
Abstract
Introduction
We previously showed that articular chondrocytes (ACs) have immune privilege and immunomodulatory functions like those of mesenchymal stem cells. To elucidate these mechanisms, we focused on interleukin-2 (IL-2), which plays critical roles in lymphocyte mitogenic activity. The purpose of this study was to explore whether ACs affect the role of IL-2 underlying immunomodulatory functions.
Material and Methods
Irradiated human ACs from osteoarthritis donors were used. Third-party ACs were added to the mixed lymphocyte reaction (MLR) with or without recombinant human IL-2 (rhIL-2), and the levels of IL-2 and the soluble form of the IL-2 receptor α (sIL-2Rα) protein in supernatant were measured by enzyme-linked immunosorbent assay. Recombinant human IL-2 (rhIL-2) was also added to the MLR. To detect the expression of IL-2 receptor α (CD25) on lymphocytes in the MLR, flow cytometric analysis was performed. Last, ACs and allogeneic activated CD4+ T cell were co-cultured, and the expression of CD25 on activated T cells was examined by flow cytometry.
Results
Third-party ACs significantly inhibited the MLR and reduced the level of sIL-2Rα in a dose-dependent manner, but did not affect the concentration of IL-2. Exogenous rhIL-2 accelerated MLR but did not rescue the inhibitory effect of ACs. ACs inhibited the expression of CD25 on activated CD4+ T cells.
Discussion
Our results showed that third-party ACs inhibited the proliferation of allogeneic activated lymphocytes, thereby inhibiting production sIL-2Rα, although ACs did not affect IL-2 secretion from lymphocytes. Also, ACs inhibited CD25 expression on activated CD4+ T cells. Thus, ACs inhibited the immune response of allogeneic lymphocytes by inducing IL-2 nonresponsiveness.
Introduction
Immunomodulatory properties of mesenchymal stem cells (MSCs) have been observed in T cell proliferation assays using a variety of stimuli, including mitogens, CD3/CD28, and alloantigens.1,2 Such immune suppression occurs irrespective of donor source, including settings using “third-party” MSCs. The immunosuppressive action of MSC is of clinical relevance of in allogeneic stem cell transplantation, because it can theoretically reduce the incidence and severity of graft-versus-host disease (GVHD). 3
MSCs and human articular chondrocytes (ACs) possess quite similar immunological characteristics. 4 We previously demonstrated that ACs have immunomodulatory functions in vitro. In that study, ACs were nonalloreactive and inhibited the mitogenic activity of polyclonally activated CD4+ T cells via cell-to-cell interaction, even in cells from aged subjects or patients with osteoarthritis condition.4,5 Autologous chondrocyte implantation (ACI) is the standard option for cartilage repair. However, ACI is limited by cell number and requires a waiting period. An allogeneic cell source could resolve these problems. However, little is known about the biological mechanisms underlying the immunomodulatory functions of these cells.
Interleukin-2 (IL-2), which was discovered in 1976 as a T-cell growth factor activity in the supernatants of activated T cells, 6 is produced primarily by CD4+ T cells following their activation by antigen. 7 IL-2 induces expression of both IL-2Rα and IL-2Rβ,8,9 and its targets are T cells, B cells, and natural killer (NK) cells. Three different IL-2 receptor chains exist; in various combinations, they generate low-, intermediate-, and high-affinity IL-2 receptors. 10 The ligand-specific IL-2 receptor αchain (IL-2Rα, CD25), which is expressed on activated but not non-activated lymphocytes, binds IL-2 with low affinity; IL-2Rβ(CD122) and IL-2Rγ(γc, CD132) form an IL-2Rβ/γ complex, primarily on memory T cells and NK cells, that binds IL-2 with intermediate affinity; and when all three receptor chains are co-expressed on activated T cells and regulatory T cells (Treg), IL-2 is bound with high affinity. 10 The intermediate- and high-affinity receptor forms effectively transduce IL-2 signals.
To elucidate the mechanisms underlying the immunomodulatory functions of ACs, we investigated the role of IL-2, which plays the critical role in the mitogenic activity of lymphocytes, focusing on its immunomodulatory functions.
Material and Methods
Human Articular Chondrocyte Culture
Human articular chondrocytes (ACs) were isolated from 10 osteoarthritis knee joints from subjects aged 64 to 75 years at the time of total arthroplasty. Informed consent was obtained from all subjects, and the study received institutional review board approval. The cartilage was minced and digested by treatment with Pronase for 90 minutes, followed by overnight incubation in HL-1 medium (BioWhittaker) with 0.5 mg/mL CLS4 collagenase (Worthington), 0.3 mg/mL hyaluronidase (Type IV hyaluronidase; Sigma), and 50 μg/mL ascorbic acid. After enzymatic dissociation, the chondrocytes were cultured in Dulbecco’s modified Eagle’s medium (DMEM; Sigma) containing 10% fetal bovine serum (FBS). Cells at passage 2 were used for all experiments.
Mixed Lymphocyte Reaction (MLR)
Peripheral blood mononuclear cells (PBMCs) were collected after Histopaque-1077 (Sigma) centrifugation of whole blood from healthy donors. In order to prepare MLR culture, 5 × 105 irradiated (15 Gy) stimulator PBMCs were co-cultured with 5 × 105 responder allogeneic PBMCs harvested from individual donors in RPMI 1640 medium (Sigma) supplemented with 10% FBS and antibiotics (streptomycin and penicillin) (complete medium) for 4 days. Samples were prepared in triplicate.
Cell Proliferation Assay
First, irradiated (30 Gy) third-party ACs were plated in triplicates onto 96-well round bottom plates at 0, 4 × 103, 2 × 104, or 1 × 105 cells/well in 100 μL of complete medium. Second, recombinant human IL-2 (rhIL-2; PeproTech) was diluted at several concentrations with complete medium, and 10-μL aliquots were added to the plate to final concentrations of 0, 5, 50, or 500 IU/mL. Third, 100 μL of prepared MLR culture (containing 2 × 105 cells) was added to each well and co-cultured for 4 days. Then, 18 hours before of the end of culture, 1 μCi/well of 3H-thymidine (Amersham) was added to each well, and the proliferation of responder lymphocytes was measured. Results are expressed as mean counts per minute ± standard deviation (mean cpm ± SD) from triplicate samples.
Cytokine Enzyme-Linked Immunosorbent Assay (ELISA)
To quantify IL-2 and sIL-2Rα protein levels in cultures, 100 μL of supernatant were taken from each culture at the end of the cell proliferation assay, and the concentration of both proteins was quantified using the appropriate ELISA kit (R&D Systems).
Activating CD4+ T Cells Proliferation Assay
PBMCs were isolated from healthy volunteers and enriched using Histopaque-1077, and purified CD14−CD4+ T cells (CD4+ T cells) were isolated using MACS Magnetic cell sorting system (Miltenyi Biotec) and MicroBeads conjugated to anti-human CD4 and CD14 mAbs (Miltenyi Biotec). Purified CD4+ T cells (responder) were activated by simultaneous ligation to the T-cell receptor complex and co-stimulatory molecules with 10 ng/mL CD3 mAbs (BD Pharmingen) and 5 μg/mL of CD28 mAbs (BD Pharmingen), and then co-cultured with ACs (stimulator) for 4 days at a responder/stimulator (R/S) ratio of 1/1 to 25/1. The proliferation of these activated CD4+ T cells was measured in the same manner as in the MLR assay.
Flow Cytometric Analysis
To investigate the expression of IL-2 receptor α, (FITC-conjugated mouse anti-human CD25, Beckman Coulter, Inc.), β (PE-conjugated mouse anti-human CD122, BD Pharmingen), and γ (PE- conjugated mouse anti-human CD132, BD Pharmingen) on activated CD4+ T cells in the proliferation assay at an R/S ratio of 1/1, flow cytometric analysis was performed with the indicated antibodies 24, 48, and 72 hours after the start of culture. Quantitative flow cytometric analyses were performed using FACSCalibur (Becton Dickinson). A total of 10,000 live cells were examined, and the data were analyzed using the CellQuest software (Becton Dickinson).
Statistical Analysis
Data are expressed as means ± SD and analyzed using StatView version 5.0 (SAS Institute Inc.). Comparisons were made by one-way ANOVA with Scheffe’s post hoc test. A value of P < 0.05 was considered significant.
We examined 10 samples in triplicate and showed representative data.
Results
Third-Party ACs Significantly Inhibited MLR in a Dose-Dependent Fashion
Third-party ACs were co-cultured with MLR culture for 4 days. They inhibited lymphocyte proliferation in a dose-dependent fashion ( Fig. 1 ). A stimulator index (SI) >10 was considered a positive response in the MLR. SI = [(responder + irradiated stimulator) − irradiated stimulator] cpm / (responder) cpm.
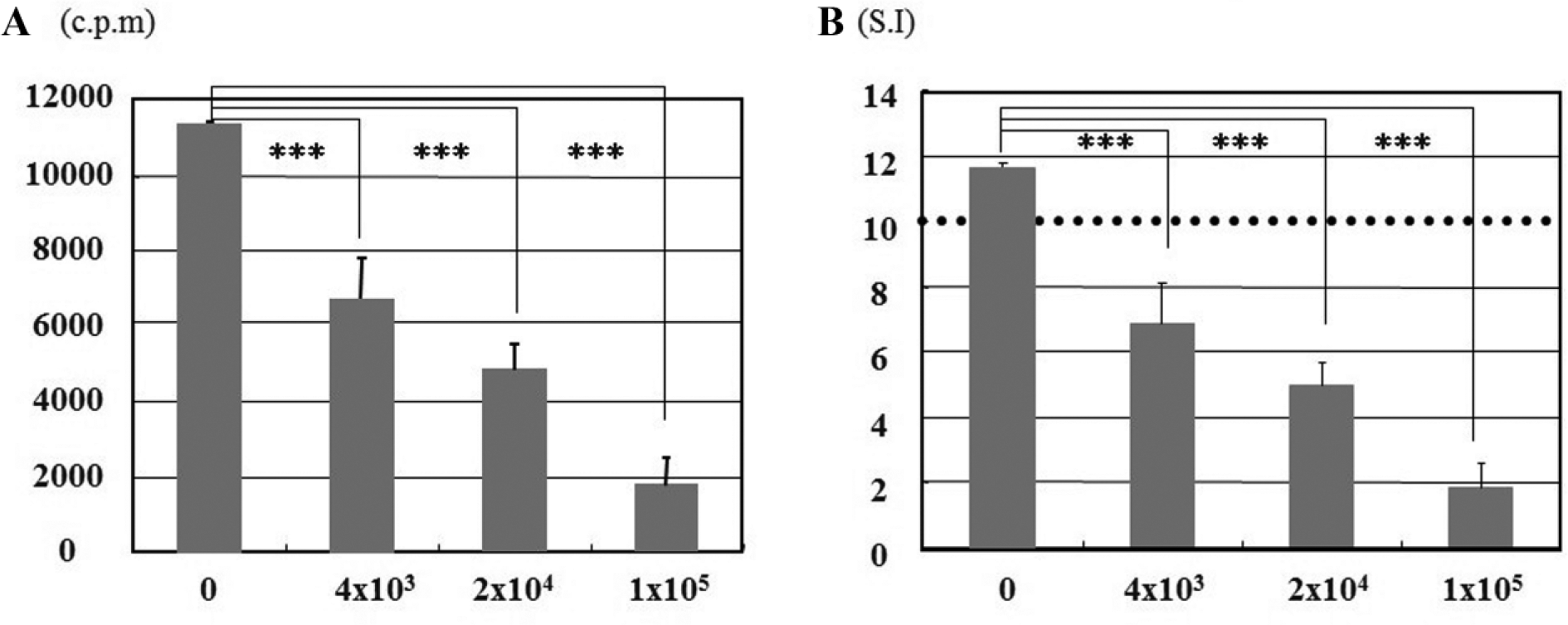
Cell proliferation assay. Third-party articular chondrocytes (ACs) were co-cultured with mixed lymphocyte reaction (MLR) cultures for 4 days. X-axis indicates the number of ACs, and Y-axis indicates the counts per minute (cpm) (
Third-Party ACs Did Not Affect the Concentration of IL-2 Protein, whereas sIL-2Rα Protein Level Was Significantly Reduced in Culture Supernatants
Supernatant was taken from each culture at the end of the cell proliferation assay, and the concentrations of IL-2 and sIL-2Rα were measured. IL-2 level increased in a dose-dependent manner, but the difference was not statistically significant ( Fig. 2A ). This observation demonstrated that third-party ACs did not affect the concentration of IL-2 protein, although they did inhibit the MLR ( Fig. 1 ). The concentration of sIL-2Rα protein was significantly reduced in a dose-dependent fashion ( Fig. 2B ).
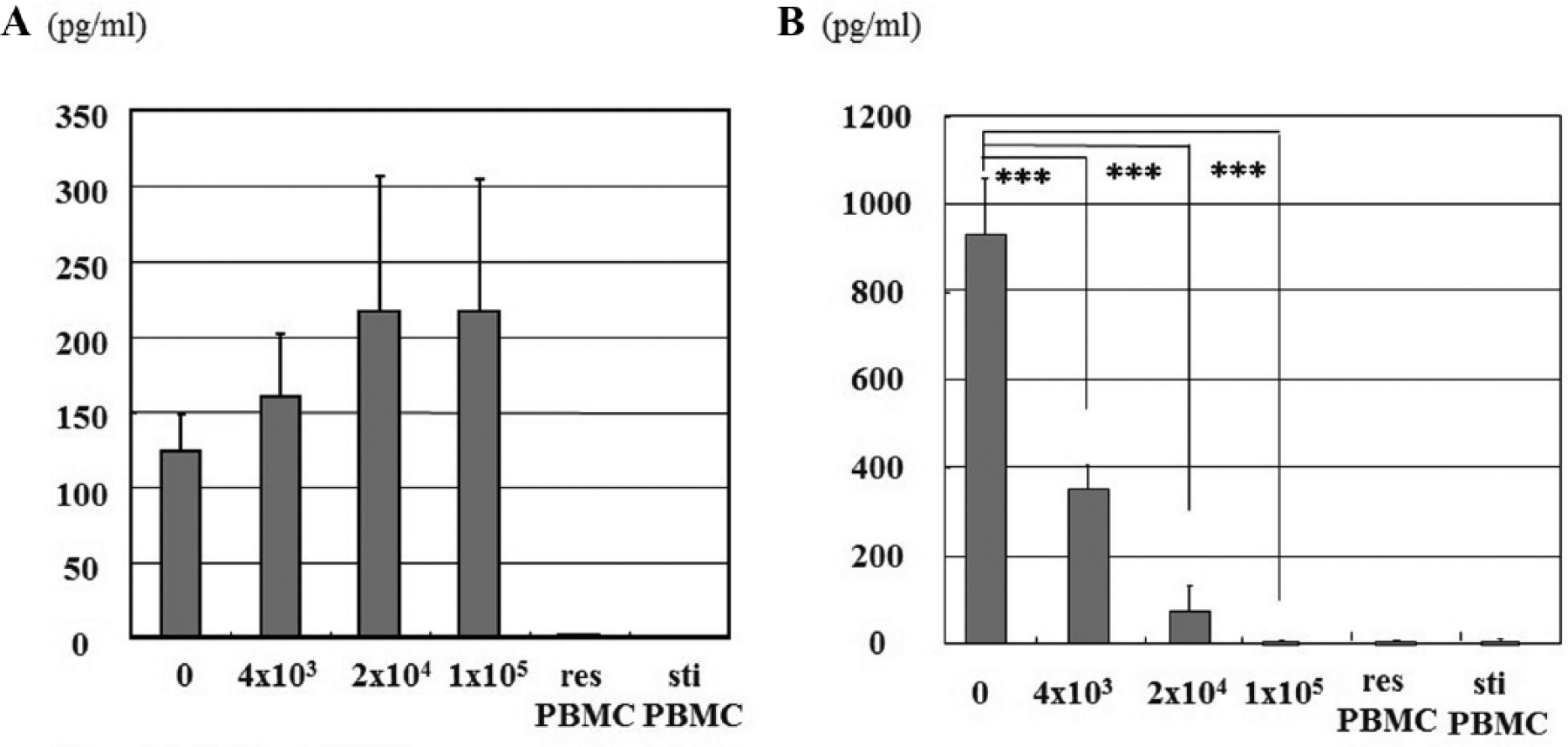
Cytokine enzyme-linked immunosorbent assay (ELISA). Concentrations of IL-2 (
Exogenous rhIL-2 Significantly Accelerated MLR in a Dose-Dependent Fashion but Barely Overcame the Inhibitory Effect of ACs
We wanted to confirm whether the concentration IL-2 in supernatant of MLR could lead to inhibition. Therefore, we co-cultured third-party ACs with MLR cultures for 4 days in the presence of 0, 5, 50, or 500 IU/mL rhIL-2. Exogenous rhIL-2 significantly accelerated MLR in a dose-dependent fashion but barely overcame the inhibitory effect of ACs in ACs dose-dependently ( Fig. 3 ).
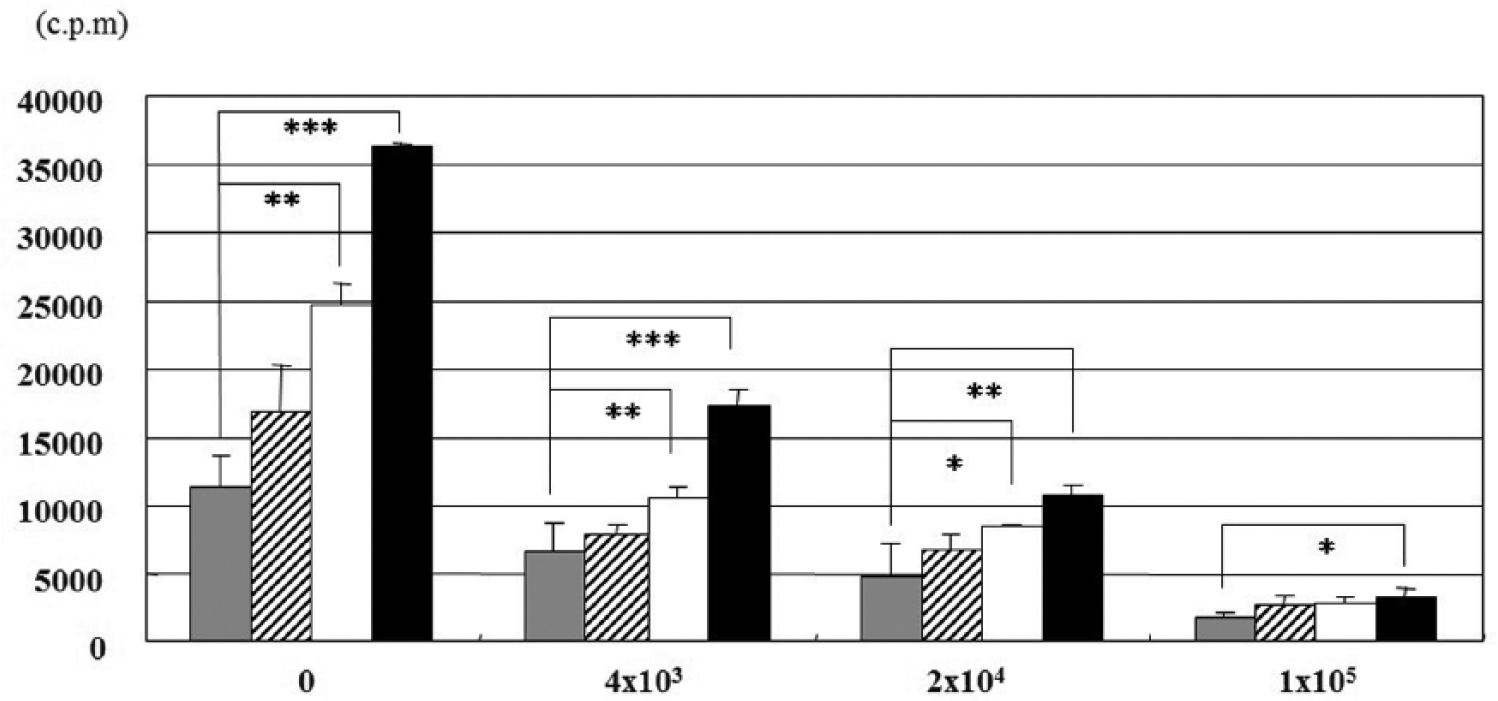
Cell proliferation assay. Third-party articular chondrocytes (ACs) were co-cultured for 4 days with mixed lymphocyte reaction (MLR) culture containing 0 (gray bars), 5 (hatched bars), 50 (white bars), or 500 (black bars) IU/mL rhIL-2. X-axis indicates the number of ACs. Y-axis indicates counts per minute (cpm).
ACs Inhibited Proliferation and Downregulated the Expression of IL-2 Receptor α in Activated CD4+ T Cells
Allogeneic activated CD4+ T cells were co-cultured with ACs at a ratio of 1/1. ACs inhibited the proliferation of activated CD4+ T cells ( Fig. 4A ).
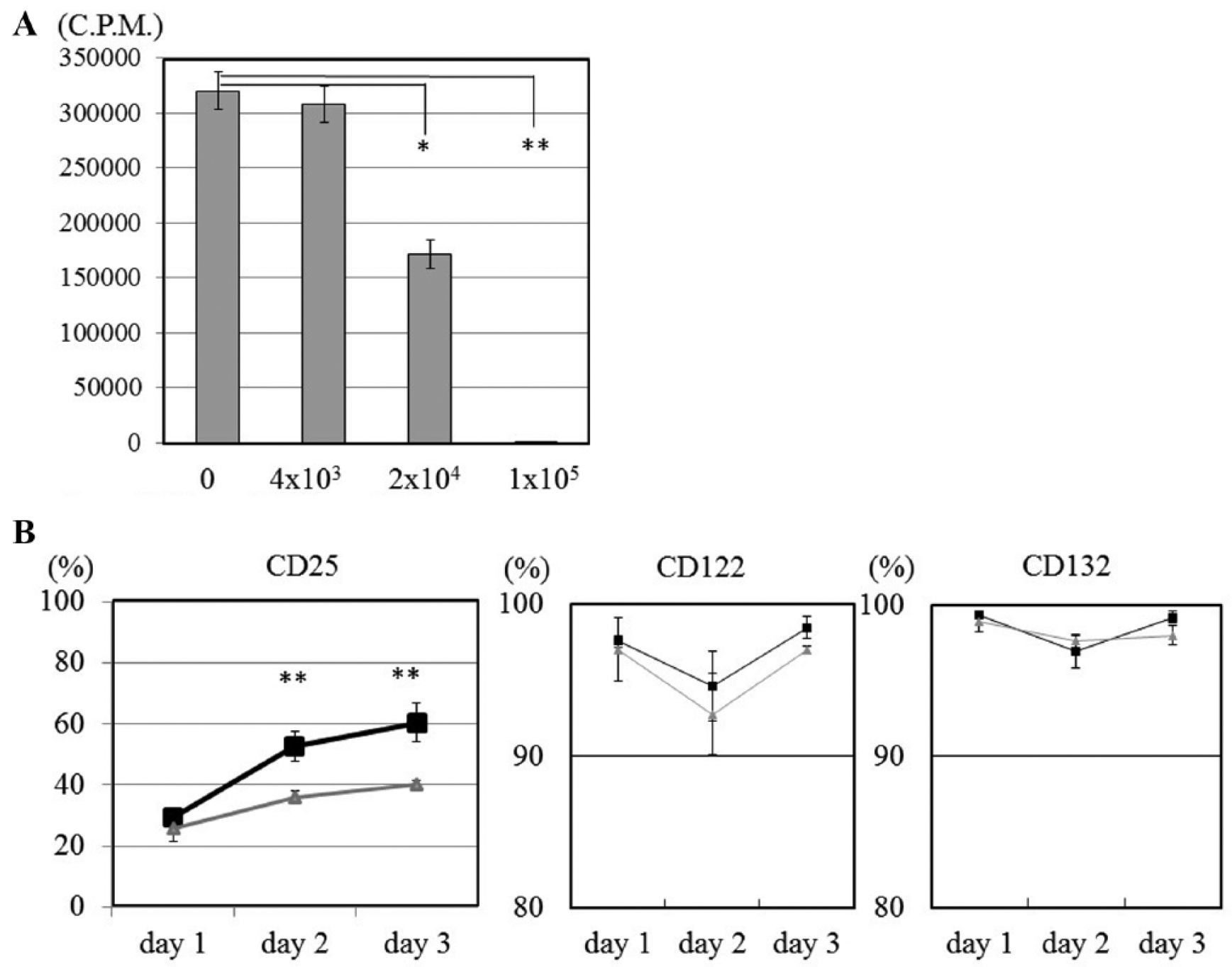
(
In flow cytometric analysis, the expression of CD25 (IL-2 receptor α) on activated CD4+ T cells co-cultured with ACs significantly decreased relative to the level on activated CD4+ T cells cultured alone, 10% at 48 hours and 20% at 72 hours. The expression of CD122 (IL-2 receptor β) and CD132 (IL-2 receptor γ) was not affected by ACs ( Fig. 4B ).
Discussion
To investigate immunomodulatory functions, we cultured third-party ACs with MLRs. ACs inhibited the proliferation of allogeneic antigen activated lymphocytes ( Fig. 1 ), and we confirmed the immune-suppressive effect of ACs. In this condition, ACs did not affect IL-2 secretion, but ACs inhibited sIL-2Rα secretion in a dose-dependent fashion ( Fig. 2 ). Even if exogenous rhIL-2 was present in the MLR, the inhibitory effect of ACs overcame the acceleration of the MLR. Thus, human ACs induce IL-2 nonresponsiveness in allogeneic lymphocytes
sIL-2R is secreted by activated CD4+ T cells,11,12 and the sIL-2R concentration has been proposed as a marker of rejection episodes after organ transplantation.13,14 Acute renal allograft rejection is mediated by T lymphocytes; T cells expressing cell surface IL-2R were present in the kidneys of renal allograft recipients. 15 Prediction of sIL-2R levels might aid in early detection of rejection episodes. 16 In this study, the lower levels of sIL-2Rα produced by third-party ACs in the MLR suggest that allogeneic ACs are difficult to reject, supporting the view that ACs escape recognition by alloreactive T cells.
Furthermore, we also showed that expression of CD25 (i.e., membrane-bound IL-2Rα) was significantly lower in ACs cultured with activated CD4+ T cells than in controls (P < 0.05). IL-2Rα is expressed on activated lymphocytes. Therefore, this observation indicates that ACs inhibit the activation of stimulated CD4+ T cells. IL-2Rα binds IL-2 with low affinity but contains no signaling domains. However, after IL-2 is bound to the α/β heterodimer, the γc chain is recruited to the IL-2/IL-2 receptor quaternary complex to form a very stable interaction. Anti-CD25 (daclizumab) monoclonal antibody therapy has been approved for many years for the prevention of allograft rejection. 17 Therefore, our observation that ACs downregulated expression of IL-2Rα on activated CD4+ T cells suggests that ACs could prevent allogeneic reject immune reaction.
Consistent with our results, Le Blanc et al. 18 reported that MSCs inhibit the expression of IL-2Rα and CD38 on phytohemagglutinin (PHA)-activated lymphocytes. Rasmusson et al. 19 reported that MSC inhibition differs depending on T-cell stimuli. PHA-stimulated PBMCs suppressed by MSCs decrease IL-2 secretion, whereas sIL-2R and IL-10 levels remain unchanged. Allogeneic stimulated PBMCs in mixed lymphocyte cultures exhibited more complex behavior: IL-2, sIL-2, and IL-10 levels were elevated, and addition of IL-10 antibodies further inhibited proliferation. Park et al. 20 reported that murine mesenchymal stem cells suppress T-lymphocyte activation by producing matrix metalloproteinases, resulting in IL-2Rα cleavage. Several studies indicated that the immunosuppressive effect of MSCs is mediated by either direct cell-to-cell contact 21 or by soluble factors22,23 produced by MSCs or immune cells; examples include the immunosuppressive cytokines interleukin 10 (IL-10) and transforming growth factor-β (TGF-β), as well as hepatocyte growth factor (HGF), prostaglandin E2 (PGE2), nitric oxide (NO), or indoleamine 2,3-dioxygenase (IDO).24-27 However, controversy persists regarding concerning the mechanism underlying the immunosuppression mediated by soluble factors secreted from MSCs.23,28 These differences could be explained by the fact that the experiments were performed in different species using different protocols. Major differences exist between murine and human MSCs; unlike their human equivalents, murine MSCs do not express major histocompatibility complex (MHC) class II. 21
Previously, we reported that immunosuppressive effect of ACs is mediated by direct cell-to-cell contact; furthermore, chondrocytes express negligible levels of MHC class II molecules and lack B7-1 and B7-2 antigens, which are co-stimulatory molecules that promote CD4+ T-cell proliferation.4,5 In these reports, we proposed that the presence of negative co-stimulatory molecules in chondrocytes reflects immune privilege. However, recent studies showed that expression of MHC molecules in allogeneic transplanted chondrocytes is upregulated in the early stage, and that the tissue fluid environment of allogeneic chondrocytes in vivo affects their immunological properties. 29 Although immune responses and the mechanism of allogeneic ACs remain unclear, the results of this study provide new explanation of the immunosuppressive mechanism.
One of the study limitation was that we did not characterize gene expression or other surface markers of the cultured T-cells, although sIL-2Rα was the marker of T-cell activation. It is remotely possible that irradiated ACs metabolized the sIL-2Rα, as an alternative possibility ACs attenuating T-cell activation. sIL-2Rα levels were lower in the presence of ACs, but the mechanisms remain to be elucidated.
IL-2 modulates differentiation of other immune effector cells via regulation of cytokine receptor expression. 30 Therefore, the immune system must be more complicated in vivo and in the assay using PBMC in vitro. IL-2 promotes Th1 differentiation by inducing IL-12Rβ (and IL-12Rβ1), promotes Th2 differentiation by inducing IL-4Rα, inhibits Th17 differentiation by inhibiting gp130 (and IL-6Rα), and drives Treg differentiation by inducing IL-2Rα. IL-2 also potently represses IL-7Rα, which decreases survival signals that normally promote cell survival and memory cell development. As I mentioned before, our results have the possibility of allogeneic cell transplantation for joint repair. Furthermore, this research will help us understand tumor and inflammatory disease.
In summary, we showed that ACs downregulated membrane-bound IL-2 receptor α (CD25) expression on allogeneic activated CD4+ T cells. Moreover, exogenous rhIL-2 significantly accelerated MLR but barely overcame the inhibitory effect of third party ACs, indicating the possibility of that ACs induced IL-2 nonresponsiveness to allogeneic lymphocytes.
Footnotes
Acknowledgment and Funding
The authors thank the patients who gave consent for their tissue to be used for research purposes at the Department of Orthopedic Surgery, Asahikawa Medical University Hospital.
Declaration of Conflicting Interests
The author(s) declared no potential conflict of interest with respect to the research, authorship, and/or publication of this article.
Ethical Approval
Ethical approval was obtained from Asahikawa Medical University (No.162-1400).
Informed Consent
Written informed consent was obtained from all subjects before the study.