
Abstract
Objective
Normal physiological movement creates different weightbearing zones within a human knee: the medial condyle bearing the highest and the trochlea bearing the lowest weight. Adaptation to different physiological loading conditions results in different tissue and cellular properties within a knee. The objective of this study was to use microarray analysis to examine gene expression differences among three anatomical regions of human knee articular cartilage at baseline and following induction of an acute impact injury.
Design
Cartilage explants were harvested from 7 cadaveric knees (12 plugs per knee). A drop tower was utilized to introduce injury. Plugs were examined 24 hours after impact for gene expression using microarray. The primary analysis is the comparison of baseline versus impacted samples within each region separately. In addition, pairwise comparisons among the three regions were performed at baseline and after impact. False discovery rate (FDR) was used to evaluate significance of differential gene expression.
Results
In the comparison of before and after injury, the trochlear had 130 differentially expressed genes (FDR ≤ 0.05) while the condyles had none. In the comparison among regions, smaller sets of differentially expressed genes (n ≤ 21) were found, with trochlea being more different than the condyles. Most of more frequently expressed genes in trochlea are developmental genes.
Conclusions
Within the experimental setup of this study, only the trochlea was displaying an acute genetic response on injury. Our data demonstrated the regional-specific response to injury in human articular cartilage.
Introduction
Articular cartilage is a connective tissue made up of chondrocytes in a collagen-rich extracellular matrix (ECM), which functions to support and distribute physical weightbearing loads across the surface of joints. Articular cartilage is essential in reducing stress on subchondral bone and minimizing joint friction 1 ; however, articular cartilage has poor blood supply, limiting its capacity for self-repair. 2 Mechanical factors, such as impact injuries often seen in trauma, have been considered one of the major causes of the resulting cartilage degeneration. If left untreated, cartilage injuries can lead to functional loss and disability, especially in the high-risk athletic population.3-8
In sports, cartilage degeneration often occurs secondary to other injuries such as ligament tears, meniscal tears, patellar dislocations, and osteochondral injuries.9,10 The most common injuries that occur simultaneously with articular cartilage lesions are medial meniscal tears (37%) and anterior cruciate ligament ruptures (36%). 11 In a population of athletes having anterior cruciate ligament reconstruction, nearly half of patients also presented with articular cartilage defects in the femoral condyle. 10 Furthermore, magnetic resonance imaging of professional basketball players showed articular cartilage abnormalities in 89% of asymptomatic players 7 ; clearly, articular cartilage defects represent a significant concern to the sports community. In addition, acute articular cartilage impact injuries can eventually progress to osteoarthritis (OA). OA of the knee is estimated to be 40% heritable (OA related to aging) 12 ; however, posttraumatic or secondary OA is presumably due to degenerative changes induced by trauma.13-15
Within the knee joint, high and low weightbearing regions of cartilage exist. 16 In addition, as body weight shifts during the walking gait, articular cartilage receives alternating periods of loading and unloading, providing mechanical conditioning within the native environment. 17 Over time, the combination of these 2 factors results in different tissue and cellular properties within articular cartilage.18-21 This may make the articular cartilage of each preconditioned region respond differently to mechanical injury.
Increasing chondrocyte death and activities of pro-inflammatory cytokines and catabolic enzymes are early detrimental events of injured cartilage.22-30 Besides those events, other changes also occur at the molecular level. Burton-Wurster et al. 31 identified 172 significantly expressed genes in canine articular cartilage 24 hours after in vitro impact injury. Fourteen days following mechanical injury of porcine patella articular cartilage explants, Ashwell et al. 32 found 39 significantly differentially expressed genes, several of which associated with matrix molecules, and cell proliferation, and inflammatory response.
Although these prior studies have characterized the molecular changes in articular cartilage following impact injury, they did not account for the possibility of baseline expression differences amongst anatomical regions of knee articular cartilage and how they might in turn affect injury response. Several studies found differences in cell density and matrix morphology between high and low weightbearing zones.33-37 To investigate these baseline differences at the genetic level, we conducted an in vitro study in human cadaver knees. We used microarray analysis to examine gene expression differences among 3 anatomical regions of human knee articular cartilage at baseline and following induction of an acute impact articular cartilage injury. We hypothesized that due to varying weightbearing requirements of different articular cartilage regions, there will be significant differences in gene expression across knee articular cartilage at baseline and this will affect how a specific area’s gene expression responds to injury.
Materials and Methods
Human Articular Cartilage Model
Fresh cadaveric human knee articular cartilage with no signs of arthritis on the articular surface was used in this study. En-bloc knee joints (dissected mid femur to mid tibia) were recovered from 7 donors (gender: 6 males, 1 female; age range: 23-50 years) within 24 hours of donor mortality by UMTB/Vivex (Miami, FL) and Articular Engineering (Northbrook, IL) according to the legal and ethical requirements (e.g., institutional review board approval and informed written consent). Samples were placed at 4°C and screened using a standard serology panel for HIV, and hepatitis B and C; within another 48 to 72 hours.
Tissue Harvest
Following receipt of the fresh sterile en-bloc knees, 4, 8-mm full-thickness cartilage explants were harvested from 3 anatomical regions: trochlea, lateral condyle, and medial condyle ( Fig. 1A ), using an osteochondral graft harvester according to the manufacturer’s guidelines (Smith & Nephew, Andover, MA; a total of 12 plugs per donor knee). Within each region, 2 plugs were assigned to the baseline group and the other 2 plugs to the impact group.
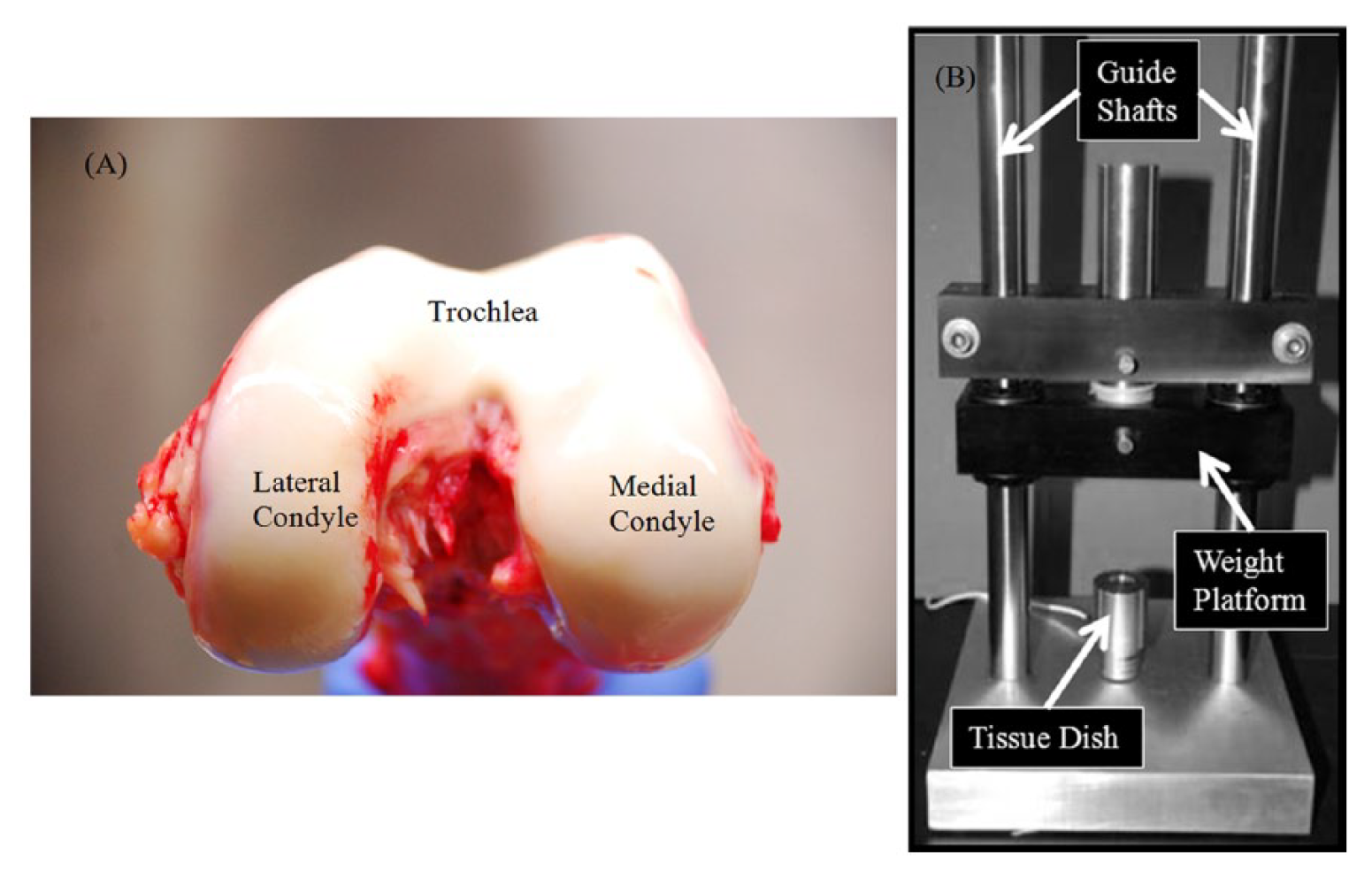
(
Next, the osseous end (bony side) of the plugs were trimmed to a height of 4 mm (full height of the osteochondral section—including cartilage and subchondral bone) using an oscillating autopsy saw with a half-inch blade. The remaining subchondral bone was smoothed and leveled so that the osseous end was parallel to the articular surface. The plugs were soaked in a 1% antibiotic saline solution for 2 hours, then rinsed in phosphate buffered saline (PBS) and fully submerged in culture medium: Dulbecco’s modified Eagle medium (DMEM, Invitrogen Corp, Eugene, OR) with 10% fetal bovine serum (FBS, Atlanta Biological, Lawrenceville, GA), and 1% antibiotic-antimycotic (Atlanta Biological). The plugs were cultured individually in a 24-well plate in 2 mL of media for 72 hours to habituate the tissue to the new environment (37°C with 5% CO2). The medium was changed daily.
Impact Injury
The impact group samples were impacted using a drop tower system ( Fig. 1B ). Pilot testing was performed to determine the optimal weight and height for creating partial thickness articular cartilage lesions, using 20 to 25 MPa of impact pressure (with variance due to individual plug variability). Individual cartilage plugs were placed, cartilage side up, in the drop tower’s tissue dish. A 500 g weight released from a height of 15 cm was used to create impact injuries of cartilage plugs (24.38 ± 9.04 MPa of impact pressure). Following impaction the cartilage, plugs were returned to the 24-well plates for an additional 24 hours and the media was replaced.
Tissue Preparation
Samples were snap frozen in liquid nitrogen for RNA extraction. Briefly, the underlying bone was dissected away from the cartilage. The cartilage sample was submerged in liquid nitrogen for a full minute. The plugs were stored in a −80°C freezer until beginning RNA extraction.
RNA Extraction
Total RNA was extracted from the cartilage plug samples using QIAzol (QIAGEN, Hilden, Germany)-based method. Briefly, frozen cartilage plugs were pulverized using the Covaris CryoPrep system (Covaris Inc., Woburn, MA) by impacting twice at level 5, according to the manufacturer’s guidelines. The cartilage sample was submerged into liquid nitrogen immediately after each impact to keep the tissue powder frozen at all times. Following pulverization, the frozen cartilage powder was transferred to 3 mL of QIAzol. The samples were vortexed vigorously for 1 minute and kept at room temperature for 4 hours and then kept at −80°C until further processing.
To begin RNA extraction, pulverized samples mixed with QIAzol lysis reagent were thawed completely to room temperature and centrifuged at 5,000 × g for 2 minutes. The supernatant was extracted using the QIAzol-chloroform-isopropanol protocol as instructed by the manufacturer. On completion of extraction, RNA samples were quantified using a Thermo Scientific NanoDrop 8000 spectrophotometer. RNA samples with 260/280 ratios less than 1.7 underwent a second purification using the QIAzol lysis reagent procedure described above. Final RNA extraction was quantified by Qubit RNA assays (Invitrogen Life Technologies) and qualified by Agilent 2100 RNA 6000 Pico Chip (Agilent Technologies). In general, ~1 µg of RNA was extracted from each plug for microarray analysis.
Microarray Analysis
Genome-wide gene expression profiling was done using the Illumina Whole Genome (WG)-DASL (cDNA-mediated Annealing, Selection, Extension, and Ligation) High Throughput (HT) Assay. The microarray processing was performed following manufacturer’s instructions. In brief, RNA was first converted to cDNA through a reverse transcription reaction with biotinylated sequences and universal primer sequences. The mixture was bound to streptavidin conjugated paramagnetic particles (SA-PMPs). Mis-hybridized and non-hybridized oligios were washed away. The hybridized oligios were extended and ligated to generate the template for a polymerase chain reaction (PCR) containing a fluorescently labeled primer. The fluorescent products were then hybridized to a BeadChip with >29,000 probes for whole genome profiling. After hybridization, signal intensity images were acquired by scanning the arrays on the Illumina BeadArray Reader. A total of 60 microarrays were performed.
Quantitative Reverse Transcription–Polymerase Chain Reaction Analysis
To validate microarray findings, top differentially expressed genes, that is, LEF1 and EMX2, were measured in total RNAs extracted from lateral condyle and trochlear. TaqMan real-time, PCR assays for each gene were carried out in triplicate and normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) expression.
Statistical Data Analysis
The raw expression data (log2 values) were transformed using variance-stabilizing transformation (VST) 38 with the LUMI Bioconductor R package, which takes into account the large number of technical replicates on the Illumina arrays. Normalization was then conducted using the robust spline normalization (RSN) algorithm, which combines features of quantile and loess normalization. Quality control was performed using the lumiQ command and differential gene expression was conducted using the limma R code. Heatmaps were produced using the heatmap.2 R code where the z-scores (scaled value of the normalized intensity scores) are plotted. Gene expression analysis was performed under 3 separate conditions.
The first compared the gene expression profiles of baseline samples (baseline vs. baseline) among the 3 anatomical regions; lateral condyle, trochlear, medial condyle (LC vs. T, MC vs. T, and MC vs. LC). The overlap in these comparisons was also examined.
The second gene expression analysis compared impacted samples (impacted vs. impacted) among the 3 anatomical regions. The third analysis compared baseline samples to impact injured samples within each anatomical region (baseline vs. impacted). T-tests were run to determine the statistical significance of each probe in a given comparison, resulting in an unadjusted P value.
These analyses were conducted using the Lumi Bioconductor software, 39 which normalizes for biological replicates, batch results, and other technical anomalies, 40 and accounts for multiple testing to identify genes that met predetermined false discovery rate (FDR) thresholds. An FDR-adjusted P value <0.05 was considered to be statistically significant. Additional Gene Ontology (GO) and pathway analyses were performed using the MetaCore GeneGo software. For these analyses, an FDR-adjusted P value <0.05 was also utilized.
Results
Baseline Regional Comparisons
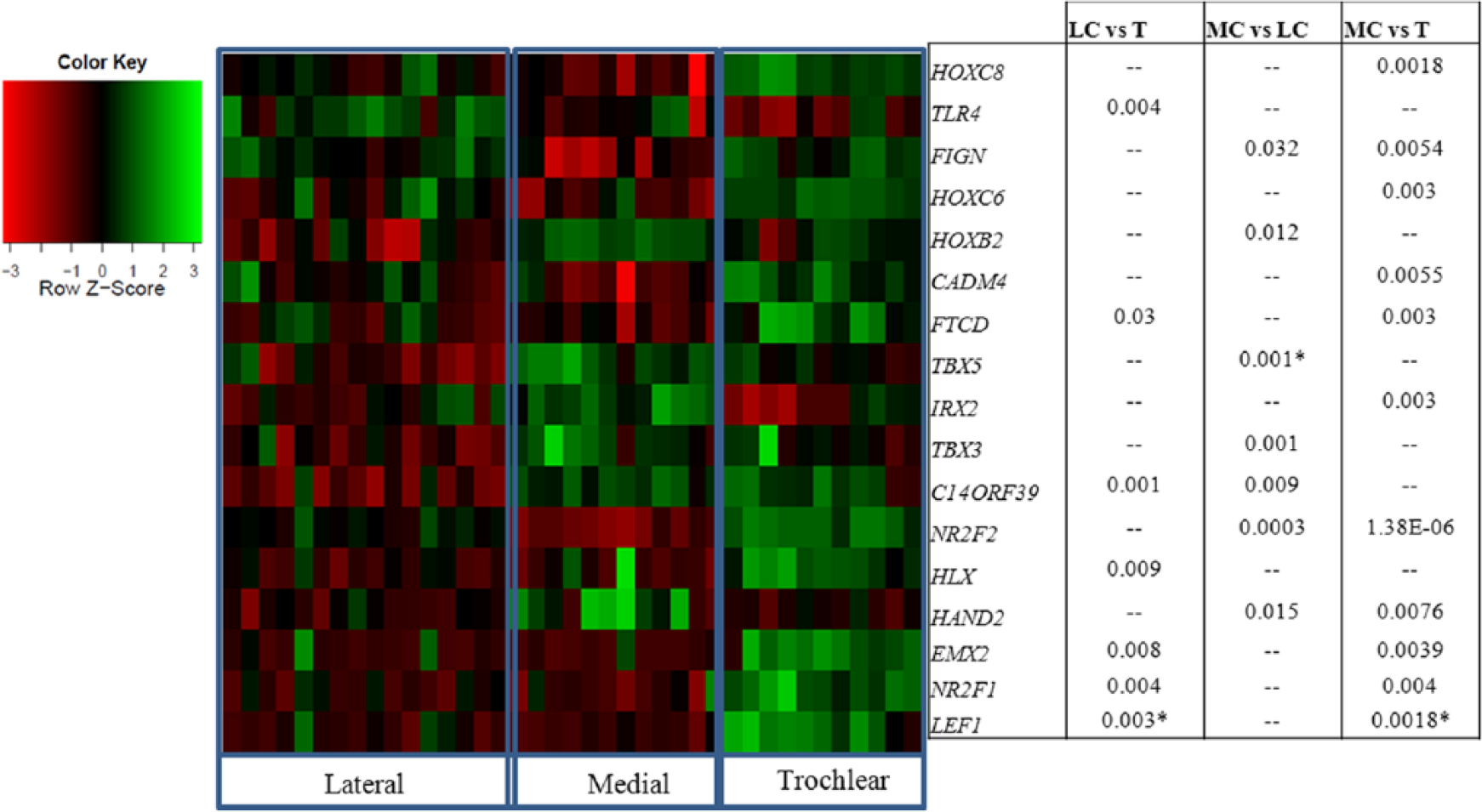
In the comparison of lateral condyle versus trochlea, 18 genes showed significant differential expression (FDR < 0.05) ( Fig. 2 ). Comparison of the medial condyle versus trochlea revealed 21 genes with significant differential expression (FDR < 0.05). In comparing expression in the medial condyle versus lateral condyle, 9 genes (NR2F2, HOXB2, TBX5, TBX3, FIGN, IRX2, IL4R, C14orf39, EBF3) showed significant differential expression (FDR < 0.05) ( Fig. 2 ). Interestingly, 10 genes (EMX2, FTCD, HLX, HOXC11, HOXC6, LEF1, NRF21, PREX2, PRR16, SHOX) overlapped in the medial condyle versus trochlea and lateral condyle versus trochlea comparisons. Specifically, 9 of the genes show significantly higher expression in trochlea than both the medial condyle and lateral condyle (EMX2, FTCD, HLX, HOXC11, HOXC6, LEF1, NRF21, PREX2, SHOX). Only 1 gene, Proline rich 16 (PPR16) showed significantly higher expression in both medial condyle and lateral condyle than in trochlea ( Fig. 2 ). Pathway/GO analyses of the genes from Figure 2 showed enrichment in WNT signaling (JNK(MAPK8-10), Tcf(Lef), Lef-1) and immune response (TLR4, JNK(MAPK8-10)) pathways, and the most significant (FDR P = 1.57 × 10−13) GO process was “0007389- Pattern Specification,” which corresponds to a developmental process that results in the creation of defined areas or spaces within an organism to which cells respond and eventually are instructed to differentiate.

Baseline gene expression analysis. Heatmap of false discovery rate (FDR) significant genes and a list of significantly differentially expressed genes in the baseline samples: trochlea (T), lateral condyle (LC), medial condyle (MC); z-scores (scaled value of the normalized intensity scores [red, decreased intensity; green, increased intensity]; P values, * represents samples with multiple FDR significant probes for the same gene).
Impacted Regional Comparisons
Seven genes were differentially expressed between impacted lateral condyle versus impacted trochlea cartilage (FDR < 0.05) ( Fig. 3 ). Similar analysis of impacted medial condyle versus impacted trochlea cartilage showed that 11 genes were differentially expressed (FDR < 0.05). Four genes (EMX2, FTCD, LEF1, NR2F1) overlapped in these 2 comparisons. These are the same 4 genes that overlapped in the baseline comparison between these same regions (medial condyle vs. trochlea, lateral condyle vs. trochlea) ( Fig. 3 ). Gene expression evaluation of impacted lateral condyle versus medial condyle cartilage showed 7 genes were significant differentially expressed (FDR < 0.05; Fig. 3 ). Of these, 6 genes are also differentially expressed in the baseline comparison ( Fig. 2 ). To validate the microarray finding, Taqman gene expression analysis on EMX2 and LEF1 in lateral condyle and trochlea samples were performed. Both EMX2 and LEF1 showed significantly higher expression in trochlea than in lateral condyle (P < 0.05; Fig. 4 ). Interestingly, FDR-significant genes in each independent comparison (lateral condyle vs. trochlea, medial condyle vs. lateral condyle, and medial condyle vs. trochlea) fall within the same GO process (0007389- Pattern Specification). Overrepresentation in this GO process met FDR significance at P < 10−6) regardless of comparison. With regard to pathways, Lef-1 and Tcf (Lef) were the only 2 genes that were significantly enriched for in the WNT signaling pathway in lateral condyle vs. trochlea (FDR P = 4.46 × 10−4) as well as medial condyle versus trochlea for (FDR P = 1.1 × 10−3) analyses. There were no significantly enriched pathways for the medial condyle versus lateral condyle analysis.
Impacted gene expression analysis. Heatmap of false discovery rate (FDR) significant genes and a list of significantly differentially expressed genes in the impacted samples: trochlea (T), lateral condyle (LC), medial condyle (MC); z-scores (scaled value of the normalized intensity scores [red, decreased intensity; green, increased intensity]; P values, *represents samples with multiple FDR significant probes for the same gene).
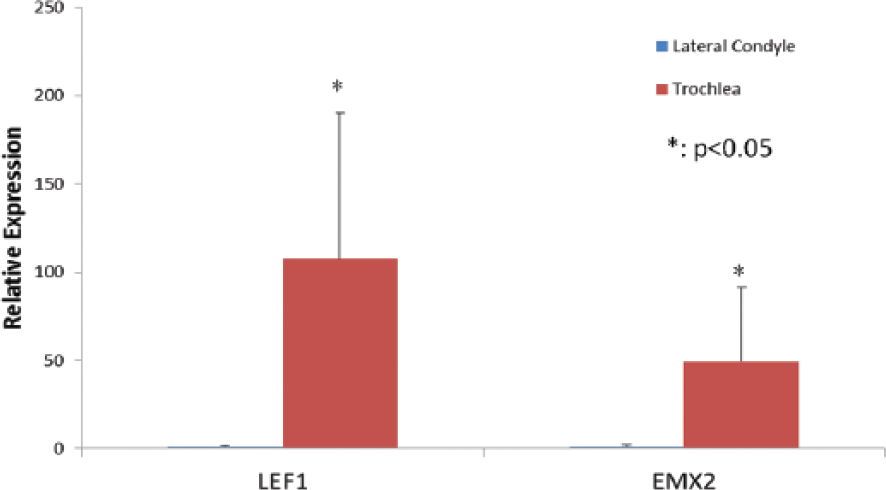
Quantitative reverse transcription–polymerase chain reaction (RT-PCR) analysis of gene expression of LEF1 and EMX2 for lateral condyle and troclea (n = 7).
Injury Response Within Each Region
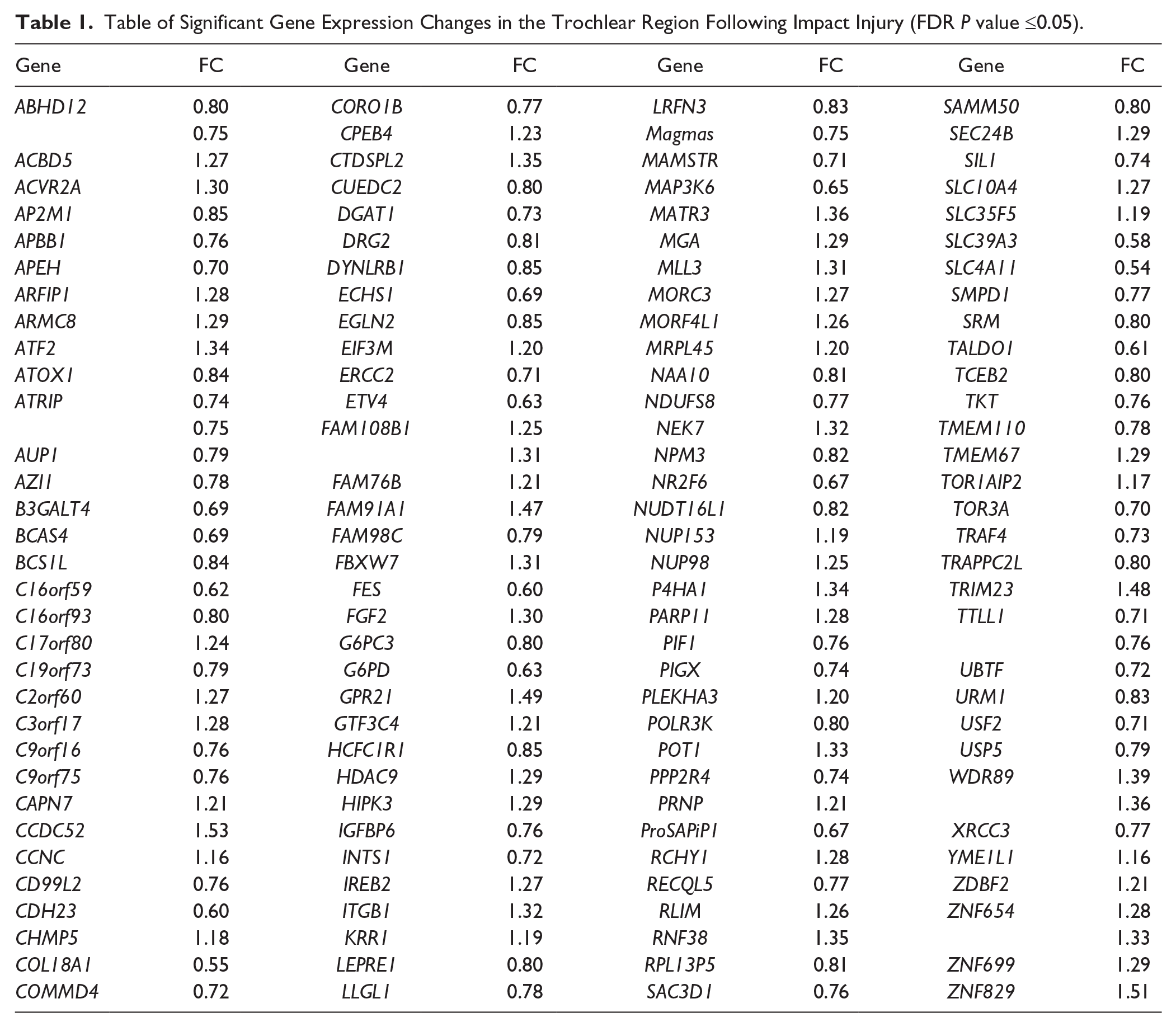
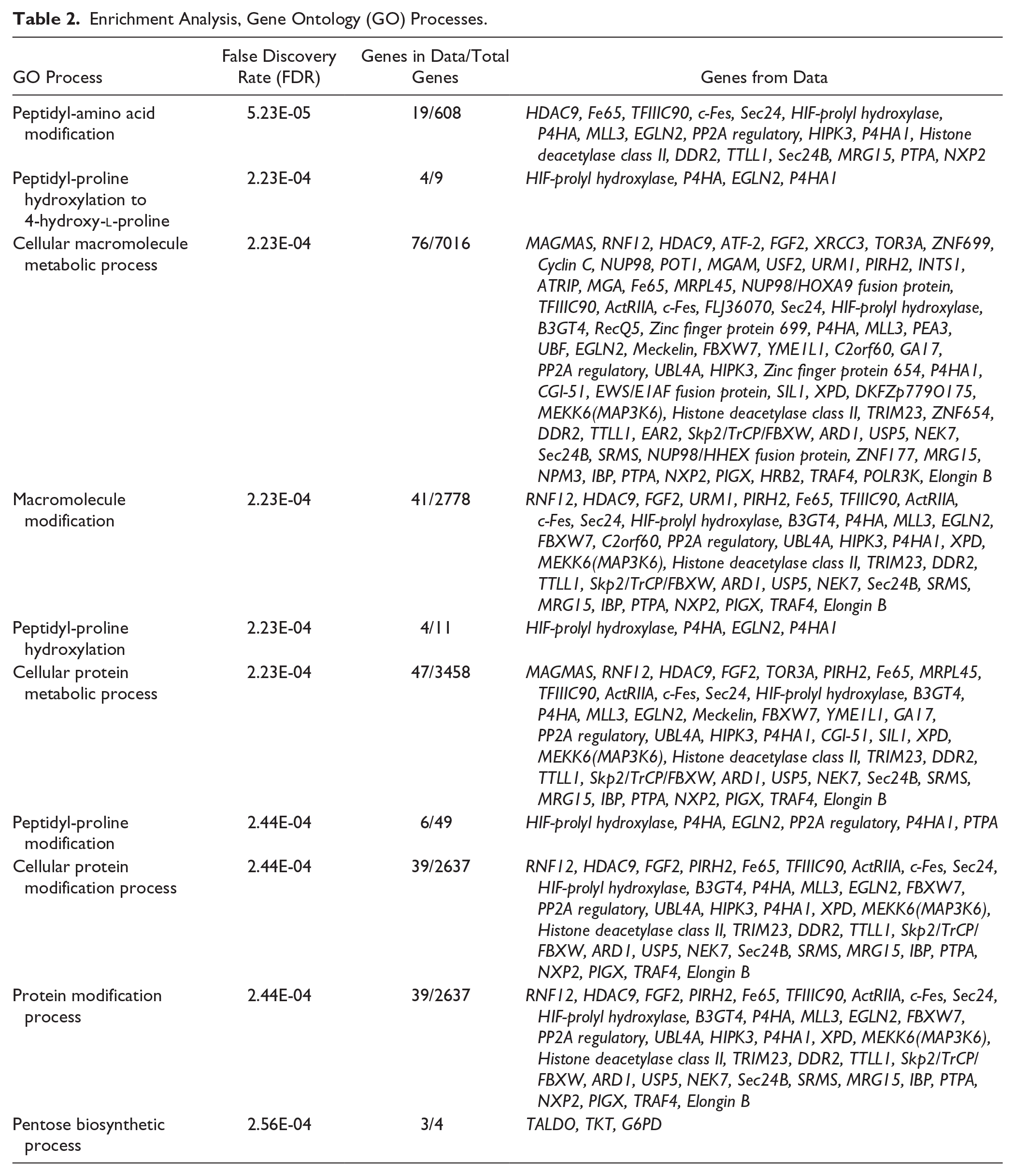
In analyzing baseline versus impacted samples by anatomical regions 130 genes were found to be significantly differentially expressed in the trochlea ( Table 1 ; FDR ≤ 0.05). No statistically significant changes in gene expression were observed within the condyle regions after injury. Significant genes mapped to GO processes identified through Metacore enrichment analysis shown in Table 2 . Pathway analyses revealed only one pathway (pentose phosphate pathway) that met FDR significance, and was enriched for 3 genes (Taldo, TKT, and G6PD). Furthermore, genes that were previously reported to be affected following impact injury and were replicated by our study are shown in Table 3 (unadjusted P value ≤0.05).
Table of Significant Gene Expression Changes in the Trochlear Region Following Impact Injury (FDR P value ≤0.05).
Enrichment Analysis, Gene Ontology (GO) Processes.
Previously Reported Gene Expressions Affected by Impact Injury That Were Replicated by Our Study (Unadjusted P Value ≤0.05).

Note. ↑, significant upregulation; ↓, significant downregulation.
False discovery rate (FDR) P value ≤0.05.
Discussion
The objective of this study was to investigate regional differences in gene expression in human articular cartilage before and after impact injury. Based on observed gene expression patterns, the lateral and medial condyle regions were more similar compared with the trochlear region, both at baseline and following impact injury. A possible explanation for these observations is that lateral condyle and medial condyle receive similar high weight bearing loads during normal daily life activities, whereas the trochlea receives lower mechanical conditioning. When examining differential baseline gene expression, 10 genes are differentially expressed in both the lateral and medial condyle when compared with the trochlea ( Fig. 2 ). Of the 10 genes, H2.0-like homeobox (HLX), homeobox C11 (HOXC11), HOXC6 and short stature homeobox (SHOX) are belong to the family of homeobox genes, which regulate cell differentiation and embryonic limb development,43 -45 suggesting that the structural differences between the trochlea and condyles are dictated by the gene expression profile. Interestingly, several of the genes that were differentially expressed in the baseline regional comparison overlap with the differences seen in the impact comparisons ( Fig. 3 ; LEF1, EMX2, NR2F1, FTCD), suggesting that these anatomical differences are preserved following injury. Previous studies have reported differences in other biological properties within different weight bearing regions. Stenhamre et al. 19 reported that chondrocytes harvested from different weight bearing regions (femur vs. tibia) had varying ability to attach and proliferate in culture. Furthermore, Stenhamre et al. 19 indicated that chondrocytes harvested from weightbearing regions are more differentiated (show less progenitor characteristics) than non-weightbearing regions. Also, Quinn et al. 33 reported differences in material properties, including thickness and proteoglycan content in different weight bearing regions of the knee. Our results support these findings, by demonstrating that regional differences in gene expression are also present.
Our study shows that in addition to baseline differences in gene expression among anatomical regions of articular cartilage, the injury response also varies by region at the molecular level. The trochlear region is the only region that showed FDR significant changes in gene expression (130 genes) after impaction, including genes in the Wnt pathway, suggesting that the trochlear region may be more responsive to injury. Previous studies have reported the significant role of the Wnt pathway in embryonic skeletal system development,46,47 as well as homeostasis of adult joint tissues.48,49 Moreover, Wnt signaling pathways influence chondrogenesis by inducing cytoskeleton reorganization and effecting cell-cell and cell-ECM adhesion.50-53 Therefore, increasing mechanical demand on the trochlear region may alter cartilage properties.
Elevation of pro-inflammatory mediators (i.e., interleukin and tumor necrosis factors) and catabolic enzymes (i.e., matrix metalloproteinase and aggrecanase), which is one of the main characteristics of OA is often found in articular cartilage after subjected to impact force in previous studies.22,23,25,26,28 However, this study did not find such responses in human knee articular cartilage 24 hours after impact. The following are the possible reasons for this difference. First, previous studies used articular cartilage from animals or elderly human donors.22,25,28 Differences in species and age between our and previous studies may result in different cartilage response to injury. Second, the impact stress (20-25 MPa) used in this study has been shown to induce cartilage injury in our previous and other animal studies.25,26,54 Since the loading condition and history of human articular cartilage are different from those of animals, different mechanical preconditioning may result in different genetic responses to injury as shown in this study. Third, previous studies have shown a time-dependent profile of gene expression in articular cartilage after subjected to impact force.25,28 Since gene expression of human articular cartilage was only examined at 24 hours after impact in this study, different findings between this and previous studies could be because human knee articular cartilage has different time-dependent genetic responses compared with articular cartilage obtained from animals or different anatomical locations of human body.22,23,25,26,28 Taken together, the time-dependent response of human knee articular cartilage to different magnitudes of impact stress needs to be further examined in order to better understand the mechanism of impact injury of human knee articular cartilage.
Articular cartilage injuries due to mechanical insults often restrict athletic participation while predisposing the athlete to progressive joint degeneration. 55 Repair of articular cartilage injuries are crucial to reduce pain, improve functionality and ultimately halt further disease progression; allowing athletes to return to preinjury performance levels. Current surgical repair using osteochondral autograft/allograft transplantations (OATS) requires that cartilage is harvested from a non-weightbearing area (e.g., trochlear region) and transplanted into the cartilage lesion or defect, often a high weightbearing region. 56 The differences observed in our study may or may not be helpful in deciding what anatomical site to harvest the cartilage from for an OATS procedure. Our results indicated the trochlear region may have plasticity to adapt to injury as evidence by its response following impact. In contrast, the condyles are more adapted to high mechanical loading, showing no genetic response after impact. Differences in the genetic and mechanical properties between the harvesting and donor locations of different anatomical regions could potentially affect long-term outcomes of OATS procedures. Furthermore, a previous study found that articular cartilage of hip and knee joints exhibited different epigenetic profiles with the enrichment of developmental genes such as the family of homeobox genes, 57 which supports our findings. Therefore, future studies comparing articular cartilage from other joints with different mechanical environments (e.g., ankle vs. knee) could provide a better understanding of adaptation of articular cartilage to mechanical environments.
Genetic markers are used as predictors of disease for both Mendelian and complex genetic disorders.58-62 As medicine moves toward a more personalized predictive and preventative approach, understanding the underlying genetic risk factors will be critical in clinical care. Knowledge of the genetic factors involved in cartilage health has the potential to predict an individual’s increased risk for injury as well as that individual’s treatment response. These data contribute to the growing body of evidence that supports genetics as a potential metric to evaluate cartilage health and the effects of respective treatments. In conclusion, our results support the potential for genetic information to be incorporated into orthopedic medicine.
The major limitations of this study are the small sample size (7 individuals resulting in 82 samples) and the evaluation of only a single time point after injury. As previously suggested, our study may not have sufficient power to detect small effects between the baseline and injury samples. Furthermore, the use of a single evaluation time point may not have allowed sufficient time to elapse for the genetic changes following injury to be statistically noticeable. Investigation into additional time points with increased biological replication is necessary to elucidate the exact molecular events that occur following an acute traumatic injury. Ascertaining the timeline of genetic response is necessary to develop functional treatments to prevent long-term cartilage damage. In addition, since cartilage sample were cultured for 3 days before analysis, in vitro culture condition may introduce some variability.
In summary, our study shows that baseline differences in regional gene expression exist between the trochlear and condylar regions and the genetic differences affect the injury response within those regions in human knee articular cartilage. These findings suggest that mechanical preconditioning could cause regional genetic changes that would result in regional-specific response of articular cartilage to injury.
Footnotes
Informed Consent
Written informed consent was obtained by UMTB/Vivex (Miami, FL) and Articular Engineering (Northbrook, IL) before the human tissue samples were provided to the study.
Acknowledgments and Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: this study was supported by AOSSM/Genzyme Osteoarthritis Basic Science Research Grant.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical Approval
Ethical approval for harvesting human tissue samples for this study was obtained by UMTB/Vivex (Miami, FL) and Articular Engineering (Northbrook, IL).