
Abstract
Objective:
ADAMTS (a disintegrin and metalloproteinase with thrombospondin type-1 motif) zinc metalloproteinases are important during the synthesis and breakdown of cartilage extracellular matrix. ADAMTS-12 is up-regulated during in vitro chondrogenesis and embryonic limb development; however, the regulation of ADAMTS-12 expression in cartilage remains unknown. The transcription factor c-Maf is a member of Maf family of basic ZIP (bZIP) transcription factors. Expression of c-Maf is highest in hypertrophic chondrocytes during embryonic development and postnatal growth. We hypothesize that c-Maf and ADAMTS-12 are co-expressed during chondrocyte differentiation and that c-Maf regulates ADAMTS-12 expression during chondrogenesis.
Design:
Promoter analysis and species alignments identified potential c-Maf binding sites in the ADAMTS-12 promoter. c-Maf and ADAMTS-12 co-expression was monitored during chondrogenesis of stem cell pellet cultures. Luciferase expression driven by ADAMTS-12 promoter segments was measured in the presence and absence of c-Maf, and synthetic oligonucleotides were used to confirm specific binding of c-Maf to ADAMTS-12 promoter sequences.
Results:
In vitro chondrogenesis from human mesenchymal stem cells revealed co-expression of ADAMTS-12 and c-Maf during differentiation. Truncation and point mutations of the ADAMTS-12 promoter evaluated in reporter assays localized the response to the proximal 315 bp of the ADAMTS-12 promoter, which contained a predicted c-Maf recognition element (MARE) at position -61. Electorphoretic mobility shift assay confirmed that c-Maf directly interacted with the MARE at position -61.
Conclusions:
These data suggest that c-Maf is involved in chondrocyte differentiation and hypertrophy, at least in part, through the regulation of ADAMTS-12 expression at a newly identified MARE in its proximal promoter.
Introduction
ADAMTS (a disintegrin and metalloproteinase with a thrombospondin type 1 motif) zinc metalloproteinases are active during the synthesis and breakdown of cartilage extracellular matrix. Several ADAMTS metalloproteinases are capable of degrading aggrecan (e.g., ADAMTS-1, -4, -5, -8, -9, -15, -16, and -18), whereas others function as procollagen N-propeptidases (e.g., ADAMTS-2, -3, and -14).1,2 ADAMTS-12 is expressed in synovium, skeletal muscle, cartilage, and fat. 3 ADAMTS-12 and ADAMTS-7 degrade cartilage oligomeric matrix protein (COMP), a major noncollagenous protein component of the cartilage extracellular matrix. 4 COMP is the only known substrate ADAMTS-12. 3 The exact role of ADAMTS-12 in chondrocyte differentiation remains to be elucidated. ADAMTS-12 is up-regulated during in vitro chondrogenesis as well as mouse embryonic limb development with prominent expression in physeal chondrocytes.5,6 Ectopic expression of ADAMTS-12 in mesenchymal stem cells paradoxically inhibited of chondrogenic differentiation. 5 ADAMTS-12 is also implicated in the pathogenesis of osteoarthritis. ADAMTS-12 expression is higher in the cartilage and synovium of patients with osteoarthritis compared with healthy controls.3,7 Given its roles in both chondrogenesis and cartilage degradation, it is important to understand the regulation of ADAMTS-12. Currently, the factors that control ADAMTS-12 expression remain unknown.
The transcription factor c-Maf is a member of Maf family of basic ZIP (bZIP) transcription factors. 8 Maf proteins interact with other proteins through the leucine zipper domain. Maf proteins bind to a specific DNA sequences termed the “Maf recognition element” (MARE) through the basic DNA recognition domain. 9 The downstream targets of the transcription factor c-Maf in chondrocytes include connective tissue growth factor (CTGF), type II collagen (Col2a1), and type 27 collagen (Col27a1). 10 The transcription factor c-Maf plays a role in chondrogenic differentiation and long bone growth.10,11 Expression of c-Maf is highest in hypertrophic chondrocytes during embryonic development and postnatal growth.11,12 Mice with a homozygous deletion of c-Maf have abnormal terminal differentiation of hypertrophic chondrocytes accompanied with a delay in vascular invasion and reduced ossification. 11 Osteoarthritic chondrocytes overexpress c-Maf. 13
The similar expression patterns of both ADAMTS-12 and c-Maf in osteoarthritis and development led to our underlying hypothesis that c-Maf may regulate ADAMTS-12 expression. Specifically, we hypothesized that c-Maf and ADAMTS-12 are co-expressed during hypertrophic chondrocyte differentiation and that c-Maf regulates ADAMTS-12 expression during in vitro chondrogenesis of human mesenchymal stem cells via a MARE responsible for c-Maf dependent ADAMTS-12 promoter activation.
Materials and Methods
Sequence Analysis of the ADAMTS-12 Proximal Promoter Region
The 3 kbp proximal promoter region of ADAMTS-12 was analyzed for putative transcription factor binding sites by the TRANSFAC MatInspector (http://www.genomatix.de/cgi-bin/matinspector/matinspector.pl) and TFSEARCH (http://www.cbrc.jp/research/db/TFSEARCH.html) programs. The transcription start site of ADAMTS-12 was predicted by the McPromoter (http://tools.igsp.duke.edu/generegulation/McPromoterMMII) program.
Gene Expression Analysis during Chondrogenic Differentiation of Human Bone Marrow Stem Cells (hBMSCs) in Three-Dimensional Cell Pellet Culture
Stem cells were isolated from femoral bone marrow aspirates collected from two patients undergoing total knee arthroplasty under an institutional review board–approved protocol with patient consent according to the method described by Pittenger et al. 14 and culture-expanded in MS medium CGM (Lonza, Basel, Switzerland). To form the cell pellets, 5 × 105 passage-2 hBMSCs were centrifuged at 150 rcf for 5 minutes in conical polypropylene centrifuge tubes and maintained in chondrogenic media. Chondrogenic media contained DMEM/F-12 supplemented with 50 µg/mL ascorbate phosphate, 100 µg/mL pyruvate, 1% ITS+ Premix (BD Biosciences, Franklin Lakes, NJ), 1% penicillin/streptomycin, 10 ng/mL recombinant human transforming growth factor-β1 (TGF-β1; PeproTech, Rocky Hill, NJ), and 500 ng/mL recombinant human insulin-like growth factor-1 (IGF-1; R&D Systems, Minneapolis, MN). The cell pellets were cultured for up to 11 days with chondrogenic media changes every 3 to 4 days, with three to four pellets from each donor analyzed per time point. RNA was isolated from cell pellets using RNeasy total RNA kit (Qiagen, Hilden, Germany), quantified on a NanoDrop2000 spectrophotometer (A260 method), and cDNA was synthesized using High-capacity Reverse Transcriptase reagent (Applied Biosystems, Foster City, CA). Real-time quantitative reverse transcription polymerase chain reaction (qRT-PCR) was performed on an ABI-7700 Sequence Detector with Assays-on-Demand TaqMan primers and probes, using universal TaqMan conditions (Applied Biosystems). The primer/probesets were the following: cMaf-Hs02576418 _s1, Col2a1-Hs01060345_m1, Col10a1-Hs00166657_m1, GAPDH-4326317E, and ADAMTS12-Hs00229594_m1. The TaqMan probe for c-Maf is to a region common to both isoforms of c-Maf (Lc-Maf and cMaf) and cannot distinguish between the two. Gene expression was normalized to glyceraldehyde 3-phosphate dehydrogenase (GAPDH) mRNA expression and the fold-change was calculated using the 2−ΔΔCt method.
Cloning of ADAMTS-12 Promoter and c-Maf Constructs
The 3 kbp 5′ upstream region (−2845 to +163) of the human ADAMTS-12 gene was cloned using two sets of nested PCR primers ( Table 1 ) from a human BAC clone template RP11-669P5 (CHORI, Oakland, CA). A series of sequential truncations of the ADAMTS-12 promoter were subcloned into the pGL4.10 luciferase vector (Promega, Madison, WI) via PCR. Specific deletions of 13 nucleotide putative MARE sequences at -2311 (Δdistal) and -61 (Δproximal) were generated in the 3 kbp ADAMTS-12 promoter or 315 bp promoter constructs using QuickChange site-directed mutagenesis (Stratagene, La Jolla, CA). Primer sequences used for deletion mutagenesis are shown in Table 1 .
Synthetic Oligonucleotide Sequences Used for PCR and EMSA
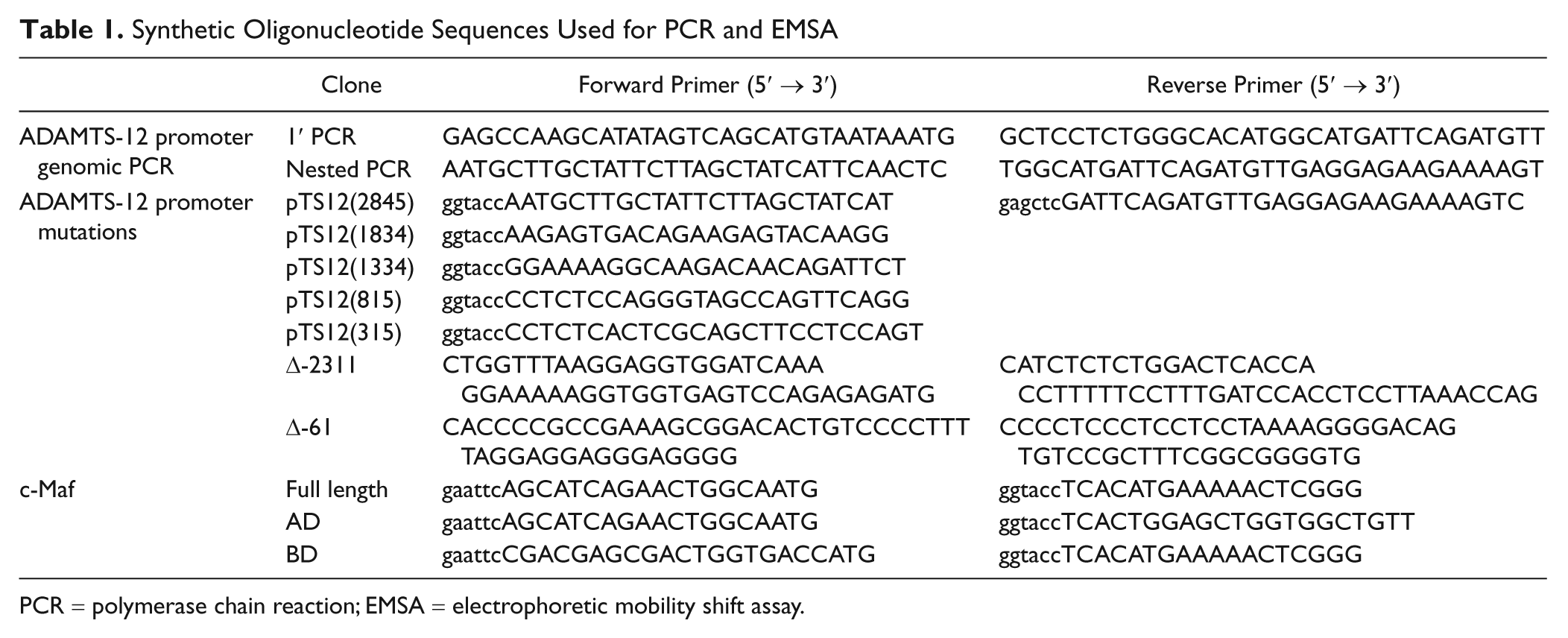
PCR = polymerase chain reaction; EMSA = electrophoretic mobility shift assay.
The coding region of human c-Maf was obtained from the cDNA clone (GenBank Accession: BC081542, Open Biosystems/ThermoFisher, Lafayette, CO) by PCR using gene-specific primers ( Table 1 ). The full-length, transactivation domain (amino acids 1-125) and binding domain (amino acids 252-373) were subcloned into the pFlagCMV2 mammalian expression vector (Sigma-Aldrich, St Louis, MO). Expression was verified by SDS-PAGE on Novex precast 4% to 12% gels, followed by blotting onto nitrocellulose and probing with anti-Flag (Sigma #F1804, 1:5,000) or anti-GAPDH (Ambion #AM4300, 1:15,000). Detection was with horseradish peroxidase–conjugated secondary and luminol substrate exposed to standard x-ray film.
ADAMTS-12 Promoter Activation Assays
SW1353 chondrosarcoma cells (ATCC) were seeded to 1 × 105 cells per well in 12-well plates 24 hours prior to transfection. Triplicate wells of cells were cotransfected with ADAMTS-12 promoter constructs and Renilla luciferase pRL-TK (Promega) (molar ratio = 50:1) with or without c-Maf constructs. The total amount of transfected DNA was kept constant by addition of pUC19 DNA. Luciferase activity was measured 48 hours posttransfection with a GloMax 20/20 luminometer using a Dual-Luciferase Reporter Assay System (Promega). Transfection efficiency was normalized with Renilla luciferase activity. Each data set is from three different experiments, and statistical comparisons were performed using JMP software (SAS, Cary, NC) using two-sided t tests with significance set at P < 0.05.
Binding of c-Maf to MARE by Electrophoretic Mobility Shift Assay (EMSA)
Single-stranded oligonucleotides corresponding to ADAMTS-12 MARE, scrambled, and consensus MARE sequences ( Fig. 5A ) were synthesized (Integrated DNA Technologies, Coralville, IA) and labeled using Biotin 3′ End Labeling Kit (ThermoFisher). Two complimentary biotin-labeled oligonucleotides were annealed to generate double-stranded probes. SW1353 cells were transiently transfected with c-Maf expression plasmid, and nuclear extracts were prepared from using NE-PER Nuclear and Cytoplasmic Extraction Reagents (ThermoFisher). EMSA was performed using the LightShift Chemiluminescent EMSA kit, following the manufacturer’s protocol (ThermoFisher). Briefly, DNA/protein binding reactions were performed using 200 fmol of biotin-labeled probe and 3 µg of nuclear protein extracts in 1× binding buffer containing 50 ng/µL poly dI:dC and 0.05% NP40 at room temperature for 20 minutes. In competition reactions, nuclear extracts were incubated with 100-fold molar excess of unlabeled oligonucleotides before the addition of probe. In supershift assays, nuclear extracts were incubated with 2 µg of anti-c-Maf antibody (Santa Cruz Biotechnology, Santa Cruz, CA) for 15 minutes at room temperature before the addition of probe. Samples were resolved in a 6% DNA retardation gel (Invitrogen Life Technologies, Carlsbad, CA) in 0.5× TBE buffer in a cold room, electro-blotted onto Biodyne B nylon membrane (ThermoFisher), and ultraviolet cross-linked at 120 mJ/cm2 for 1 min. Chemiluminescent detection was performed following the manufacturer’s protocol.
Statistical Analysis
Experiments were done in triplicate and repeated unless otherwise stated. Statistical comparisons were made in Microsoft Excel software using Student’s t test with P < 0.05 considered statistically significant. In all figures, error bars indicate standard deviation and significant comparisons are indicated with an asterisk (*).
Results
The Proximal Promoter Region of ADAMTS-12 Contains Seven Putative Maf Binding Sites
Sequence analysis of the 3 kbp promoter indicated that ADAMTS-12 promoter does not contain a typical TATA or CAAT box. Instead, the proximal promoter has a GC-rich region containing four putative Sp1 binding sites (at -89, -134, -173, and -227) that are often found in TATA-less promoters. Among many predicted transcription factor binding sites in the 3 kbp region, the ones with the highest core similarity to the consensus sequence are Maf, AP1 related factor, nuclear factor κB (NFκB), glucocorticoid receptor, and hypoxia inducible factor (HIF). Seven Maf binding sites were predicted in the proximal 3 kbp ADAMTS-12 promoter ( Fig. 1A ), with varying degrees of similarity to the MARE consensus sequences ( Fig. 1B ).8,9 The predicted Maf binding sites at -61 and -654 positions contain the highest degree of conservation with the MARE consensus sequences in their 5′ half of their palindromic sequences ( Fig. 1B ). A sequence comparison of the human and mouse ADAMTS-12 promoters revealed that the MARE containing region at -61 position is highly conserved, whereas the sequences at -654 position is not, indicating that the predicted Maf binding site at -61 position may be functionally conserved ( Fig. 1C ).
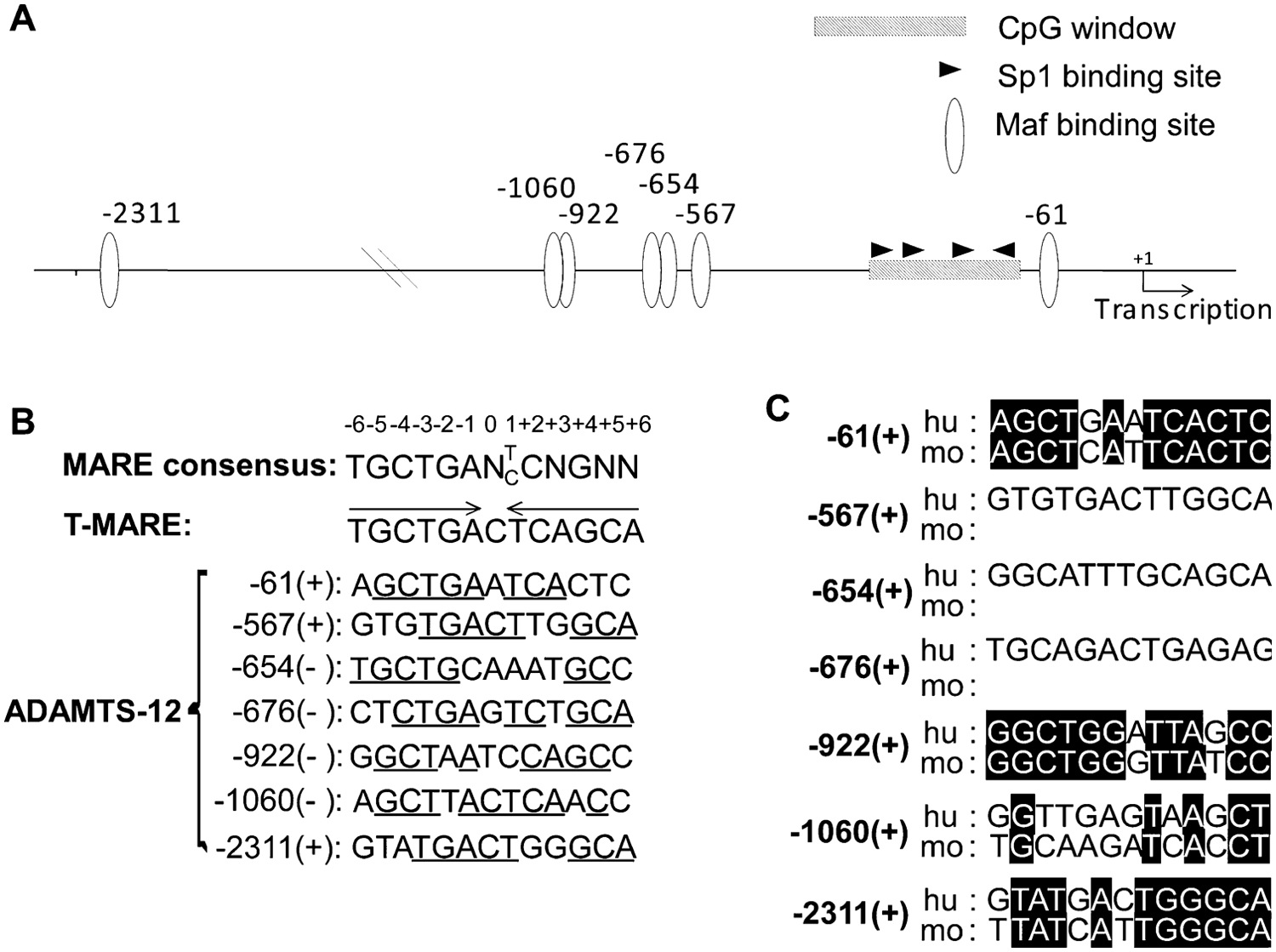
Sequence analysis of the proximal 3-kb promoter region of ADAMTS-12. (
c-Maf and ADAMTS-12 Are Co-expressed During In Vitro Chondrogenic Differentiation
To test whether there is co-expression of c-Maf and ADAMTS-12 during chondrogenic differentiation, hBMSCs were cultured in three-dimensional cell pellets for 11 days in chondrogenic media. ADAMTS-12 and c-Maf mRNA was monitored by real time qRT-PCR ( Fig. 2 ). Differentiation along the chondrogenic pathway was assessed by the up-regulation of a chondrocyte marker, type 2 collagen (Col2a1), and a chondrocyte hypertrophy marker, type X collagen (Col10a1). Both markers increased during this period, indicating evidence of in vitro chondrogenesis. The mRNA expression of c-Maf and ADAMTS-12 increased 12-fold and 11-fold, respectively, during this in vitro chondrogenic differentiation assay.
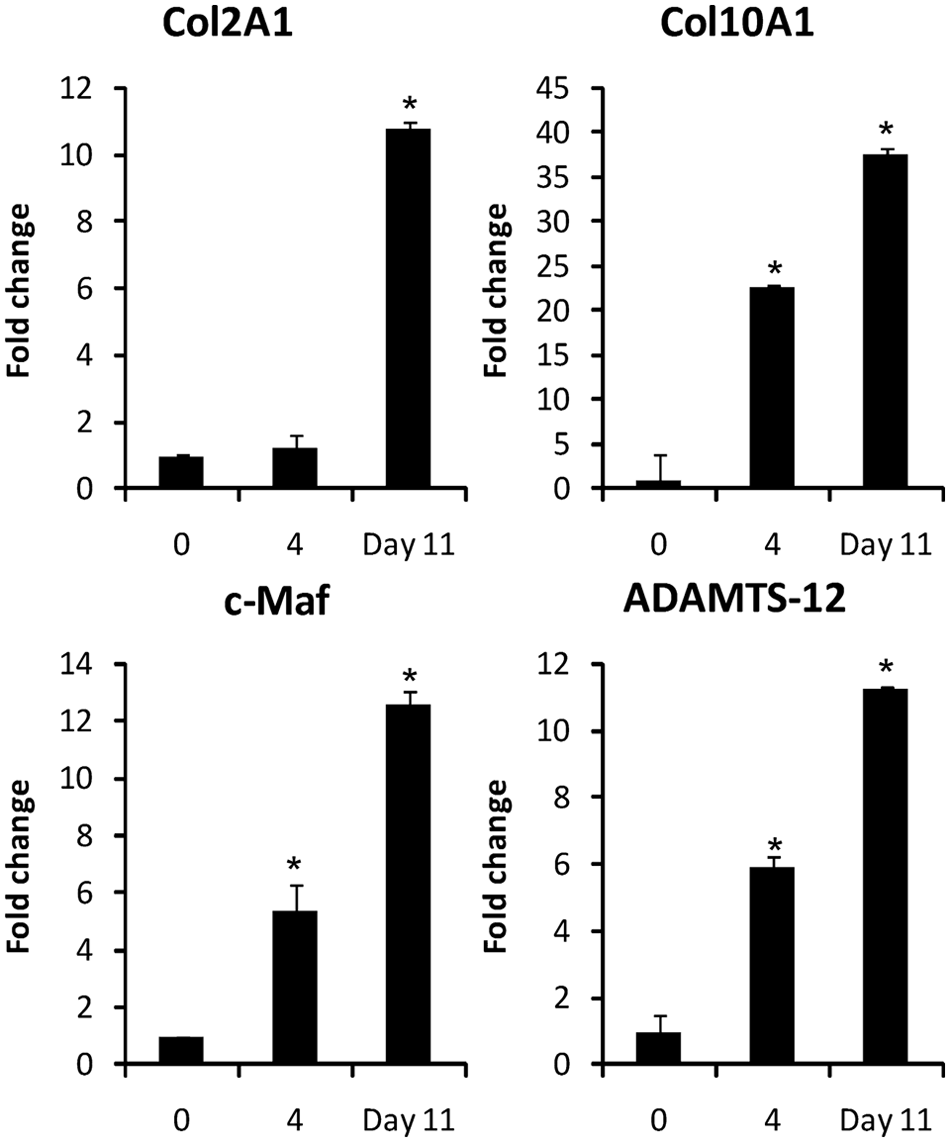
Co-expression of c-Maf and ADAMTS-12 during in vitro chondrogenic differentiation. Endogenous c-Maf and ADAMTS-12 mRNAs were monitored by qRT-PCR during chondrogenic differentiation of hBMSCs in three-dimensional pellet cultures. Col2a1 expression was measured to confirm chondrogenic differentiation, and Col10a1 expression was measured to confirm initiation of chondrocyte hypertrophy. (*) Indicates statistically significant difference in gene expression relative to day 0, with P < 0.05. qRT-PCR = quantitative reverse transcription polymerase chain reaction.
The -315 Region of the ADAMTS-12 Promoter Is Sufficient for Activity in Chondrocytic Cells
We next addressed the transcriptional functionality of the ADAMTS-12 promoter. The ability of the ADAMTS-12 promoter to drive luciferase activity was tested in transient transfection assays in SW1353, 293T human embryonic kidney cells, and C20/A4 immortalized chondrocytes. The 3 kbp promoter directed an increase of luciferase activity in all cell lines tested compared to the basal control vector ( Fig. 3B ). All subsequent reporter assays were performed in SW1353 cells. Sequential 5′ deletions in the promoter were generated, which spanned different numbers of the predicted Maf binding sites ( Fig. 3A ). Analysis of all five different constructs for basal expression showed that the most proximal 315 bp region conferred similar full basal expression ability as 3 kbp promoter. No significant increase of promoter activity was observed in larger constructs. This indicates that the proximal 315 bases are sufficient for the transcriptional basal activity of the human ADAMTS-12 promoter ( Fig. 3C ).
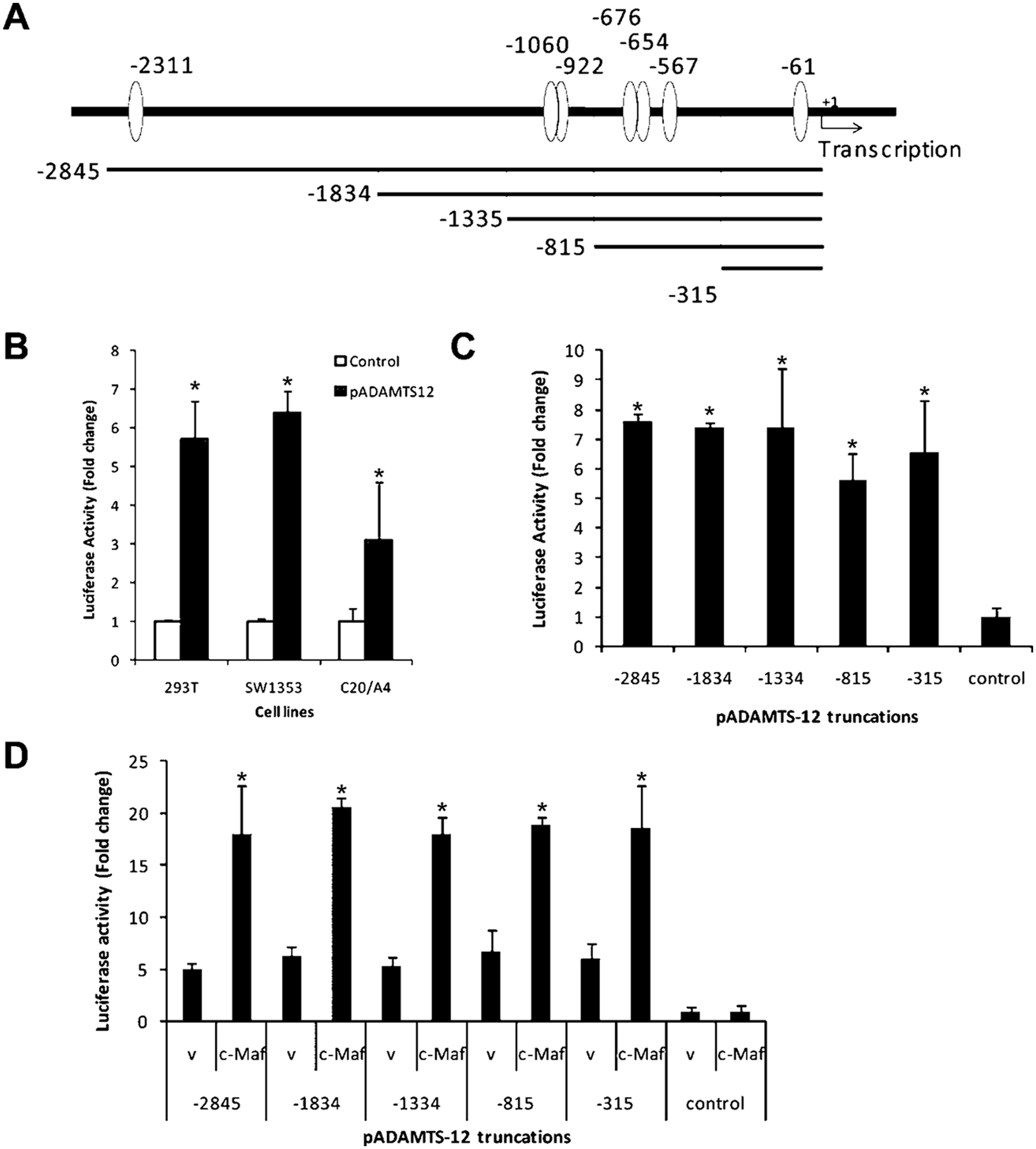
The activity of ADAMTS-12 promoter is up-regulated by c-Maf. (
The Proximal -315 Promoter Region of ADAMTS-12 Contains an Active MARE Site
To determine the relative contributions of the promoter region containing predicted Maf binding sites to c-Maf activation, cotransfection experiments were performed with each sequential 5′ deletion constructs along with c-Maf expression construct. The activity of the 315 bp promoter was up-regulated approximately threefold by c-Maf. Additional upstream promoter sequences conferred no additional activation by c-Maf. This demonstrates that the proximal 315 bp region of the ADAMTS-12 promoter was sufficient to direct c-Maf-specific transactivation ( Fig. 3D ).
To test which predicted c-Maf binding sites are necessary for transactivation by c-Maf, we quantified the activities of the promoters with specific deletions of the putative binding sites. The Δproximal deletion abrogated c-Maf-induced transactivation, whereas Δdistal did not affect c-Maf-induced transactivation ( Fig. 4A ). Furthermore, the Δproximal (-315) also abrogated c-Maf-induced transactivation of ADAMTS-12 (-315) ( Fig. 4B , compare bars 2 and 8 labeled cMaf). Thus, we conclude that the putative MARE at the -61 position is active in ADAMTS-12 promoter.
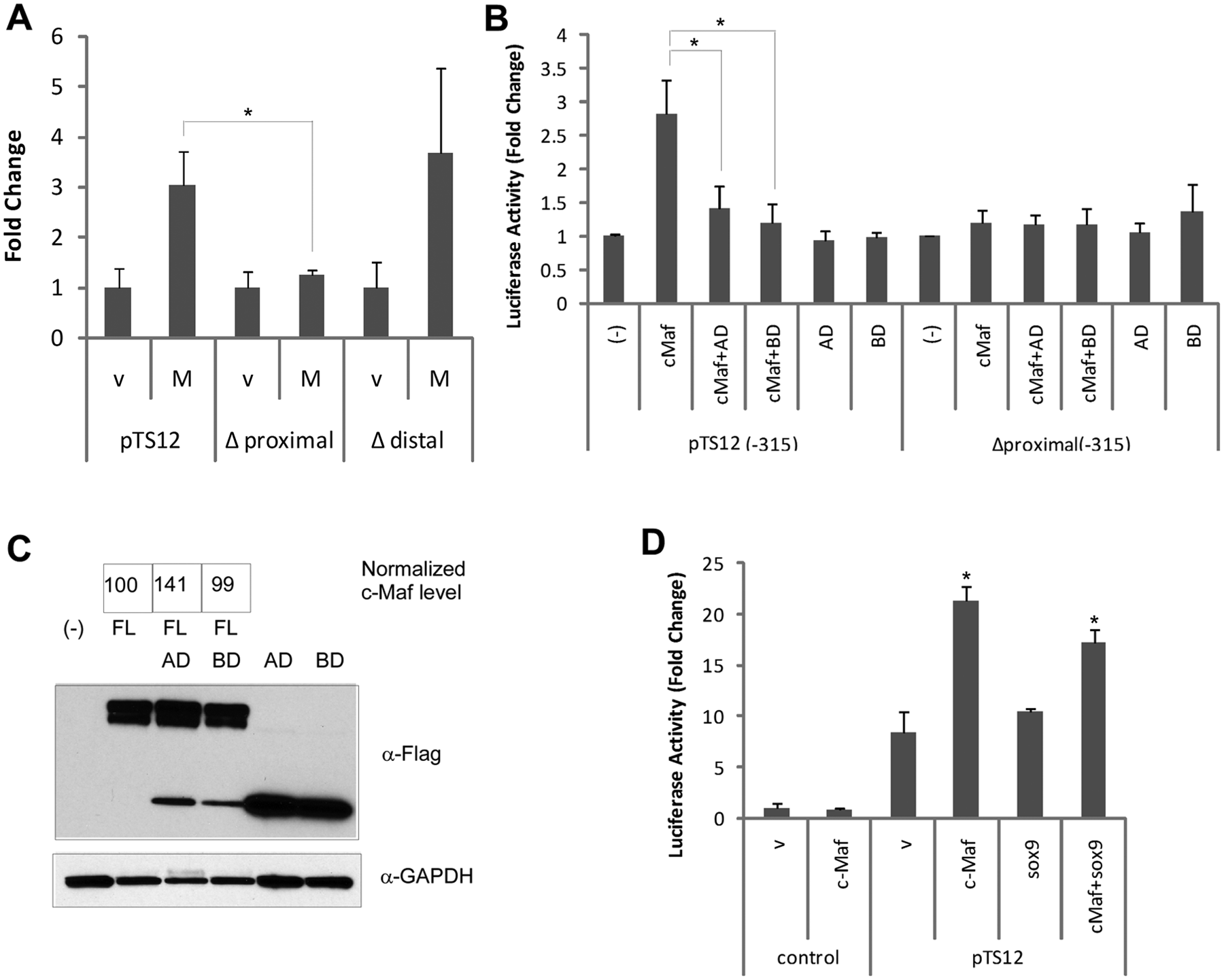
Functional c-Maf-response element was identified in the ADAMTS-12 promoter. (
Specificity of c-Maf in Transactivation of the ADAMTS-12 Promoter
A functional c-Maf consists of an N-terminal transactivation domain and C-terminal bZIP binding domain. To further establish the specificity of the interaction between c-Maf and the ADAMTS-12 promoter, we made two constructs of c-Maf, one containing only the binding domain (BD) that can act as a dominant-negative mutant and one construct containing only the activation domain (AD). Neither BD nor AD alone effected ADAMTS-12 promoter activity ( Fig. 4B , bars 5 and 6), suggesting that both domains are required for ADAMTS-12 promoter activation. BD abrogated full-length (FL) c-Maf-induced transactivation to the empty vector control level, probably because of its dominant negative effect ( Fig. 4B , bars 1, 2, and 4). AD also repressed FL c-Maf-induced transactivation, possibly by competing for binding to other factors ( Fig. 4B , bar 3). The decrease of the transactivation by c-Maf truncations is not due to the change of FL c-Maf protein level, because the values of the normalized c-Maf level do not reflect the change in the reporter assay ( Fig. 4C ). Neither BD nor AD effected Δproximal (-315) promoter activity ( Fig. 4B , lanes 9-12). In addition, we tested for an interaction with the Sox9 transcription factor and found that Sox9 itself did not confer any transactivation on ADAMTS-12 promoter, either alone or in combination with c-Maf ( Fig. 4D ).
c-MAF Directly Binds to the Proximal MARE Sequences of ADAMTS-12 In Vitro
To examine whether c-Maf directly binds to the proximal MARE sequences in ADAMTS-12 promoter in vitro, we performed EMSA using the proximal MARE sequences as a probe and the nuclear extract from c-Maf-transfected SW1353 cells. The 23 bp MARE probe (position from -66 through -44) contains 13 nucleotides of predicted MARE sequences and 5 extra nucleotides at each end ( Fig. 5A ). As a negative control, the sequences were scrambled so that no statistically significant putative transcription factor binding sites were detected in these scrambled sequences using TFSEARCH program (data not shown). The consensus MARE sequences (T-MARE) were used as a positive control. EMSA with proximal MARE probe revealed two shifted bands (bands 1 and 2). Excess of nonlabeled probe disturbed the specific shifted band (band 1), but not the nonspecific band (band 2) ( Fig. 5 , lanes 1-3). Scrambled negative control probe did not generate specific shifted band ( Fig. 5 , lanes 5 and 6). We observed disappearance of band 1 when anti-c-Maf antibody was added to the binding reaction ( Fig. 5 , lane 4), which may occur through disruption of the protein–DNA binding. These results confirm that a specific binding interaction occurs between c-Maf and the MARE sequence at -61 of the human ADAMTS-12 promoter.
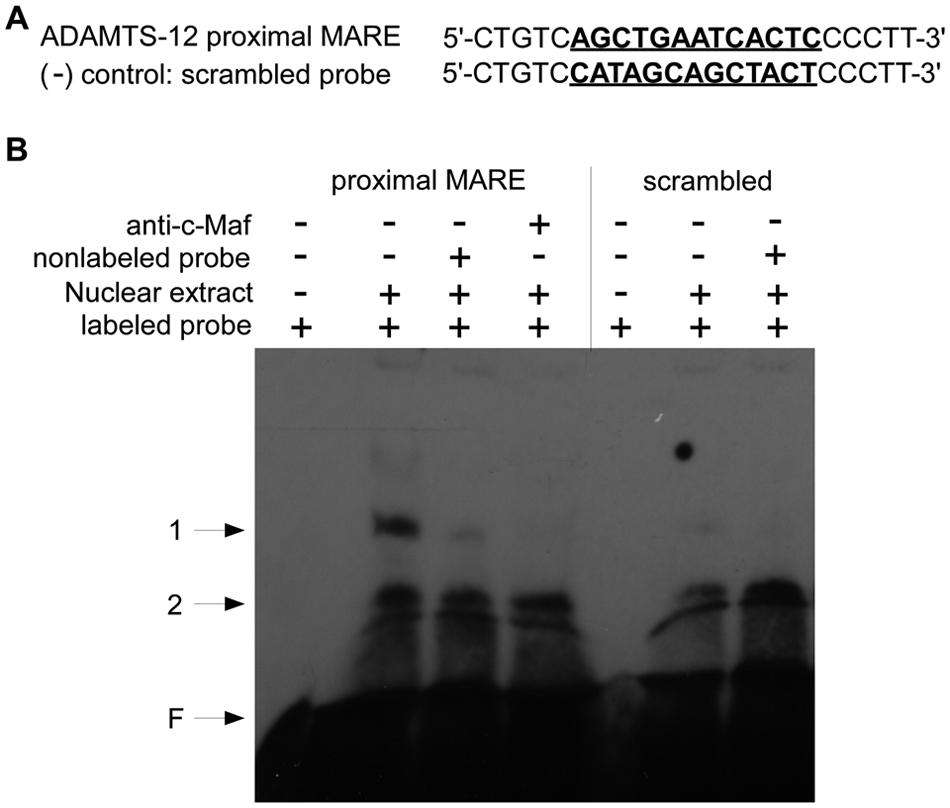
c-Maf binds to the proximal MARE in ADAMTS-12 promoter in vitro. (
Discussion
We found that ADAMTS-12 expression was up-regulated during chondrogenesis in vitro, which is consistent with existing literature. 5 As the regulation of ADAMTS-12 expression remains largely unknown, we further examined the human and mouse ADAMTS-12 promoters for potential transcription factor binding sites and identified several MARE sites for c-Maf binding. Induction of ADAMTS-12 expression during chondrogenesis in vitro coincided with c-Maf induction, suggesting a functional relationship between the two during chondrogenesis. This interaction is substantiated by reports that hypertrophic chondrocytes express c-Maf during mouse embryonic development and that c-Maf facilitates terminal chondrocyte differentiation in postnatal mice.11,12 Of the potential MARE sites we identified in the human ADAMTS-12 promoter, we found that a single MARE site at position -61 was sufficient to confer regulation of ADAMTS-12 expression by c-Maf in our experimental systems.
In addition to cMaf, the expression of ADAMTS-12 mRNA was also shown to be regulated by parathyroid hormone related peptide (PTHrP). 5 In this context, it is possible that ADAMTS-12 may also inhibit hypertrophic chondrocyte differentiation in some circumstances. 5 PTHrP maintains proliferation and inhibits chondrocyte hypertrophy in the physis. 15 ADAMTS-12-null mice have no apparent growth defect but their cartilage has not been specifically examined. 16 Additionally, c-Maf-null mice display advanced terminal chondrocyte differentiation. 11 Hence, future studies will focus on the in vivo localization of ADAMTS-12 and c-Maf in physeal cartilage as well as ADAMTS-12 expression in c-Maf-null mice, to elucidate the functional interaction of these proteins in chondrogenesis and hypertrophy.
In addition to ADAMTS-12, at least two c-Maf target genes are also involved in chondrocyte differentiation, namely, CTGF and matrix metalloproteinase-13 (MMP-13).17,18 CTGF and MMP-13 are localized in hypertrophic chondrocytes during mouse skeletal development. 17 The expression patterns of CTGF and MMP-13 overlap with that of c-Maf.11,17 The CTGF promoter does not contain a consensus MARE-like sequence, whereas the MMP-13 promoter contains sequences highly homologous to T-MARE. Col 2a1 and Col27a1 were also identified as targets of longer transcriptional variant of c-Maf (Lc-Maf).19,20 The Lc-Maf-responsive sequences in Col2a1 and Col27a1 promoters do not share any homology with MARE consensus sequence. Studies have been unable to demonstrate colocalization of Lc-Maf with Col2a1 during precartilaginous mesenchymal condensations or Col27a1 in the proliferating and prehypertrophic chondrocytes.19,20 Finally, Col10a1 is a marker of chondrocyte hypertrophy, but it remains to be determined if c-Maf also regulates Col10a1 expression.
Members of the Maf transcription factor family form heterodimers with other Maf and bZIP transcription factors through their leucine zipper domains. 10 Maf proteins may also interact with non-bZIP proteins. 10 Sox9, an important regulator of chondrogenesis, has been shown to synergize with Lc-Maf to activate Col2a1 and Col27a1 expression.19,20 However, in our system we did not observe a synergistic interaction between Sox9 and c-Maf during the activation of the 3 kbp ADAMTS-12 promoter ( Fig. 4D ). Aberrant expression of c-Maf nonchondrocytic cells has a wide range of effects from proliferation to apoptosis, depending on the cellular context.21-23 We did not observe any change in cell number related to apoptosis or proliferation on c-Maf overexpression in SW1353 and C20/A4 cells (data not shown).
There are two types of MARE sites, T-MARE and C-MARE, which are perfect palindromic 13 and 14 nucleotides sequences, respectively. 9 However, the Maf transcription factors frequently activate many genes through response elements that contain only one half of the palindromic MARE. 10 The MARE consensus sequences generated by analyzing known naturally occurring Maf binding sites revealed that the 5′ half region is more conserved than the 3′ half. 8 Consistent with this observation, we found that the functional c-Maf binding site identified in the ADAMTS-12 promoter has a well-conserved 5′ half region but a less conserved 3′ half region. However, despite being not so well conserved with other MARE sequences throughout the genome, the sequence of the proximal region of the ADAMTS-12 promoter is highly conserved between human and mouse, indicating that this region is important for the regulation of ADAMTS-12 expression across species.
Our population is aging and the prevalence of osteoarthritis is increasing. As of today, there is no cure for osteoarthritis, and thus a greater understanding of the factors that influence cartilage homeostasis and arthritis progression is required. In this article, we show that the cartilage-degrading enzyme ADAMTS-12 is regulated by the transcription factor c-MAF. We demonstrated that ADAMTS-12 is co-expressed with c-Maf during in vitro chondrogenic differentiation. The ADAMTS-12 promoter was up-regulated by c-Maf and this up-regulation occurred via a direct interaction with the proximal c-Maf binding site at position -61 of the ADAMTS-12 promoter. A primary target of ADAMTS-12 proteolytic activity is cartilage oligomeric matrix protein (COMP), and the concentration of COMP fragments in serum is used as a biomarker for arthritis. 3 We have recently shown enhanced activity of TGFβ1 when bound to COMP, 24 and this raises the intriguing possibility that proteolytic activity by ADAMTS-12 may indirectly affect TGFβ activity in osteoarthritic cartilage. Further studies will be required to elucidate a definitive role of c-Maf in regulating ADAMTS-12-mediated matrix degradation during the progression of osteoarthritis and embryonic development.
Footnotes
Acknowledgments and Funding
This work was funded entirely through departmental funds from the UC Davis Department of Orthopaedic Surgery.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical Approval
Human cells were isolated from discarded tissues during voluntary joint replacement surgeries, with appropriate IRB approval and patient consent.