laryngotracheal system.
Conclusion
Laryngotracheal tissue engineering aims to create biomimetic scaffolds for the replacement of this complex organ. Although a great deal of progress has been made, the development of a clinically applicable, tissue engineered larynx has yet to occur. To facilitate further development, more detailed investigation of the normal mechanical, cellular, and proteomic properties of laryngotracheal cartilage is required.
Footnotes
Acknowledgments and Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.