
Abstract
Objective:
To identify factors that are responsible for the phenotypic differences between transient chondrocytes within human osteophytes prone to endochondral ossification and permanent chondrocytes within articular cartilage persisting for decades.
Methods:
Differential gene expression of chondrocytes from human osteophytes or from articular cartilage was detected by cDNA microarray analysis. The expression of pigment epithelium-derived factor (PEDF), one of the most impressively differentially expressed genes, was validated by quantitative reverse transcriptase polymerase chain reaction as well as immunohistochemistry. The mode of action of PEDF was explored by cell viability assays and by detecting target genes.
Results:
PEDF mRNA expression was upregulated by 118.5-fold (P = 0.01) in human osteophytic cartilage compared with articular cartilage, which was reflected by strong immunostaining for PEDF in the cartilaginous layer of osteophytes but largely negative staining in articular cartilage. Elevated levels of PEDF in osteophytes were associated with enhanced apoptosis. PEDF increased the expression of the proapoptotic factor FasL and induced cell death in cell culture. Osteochondral progenitor cells were more responsive to PEDF than differentiated articular chondrocytes.
Conclusions:
The induction of the proapoptotic factor PEDF within the osteophyte cartilage suggests a molecular concept for the transient chondrocyte phenotype that arises from progenitor cells and is prone to terminal differentiation and cell death.
Introduction
Osteophytes are cartilaginous-osseous outgrowths that may be regarded as an attempt for regeneration in osteoarthritic joints. However, the cells within the cartilaginous layer of human growing osteophytes do not retain a permanent stable chondrocyte phenotype but are finally prone to terminal differentiation and endochondral ossification. 1 The factors that are associated with this transient phenotype, instead of a stable chondrocyte phenotype, have not yet been clarified. Corresponding to the growth plate, terminal chondrocyte differentiation is closely associated with cell death, which suggests a role of proapoptotic factors.2,3
Pigment epithelium-derived factor (PEDF) has recently been identified as a factor that is significantly upregulated in late-stage osteoarthritic cartilage in which chondrocytes are confronted with terminal differentiation and cell death.4,5 Because PEDF is known to induce cell death of endothelial cells, 6 it may also be responsible for terminal differentiation and cell death in cartilage.
Using cDNA microarray analysis, we found PEDF among the factors with the strongest differential expression and significant higher levels (118.5-fold) in osteophytic cartilage compared with articular cartilage. This study explored if PEDF interferes with the stable chondrocyte phenotype by promoting terminal differentiation or cell death.
Methods
Tissue Samples and Cell Preparation
Articular and osteophytic cartilage probes were obtained as matched pairs from the same knee joints of 15 patients undergoing total knee arthroplasty for osteoarthritis. Macroscopically intact articular cartilage was isolated from the dorsal part of the femoral condyles. Osteophytic cartilage was isolated from the same respective joints from the cartilaginous cap of the osteophytes located at the edges of the femoral condyles. Osteophytes were distinguished from the marginal transition area of the joint surface by the existence of a concave ridge toward the joint surface. To selectively isolate the cartilaginous cap of the osteophytes and to exclude any abrasion of bone trabeculae and calcified tissue of the deepest cartilaginous zone, only minimal forces were applied with a scalpel by cutting tangentially to the surface to yield thin consecutive slices of less than 1 mm thickness. Each patient gave informed consent prior to surgery, the institutional ethics committee approved the study and it also has the approval of ethics committee of the University of Erlangen-Nuremberg, Ref. No. 3555.
Microarray
mRNA was isolated from cartilage as described previously. 7 The quality of isolated RNA was assessed using the Agilent 2100 bioanalyzer (Agilent Technologies, Santa Clara, CA), and the concentration was determined with a spectrophotometer (ND 1000; NanoDrop Technologies, Rockland, DE). RNA preparations from a total of 15 different donors were used. Three paired sets of mRNA preparations of articular and osteophytic cartilage each of five donors in equal quantities were used for analysis. Biotin-labeled cRNA was obtained using the two-cycle eukaryotic target labeling assay and hybridized to Affymetrix GeneChip Human Genome U133 Plus 2.0 arrays according to the standard protocol (Affymetrix Inc., Santa Clara, CA). GeneChip Operating Software was used for quality controls of the assays and included scaling factor (SF) and percentage of genes present (GP). Arrays with SF difference <3 and GP >20% were analyzed further. Raw data from gene expression arrays (CEL-files) were processed in Partek Genomics Suite, version 6.5 (Partek Inc., St. Louis, MO) using GCRMA algorithm normalization, PCA (principle components analysis) without outliers, and analysis of variance (ANOVA) yielding P-values and fold change values.
Microarray data presented in this article can be retrieved from the NIH Gene Expression Omnibus (GEO; National Center for Biotechnology Information [NCBI], GSE43923).
Cell Culture Experiments
Articular chondrocytes (AC) and osteophytic chondrocytes (OC) were isolated from intact knee articular cartilage or the cartilaginous layer from osteophytes from five different donors as described previously. 8 As a progenitor cell population that potentially gives rise to forming osteophytes, we isolated osteochondral progenitor cells (PC) from epiphyseal bone marrow by flushing resected osteochondral specimen obtained from total knee joint arthroplasty. Suspended cells were purified from debris with a sterile nylon filter and resuspended in full Dulbecco’s modified Eagle’s medium (DMEM; Gibco, Grand Island, NY) with 10% fetal calf serum (FCS). The plastic adherent cell population was used for the experiments.
After the first passage, cells were plated at semiconfluence, cultivated in DMEM, supplemented with 1% FCS. Recombinant PEDF (R&D Systems, Minneapolis, MN) was added in concentrations of 10, 100, and 200 ng/mL. After stimulation for 6 hours, total RNA was isolated as described previously. 8 For evaluation of the cell viability, cells were stimulated for a total of 6 days, whereas medium and PEDF were refreshed after 3 days. After 6 days, dead cells were detected by Trypan blue staining (Invitrogen, Darmstadt, Germany).
Quantitative RT-PCR
Gene expression of PEDF, Fas ligand (FasL), alkaline phosphatase (ALP), osteocalcin (BGLAP), type 10 collagen (Col10a1), and matrix metalloproteinase-13 (MMP13) was analyzed by quantitative reverse transcriptase polymerase chain reaction (qRT-PCR) using an ABI Prism 7900 sequence detection system (Applied Biosystems, Foster City, CA) and Verso One-Step qRT-PCR Rox Kit (Abgene, Hamburg, Germany). The relative amount of target mRNA was determined and normalized to that of β-2-microglobulin (β2m) mRNA as described previously. 7 Primer and probe sets were purchased by Applied Biosystems (Foster City, CA).
Immunohistological Analysis
Specimens from osteophytes and from macroscopically intact articular cartilage of the same joints were stained with safranin-O/fast green for morphological assessment.
For immunohistochemical detection of PEDF, sections were pretreated with 0.05% trypsin and incubated with goat antihuman PEDF antibodies (Santa Cruz Biotechnology, Santa Cruz, CA), followed by biotinylated secondary antibodies (Dianova, Hamburg, Germany). Bound antibodies were visualized by a complex of streptavidin and biotinylated alkaline phosphatase (Vectastain, ABC-AP, Vector Laboratories, Burlingame, CA). The sections were developed with fast red and counterstained with hematoxylin.
TUNEL Staining
For the detection of in situ DNA breaks, the TUNEL reaction was applied using the In Situ Cell Death Detection Kit, AP (Roche, Mannheim, Germany) as described previously. 9
Statistical Analysis
The microarray data of N = 3 paired preparations were analyzed with Partek Genomics Suite software (St. Louis, MO). Differential gene expression between the two cartilaginous tissues was considered to be significant if the false discovery rates were less than 0.01 and the fold-changes were 20-fold or higher between the different microarrays. Quantitative gene expression of paired individual preparations from N = 15 different donors was analyzed using Student’s t-test, with P < 0.05 considered significant. Cell culture assays were performed in at least three independent experiments and analyzed using Student’s t-test, with P < 0.05 considered significant.
Results
The microarray analysis served as a genome-wide screening for genes with differential expression in osteophytic versus articular cartilage ( Table 1 ). Among the 47,000 screened transcripts, we found PEDF (SERPINF1) among the genes with the greatest differential expression. The validity of the microarray data was confirmed by qRT-PCR with a 118.5-fold enhanced expression of PEDF in the osteophytic cartilage compared with normal articular cartilage (P = 0.01). Putative receptors with affinity for PEDF, PNPLA2, and LRP6 were not differentially expressed.
Differential Gene Expression in Osteophytic Cartilage (“O”) Compared with Articular Cartilage (“A”).
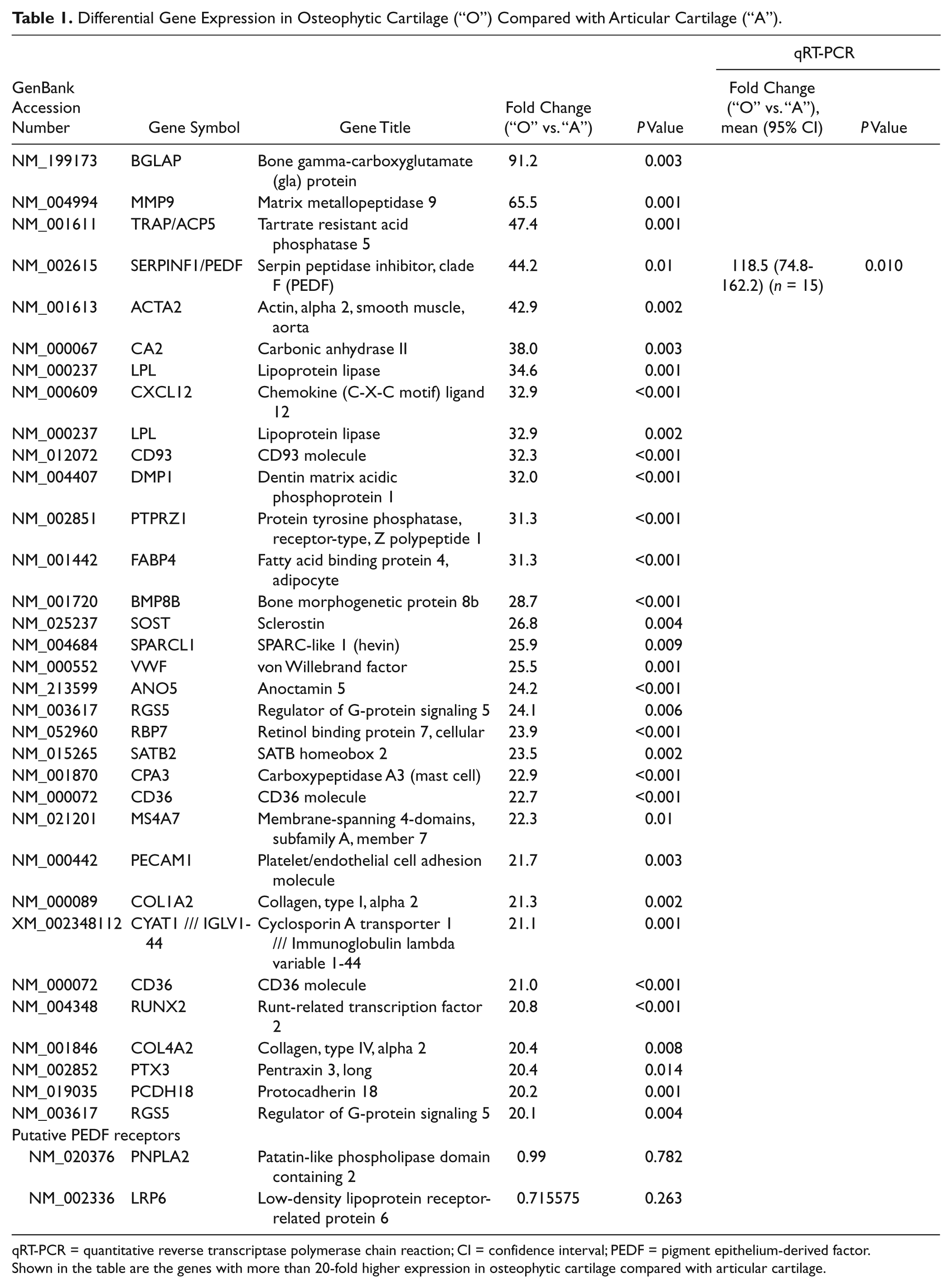
qRT-PCR = quantitative reverse transcriptase polymerase chain reaction; CI = confidence interval; PEDF = pigment epithelium-derived factor.
Shown in the table are the genes with more than 20-fold higher expression in osteophytic cartilage compared with articular cartilage.
Histologically, human osteophytes are characterized by a superficial fibrocartilaginous layer, a zone of hyaline-like cartilage with interspersed hypertrophic chondrocytes in the lower half, followed by bone trabeculae underneath ( Fig. 1a-c ). In contrast, intact articular cartilage lacks hypertrophic chondrocytes ( Fig. 1d ).
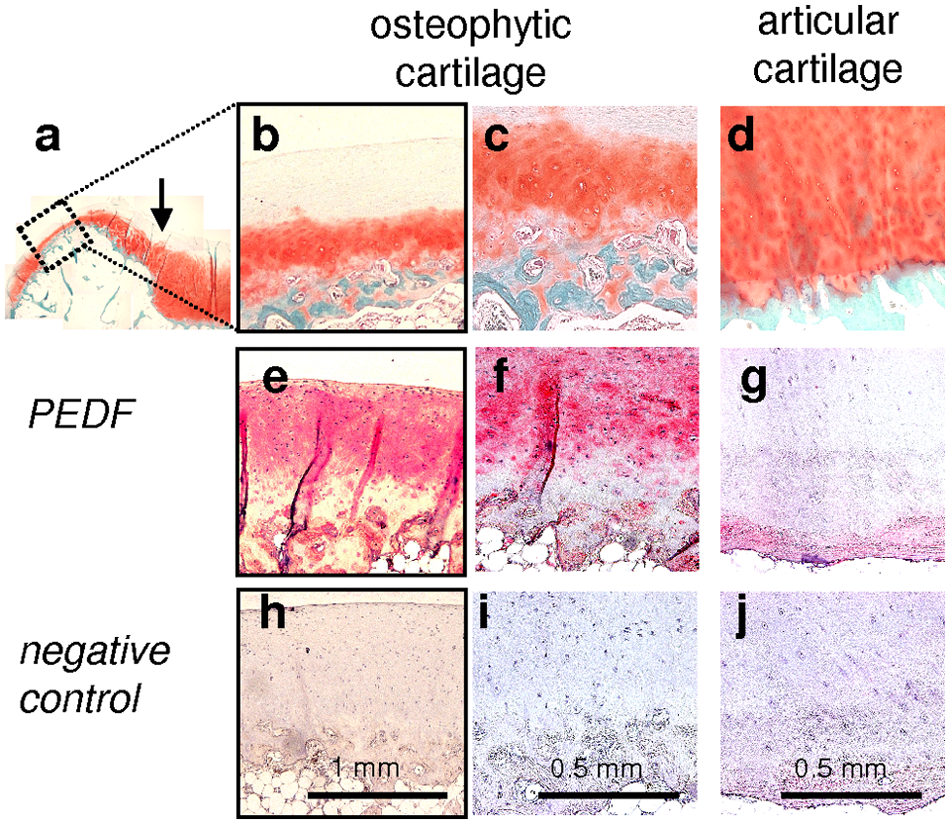
Immunohistochemical detection of PEDF in human osteophytic cartilage and articular cartilage. Safranin-O/fast-green staining depicts an overview of an osteophyte at the joint margin. The arrow indicates the border between articular cartilage and osteophyte (
Immunohistochemistry of osteophytes revealed that PEDF staining was not confined to the deeper zone of hypertrophic cells but present within the extracellular matrix of the whole cartilaginous layer of osteophytes. Except for vessels within the subchondral space, bone trabeculae were negative for PEDF ( Fig. 1e and f ). Intact articular cartilage from the same joints was largely negative for PEDF throughout all layers ( Fig. 1g ).
Cell death within the tissue specimen was assessed by TUNEL staining, which demonstrated a striking number of positively stained (dead) cells in the middle and deeper cartilaginous layer of osteophytes ( Fig. 2b ). Cells of the most superficial fibrocartilaginous layer of osteophytes ( Fig. 2a ) or those of articular cartilage ( Fig. 2c ) stained negative indicating their viable status.
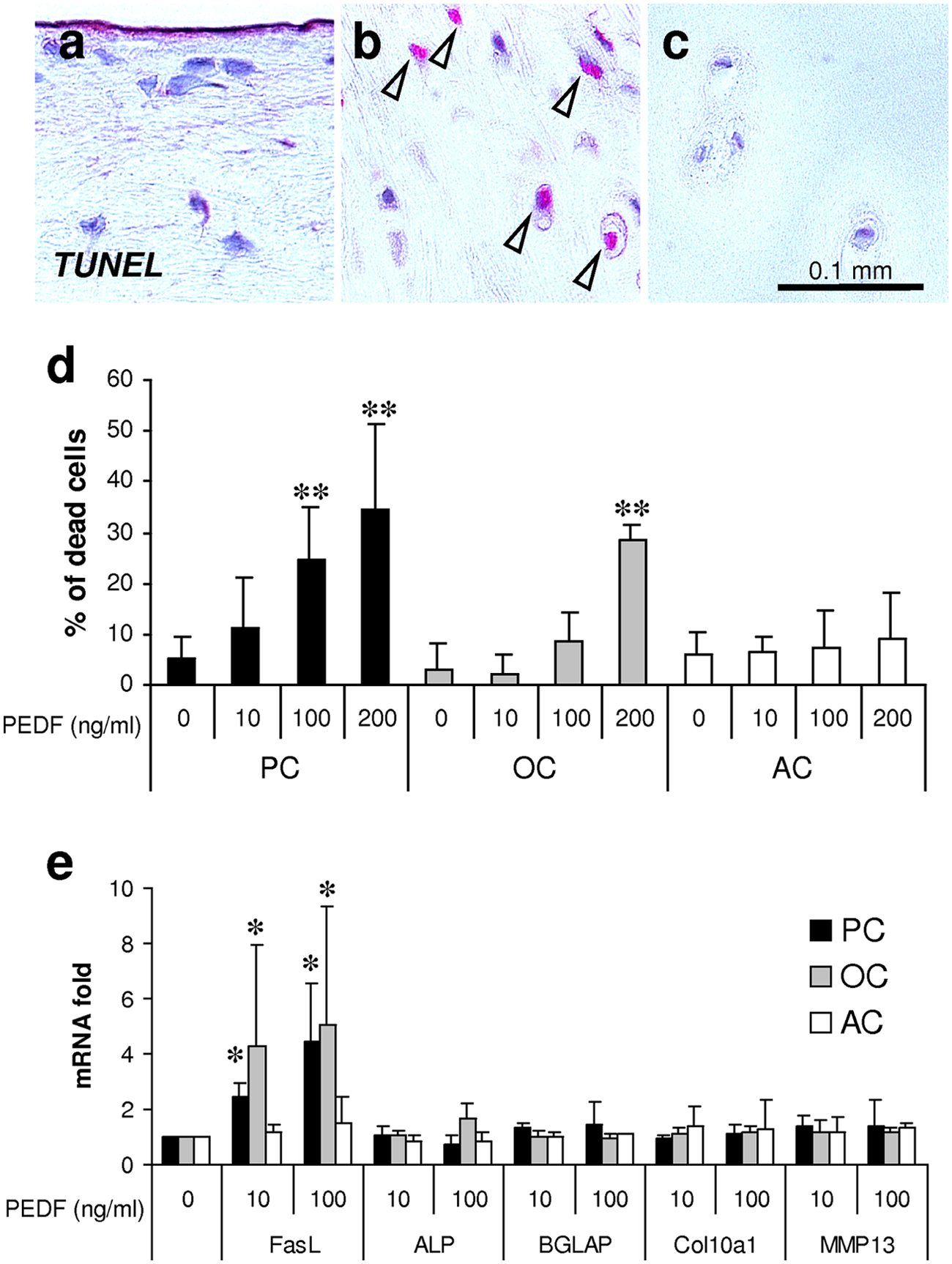
Detection of cell death by the TUNEL method depicts negative staining of cells within the most superficial layer of osteophytes (
The influence of PEDF on cell death was investigated in vitro using freshly isolated AC, OC, and an osteochondral PC population. Stimulation of PEDF induced cell death of OC and PC in a dose-dependent manner. The percentage of dead cells amounted to 34.3% (P = 0.002) for PC or 28.33% (P = 0.01) for OC under the influence of 200 ng/mL PEDF, respectively ( Fig. 2d ). In contrast, the percentage of induced cell death of articular chondrocytes by PEDF did not reach the level of significance ( Fig. 2d ).
To explore the potential mechanism of action of PEDF, we detected its influence on the expression of selected genes involved in apoptosis and terminal chondrocyte differentiation. In OC and PC, PEDF significantly induced the expression of FasL ( Fig. 2e ). In AC, however, the upregulation of FasL by PEDF did not reach the level of significance. PEDF had no influence on the expression of markers for terminal chondrocyte differentiation including ALP, BGLAP, Col10a1, and MMP13.
Discussion
In this study, we demonstrate that PEDF is abundantly expressed by chondrocytes of human osteophytic cartilage. Osteophytes may be regarded as an attempt for cartilage regeneration; however, human osteophytic cartilage does not permanently persist but undergoes the program of endochondral ossification. Interestingly, high levels of PEDF could also be detected in the murine fetal growth plate, a tissue that physiologically relies on the endochondral ossification process with replacement of cartilage by bone during skeletal growth. 10 Thus, we assume that the proapoptotic properties of PEDF may contribute to the transient status of human osteophytic chondrocytes by inducing their terminal differentiation and limited survival. Whereas the morphological patterns and cellular events during skeletal development and joints diseases, including osteophyte formation, may differ to some degree between different species, the action of PEDF may notwithstanding represent a basic biological principle for terminal chondrocyte differentiation that can be observed both within osteophytes in adult organisms and the growth plate cartilage in the fetal or juvenile organism.
In contrast, healthy human articular cartilage is characterized by absence of PEDF activity. Without a doubt, the long-term survival and phenotypic stability of articular chondrocytes involves a plethora of different physical, biochemical, and biomechanical parameters. Thus, absence of PEDF may not be regarded as the primary cause but rather as a relevant prerequisite in this aspect. Indeed, other genome-wide cDNA array analyses support the functional relevance of PEDF, because the stable phenotype of articular chondrocytes was shown to be associated with significant lower PEDF expression than dedifferentiated senescent chondrocytes following prolonged culturing or nondifferentiated mesenchymal stem cells.11,12
PEDF is known as a potent inhibitor of angiogenesis 13 and tumor growth. 14 Its antiangiogenic and antitumorigenic properties can largely be ascribed to its proapoptotic activities. 14 In this study, we could confirm proapoptotic properties of PEDF predominantly on incompletely differentiated chondroprogenitor cells, which supports the rationale for the treatment of malignancies of osteochondral tissues by PEDF.15,16 Importantly, articular cartilage may not be affected by these treatments, because phenotypically stable articular chondrocytes are less responsive to external PEDF stimuli, which may be ascribed to a lower expression of a yet unknown PEDF receptor. The data suggest that healthy articular cartilage is protected from the proapoptotic PEDF signal in two ways, namely, by a lower intrinsic expression and a lower responsiveness. However, the detailed exploration of the PEDF-receptor status in osteochondral tissues will be a task for future studies, because PNPLA2 and LRP6, two receptors with putative PEDF affinity,17,18 were not differentially expressed in the present analysis.
As a conclusion, the induction of PEDF during osteophyte formation provides a molecular concept for the transient phenotype of chondrocyte-like cells arising from mesenchymal progenitor cells that are prone to terminal differentiation and cell death.
Footnotes
Acknowledgements
We thank M. Pflügner and H. Rohrmüller for expert technical support. This study was supported by the German Research Foundation (DFG; Grant GE 1975/2-1; to KG) and by the Interdisciplinary Center of Clinical Research (IZKF; Grant J29; to CB).
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical Approval
This study was approved by the ethics committee of the University of Erlangen-Nuremberg, Ref. No. 3555.