
Abstract
Glioblastoma (GBM) is the most common and malignant primary adult brain cancer. Allelic deletion on chromosome 14q plays an important role in the pathogenesis of GBM, and this site was thought to harbor multiple tumor suppressor genes associated with GBM, a region that also encodes microRNA-203 (miR-203). In this study, we sought to identify the role of miR-203 as a tumor suppressor in the pathogenesis of GBM. We analyzed the miR-203 expression data of GBM patients in 10 normal and 495 tumor tissue samples derived from The Cancer Genome Atlas data set. Quantitative real-time PCR and in situ hybridization in 10 high-grade GBM and 10 low-grade anaplastic astrocytoma tumor samples showed decreased levels of miR-203 expression in anaplastic astrocytoma and GBM tissues and cell lines. Exogenous expression of miR-203 using a plasmid expressing miR-203 precursor (pmiR-203) suppressed glioma cell proliferation, migration, and invasion. We determined that one relevant target of miR-203 was Robo1, given that miR-203 expression decreased mRNA and protein levels as determined by RT-PCR and Western blot analysis. Moreover, cotransfection experiments using a luciferase-based transcription reporter assay have shown direct regulation of Robo1 by miR-203. We also show that Robo1 mediates miR-203 mediated antimigratory functions as up-regulation of Robo1 abrogates miR-203 mediated antimigratory effects. We also show that miR-203 expression suppressed ERK phosphorylation and MMP-9 expression in glioma cells. Furthermore, we demonstrate that miR-203 inhibits migration of the glioma cells by disrupting the Robo1/ERK/MMP-9 signaling axis. Taken together, these studies demonstrate that up-regulation of Robo1 in response to the decrease in miR-203 in glioma cells is responsible for glioma tumor cell migration and invasion.
Introduction
Glioblastoma (GBM) is the most common malignant intracranial tumor. 1 Despite aggressive surgery combined with radiation, chemotherapy, and biological therapy, 2 it remains an enormous therapeutic challenge. There is a recognized need for new approaches based on increased understanding of the biological and molecular nature of these tumors. GBM is characterized by high levels of cellular heterogeneity associated with therapeutic resistance due to multiple genetic alterations involved in the control of cell cycle, growth, apoptosis, invasion, and neovascularization. 3 Recent important developments include the discovery of microRNAs (miRNAs), which have a central role in the regulation of gene expression and are now clearly linked with cancer. 4 Distinct patterns of microRNA expression have been observed in many cancers including glioblastomas, and the functional significance of some of these microRNA alterations is beginning to emerge. 5 These data indicate that miRNAs play a role in multiple hallmark biological characteristics of GBM, including cell proliferation, invasion, and angiogenesis.
MicroRNAs are small, noncoding RNAs, ≈22 nucleotides long that repress gene expression in a variety of eukaryotic organisms with specific target mRNAs. An individual miRNA is capable of regulating dozens of distinct mRNAs. In mammalian cells, miRNAs affect gene silencing via both translational inhibition and mRNA degradation, where they induce mRNA degradation or translational inhibition. 6 The biogenesis of miRNA is a multistep process in which primary miRNAs (pri-miRNA) are cleaved within the nucleus into precursor miRNA (pre-miRNA) by an enzyme complex including Drosha, an RNase III family nuclease. 7 This is followed by processing of pre-miRNAs in the cytoplasm by Dicer, another RNase III that cleaves the double-stranded pre-miRNA to form mature miRNA. 8 The mature miRNA can regulate gene expression of complementary mRNAs by binding to the miRNA recognition element in the 3′-untranslated region (3′UTR) of target mRNAs. 9 Since their discovery in 1993, miRNAs have been shown to play important roles in several cellular processes such as proliferation, differentiation, apoptosis, and development, by simultaneously controlling the expression levels of hundreds of genes. 10 Some miRNAs exhibit a high frequency of DNA copy changes that correlate with altered levels of expression in various human malignancies. 11 Other miRNAs are inactivated by epigenetic mechanisms or lost, and their reexpression can lead to down-regulation of target oncogenes. 12
Recent studies have shown that miRNA expression profiles differ between normal tissues and derived tumors and between tumor types in human cancer. 4 MiRNAs can act as oncogenes or tumor suppressors, exerting a key function in tumorigenesis. 13 Functionally, aberrant miRNA expression can affect cell proliferation, 14 apoptosis, 5 chemo- and radiation-sensitivity, 15 development of metastases, 16 epithelial-mesenchymal transition, and could even potentially define the cancer stem cell phenotype. 17 Furthermore, several clinical studies have identified correlations between miRNA expression and recurrence, development of metastases, and/or survival. 17 These lines of evidence strongly suggest that miRNAs function in tumor promotion and suppression. 18
A potential role for miRNAs in malignancy has been suggested by the location of the genes for several miRNAs at sites of translocation breakpoints or deletions linked to a specific neoplastic state. Allelic deletion on chromosome 14q plays an important role in the pathogenesis of GBM and was thought to harbor multiple tumor suppressor genes associated with GBM. Loss of chromosome 14q was observed in 20% of gliomas in one study, 19 whereas Hu et al. observed such losses in more than 40% of tumors. 20 This region encodes several miRNAs, including microRNA 203 (miR-203). We identified that miR-203 is suppressed in GBM. We also show that reintroduction of miR-203 in GBM cells suppressed tumor cell migration and invasion. Robo1 was identified as a direct functional target of miR-203 in glioma xenograft cells and cell lines. Furthermore, we demonstrate that the antimigratory effects of miR-203 were due to suppression of Robo1/ERK/MMP-9 signaling axis. Our findings define an antimigratory function for miR-203 in GBM and suggest that restoring miR-203 expression might be useful for its clinical management.
Results
miR-203 expression is suppressed in primary glioma tumors
Comparative studies of gene expression between normal and tumor tissues is one of the most widely used strategies for unraveling the molecular circuitry underlying cancer. 21 The Cancer Genome Atlas (TCGA) data are a valuable resource for the identification of novel tumor suppressive miRNAs in GBM tumors as well as other cancers. Large-scale data sets to catalogue cancer alterations were generated by TCGA. 22 In the case of GBM, expression of 534 miRNAs were profiled in 10 normal and 495 tumor tissue samples. A detailed analysis on these data was done by considering only a partial data set. 23 In this present study, we have used the complete data set. Supplementary Table S1 presents statistical analysis done on gene-level data obtained from the TCGA site for miR-203. The details of data normalization and process were provided with the downloaded data and can be accessed at http://cancergenome.nih.gov/. R program (http://www.r-project.org/) was used for the computations. For t test, unequal variance was assumed. The results demonstrate that miR-203 levels are suppressed in tumor samples by 23%.
To further validate the role of miR-203 in glioma progression, we determined the levels of miR-203 using RT-PCR from 10 snap-frozen anaplastic astrocytoma (A1-A10) and 10 GBM (G1-G10) human tissue samples and compared them with 3 normal brain samples (NB1-NB3). We consistently observed that relative miR-203 expression is attenuated in both anaplastic astrocytoma and GBM samples (Fig. 1A). We extended our analysis by assessing miR-203 levels by in situ hybridization analysis of a glioma tissue array and compared them with normal tissues. miR-203 expression was significantly attenuated in glioma tissues compared to normal tissues (Fig. 1B and C).
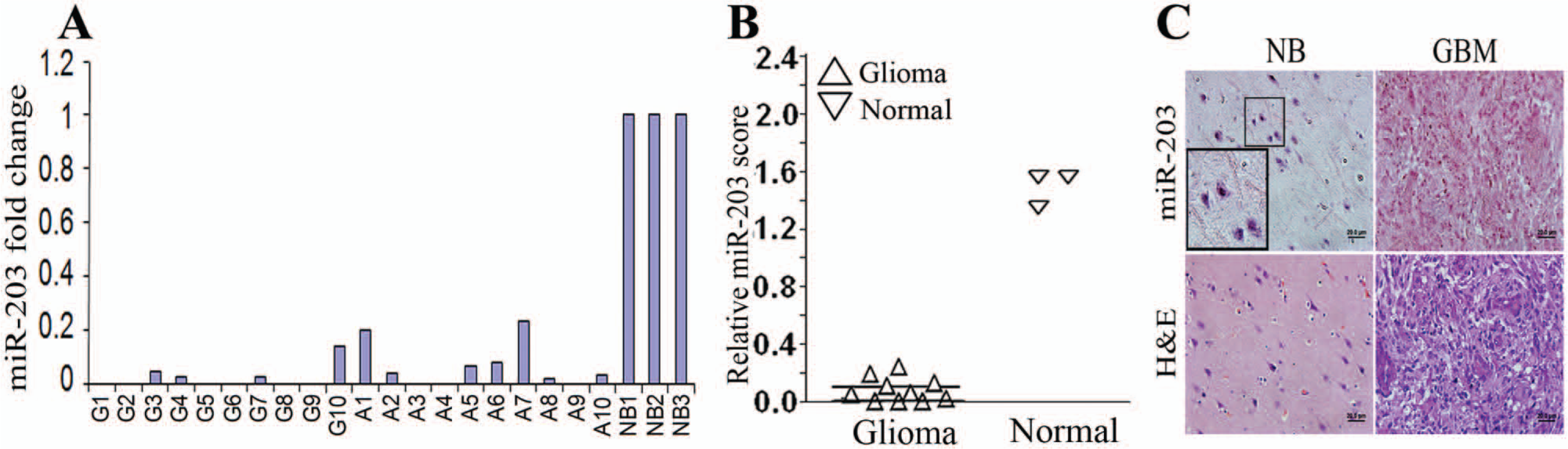
(
MiR-203 expression inhibits migration and invasion potential of glioma cells
In view of the observed inverse correlation between miR-203 expression in glioma tissues, we assessed the potential for an antitumor role of miR-203. Thus, miR-203 was reintroduced in glioma cells (U251) and 2 xenograft cells (4910 and 5310) by transient transfection with plasmid expressing miR-203 precursor (pmiR-203) followed by functional assays. As shown in Figure 2A, transfection of cells with pmiR-203 resulted in 3.5- to 4-fold increase in miR-203 levels compared to mock (PBS) and empty vector (pEV) controls as determined by quantitative PCR. Cell growth rate measured by MTT assay was reduced by 15% at 24 hours and this suppression increased up to 60% by 72 hours in 4910 cell line and U251 glioma xenograft cells transfected with pmiR-203 (Fig. 2B). Migration and invasion are 2 key elements of brain tumor progression. The wound healing and transwell invasion assays were carried out to evaluate the effects of miR-203 expression on the migratory and invasive behavior of glioma cells transfected with pmiR-203 at 36 and 24 hours after transfection, respectively (Fig. 2C and D). Figure 2C indicates that pmiR-203 transfected cells were less proficient than pEV transfected cells at closing an artificial wound created over a confluent monolayer. Also, transwell invasion assays showed that miR-203 reintroduction decreases invasive ability of glioma xenograft cells through the Matrigel basement membrane by ~70% (Fig. 2D), suggesting that miR-203 expression suppresses invasiveness and motility of glioma cells.
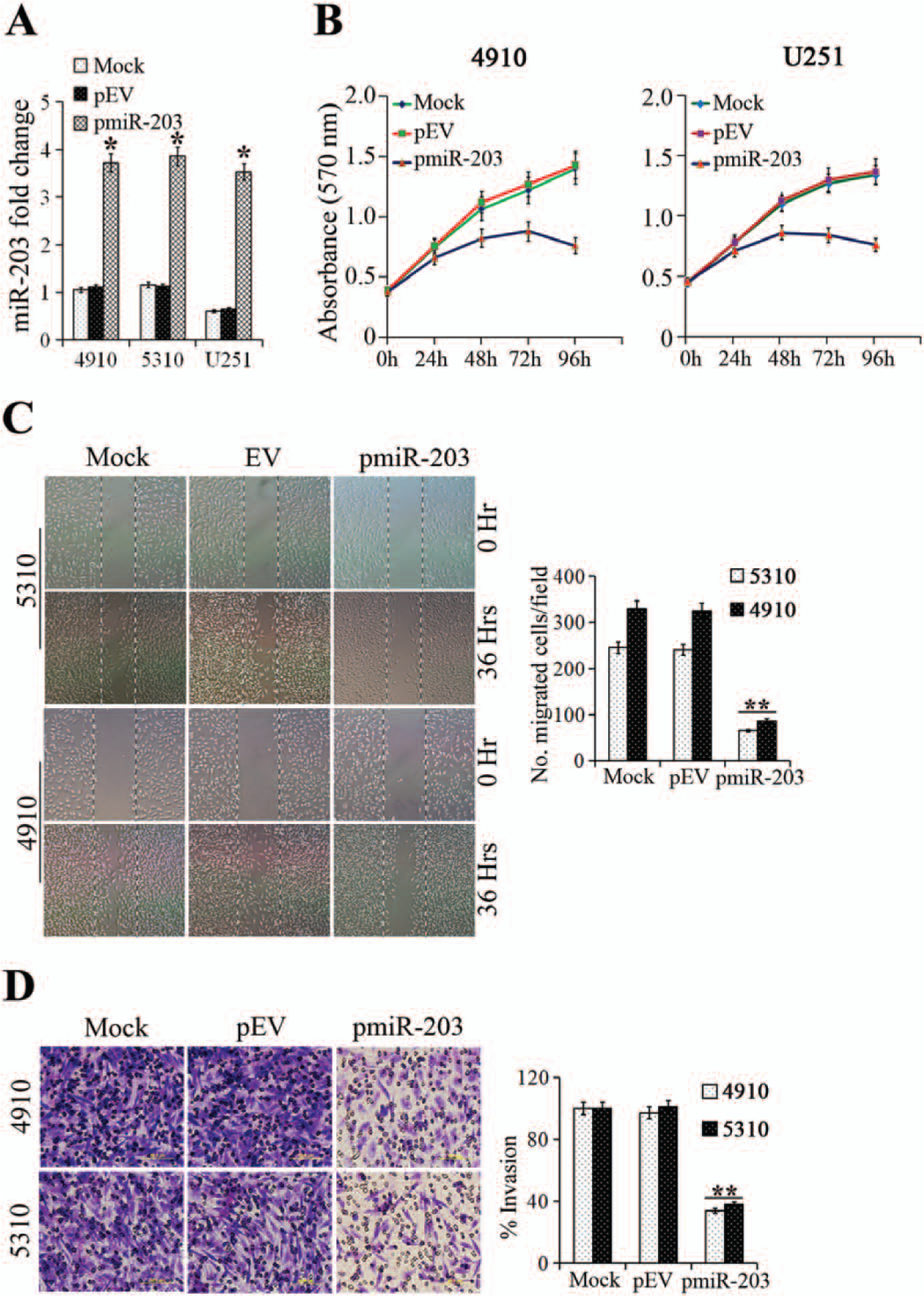
4910, 5310, and U251 cells were transfected with Mock or pEV (empty vector) or pmiR-203 and analyzed for (
MicroRNA 203 targets Robo1 in glioblastoma cells
We next determined the molecular mechanisms underlying the miR-203 mediated antimigratory effects. Analysis using TargetScan 6.0 to search for target genes of miR-203 identified roundabout, an axon guidance receptor, homolog1 (Robo1), a transmembrane receptor of the immunoglobulin family that with SLIT1 and SLIT2 is regarded as a proto-oncogene and harbors migration-promoting activity. 24 Human GBM tumors and cell lines data show that Robo1 expression levels are detected in almost all tumor samples and majority of cell lines with significant P values. 25 Furthermore, gene expression profile from 80 GBM specimens and 4 nontumoral brain samples showed Robo1 with fold change 1.9 (in top 5% of significantly regulated list with P value 8.32 × 10−7). Another study, with 23 samples from epilepsy patients used as nontumor samples and 81 GBMs, showed up to 2-fold increase of Robo1 expression levels. 26 The results demonstrate that Robo1 levels are enhanced in GBM tumor samples, which suggests an inverse correlation between miR-203 expression and Robo1 levels. We therefore determined the levels of Robo1 in glioma xenograft cells and cell lines transfected with miR-203 precursor for 24 hours. Figure 3A indicates that there was a 50% to 60% decrease in Robo1 mRNA levels in glioma cells transfected with pmiR-203. Figure 3B indicates that when adjusted with cells transfected with controls, western blot densitometry analysis revealed about 60% decrease in the expression of Robo1 levels in pmiR-203 transfected U251 cell line and 4910 glioma xenograft cells. It was reported that miRNA may cause either mRNA degradation or translation repression. To obtain further direct evidence that Robo1 is a target of miR-203, we performed analysis of the putative miR-203 binding sites from the Robo1 3′UTR using luciferase assays as previously described. 27 Target sites, that is, 3′UTRs, were amplified and cloned into pHCMV-C-Luciferase after the stop codon of firefly luciferase of the vector. Cotransfection of Robo1 3′UTR-Luc vector with pmiR-203 decreased firefly luciferase activity to about 50% of the vector alone control (Fig. 3C). Further cotransfection of anti-miR-203 and pmiR-203 simultaneously with the Robo1 3′UTR luciferase vector inhibits the effect of pmiR-203 (Fig. 3C). These results suggest that the effects on Robo1 mRNA are specific to miR-203 mediated regulation.
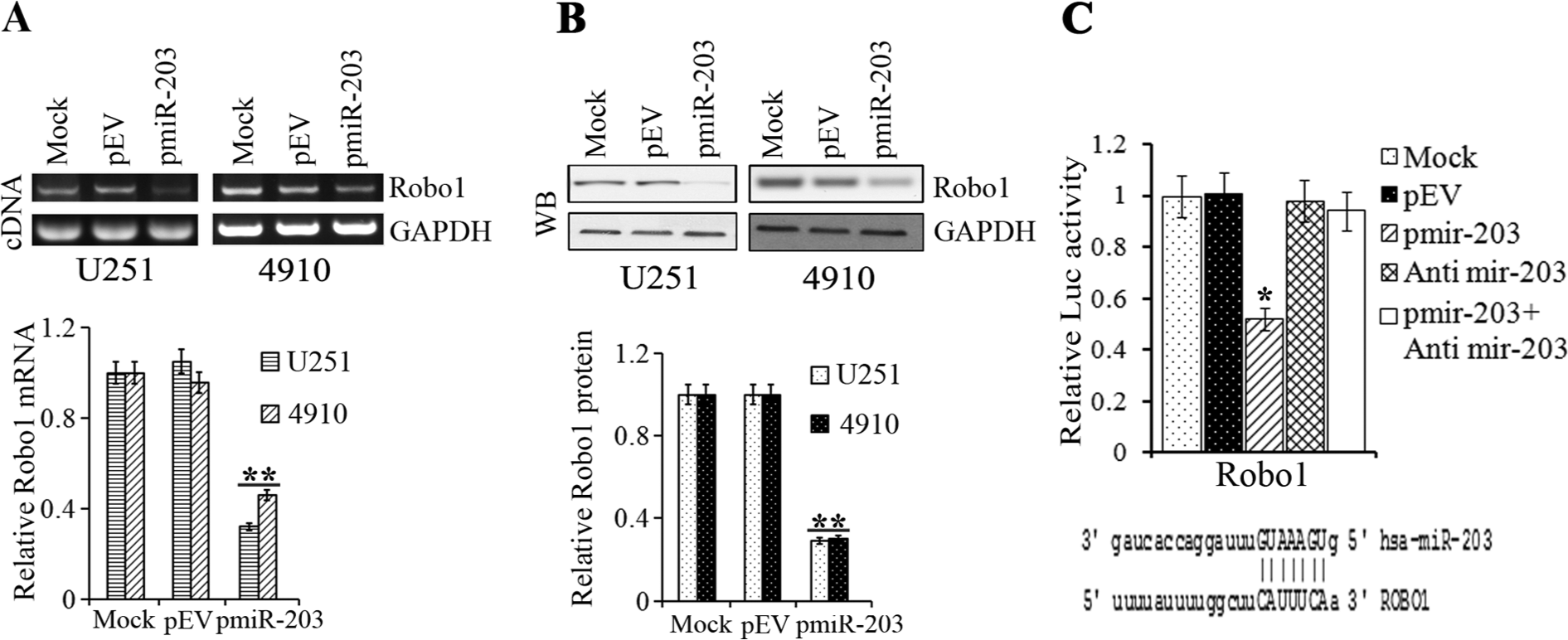
U251, 4910 cells were transfected with Mock or pEV or pmiR-203 and analyzed for (
miR-203 mediated antimigratory effects were attenuated by the overexpression of Robo1 cDNA
Because we observed that Robo1 expression was suppressed in pmiR-203 transfected glioma cells, we first determined the role of Robo1 in glioma migration. Cells were transfected with control SiRNA or pRobo1-Si. The expression of Robo1 protein in the cells transfected with Robo1 siRNA was decreased by 50% to 60% when compared with cells treated with mock and control SiRNA (pSV) (Fig. 4A), indicating that the expression of Robo1 was effectively inhibited by Robo1 siRNA. Subsequent studies showed that the migratory capacity of cells transfected with pRobo1-Si was significantly lower than that of cells treated with control siRNA (Fig. 4B). To determine whether miR-203 antimigratory effect was due to suppression of Robo1, we transfected glioma cells with miR-203 precursor and a plasmid expressing Robo1 cDNA (pRobo1-FL). Overexpression of Robo1 protein by pRobo1-FL plasmid resulted in increased migration of cells (shown as Suppl. Fig. S1). Figure 4C indicates that pRobo1-FL transfection induced the expression of Robo1 by 2.5- to 3-fold in glioma cells compared to mock and empty vector controls (pEV) as determined by western blotting. The migration assay showed that cotransfection of pRobo1-FL along with pmiR-203 could rescue the migratory capacity of glioma cells to that of the controls compared to cells transfected with pmiR-203 alone (Fig. 4B), suggesting that the miR-203 antimigratory effect was mediated, at least partially, by suppressing Robo1 expression.
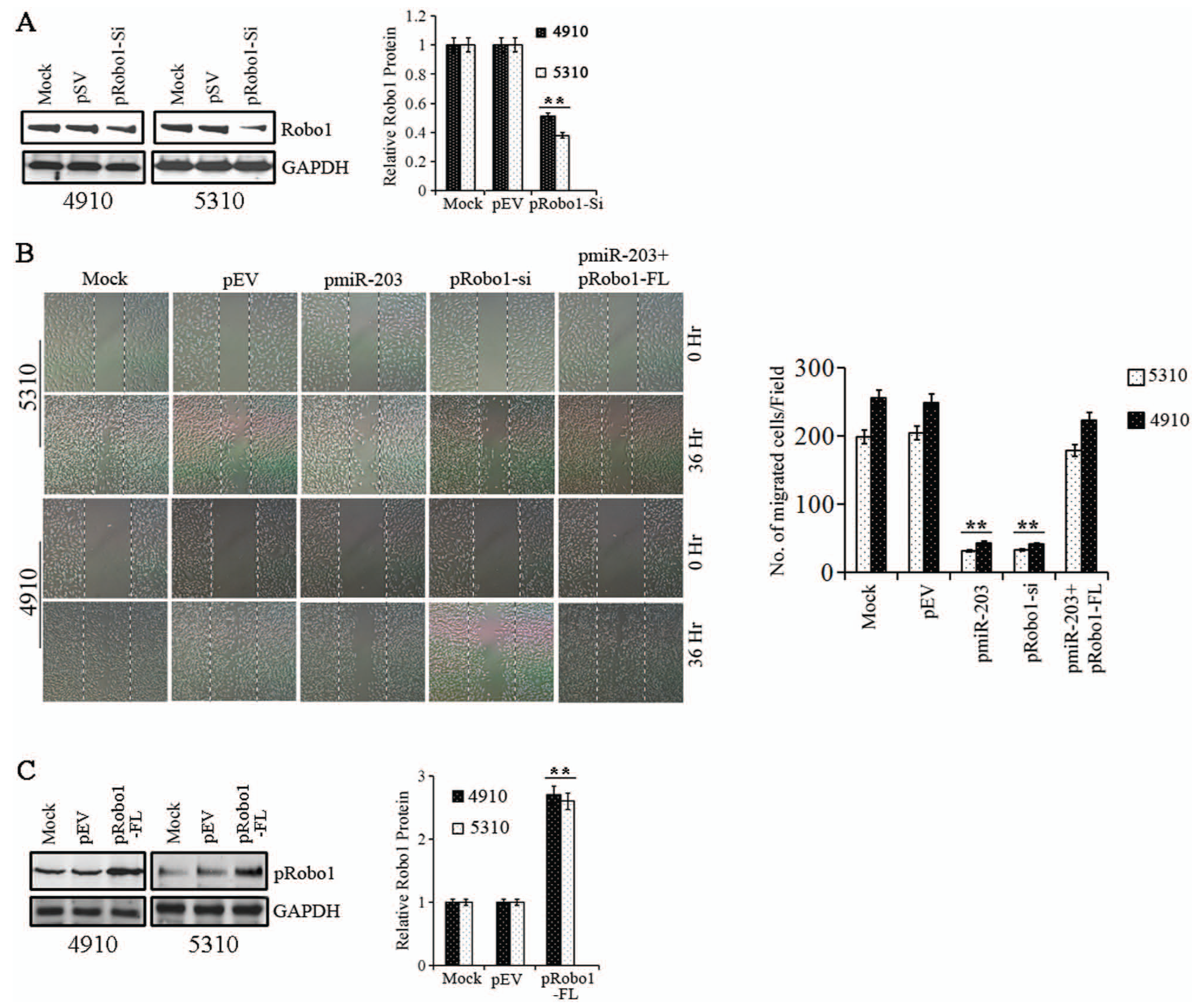
(
miR-203 suppressed Robo1/ERK signaling in glioma cells
We next sought to identify the signaling pathway mediating the miR-203 mediated regulation of glioma migration. Recent studies have implicated the involvement of the ERK1/2 pathway in cell migration. 28 To investigate the potential role of the ERK1/2 signaling pathway in pmiR-203 transfected cells, we examined both ERK activity and cell migration. Immunoblotting with anti-phospho-ERK1/2 (Thr202/Tyr204) monoclonal antibody showed that ERK1/2 phosphorylation (ERK activity) was suppressed by 70% to 50% in cells transfected with pmiR-203 compared to mock and pEV controls (Fig. 5A). However, cotransfection of plasmid pRobo1-FL expressing Robo1 protein along with pmiR-203 restored phosphorylation of ERK1/2 levels to that comparable to controls, suggesting that Robo1 regulates miR-203 mediated ERK1/2 phosphorylation in glioma cells (Fig. 5B). We next used constitutively active ERK construct (ERK 1/2) to study the role of ERK activation on cell migration in glioma cells transfected with pmiR-203. Western blotting indicated that phospho-ERK levels were restored to control levels in cells cotransfected with ERK 1/2 and pmiR-203 compared to cells transfected with pmiR-203 alone (Fig. 5C). However, there was no change in Robo1 levels in cells transfected with ERK 1/2, suggesting that Robo1 functions upstream of ERK signaling (Fig. 5C). Matrigel invasion assay showed that restoration of ERK activity in pmiR-203 transfected cells restored cell invasion comparable to controls (Fig. 5D). Similar results were also obtained with the spheroid migration assay (Suppl. Fig. S2). These results suggest a correlation with ERK signaling and migration in pmiR-203 transfected cells and suggests that miR-203 regulation of glioma cell migration is mediated by Robo1/ERK signaling.
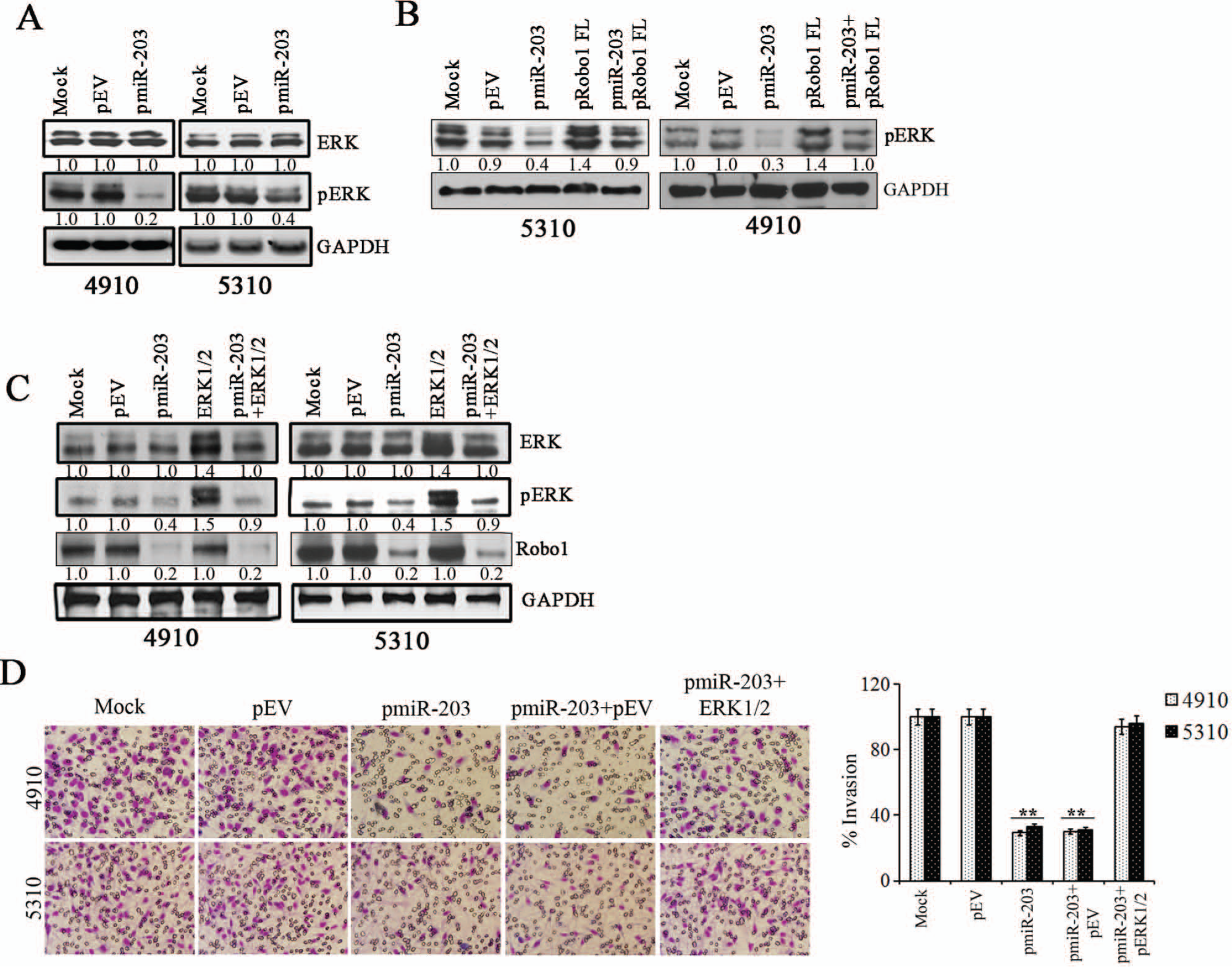
(
miR-203 suppressed MMP-9 expression in glioma cells
To define the role of Robo1/ERK1/2 in cell migration, we performed a PCR array analysis using cell motility RT 2 PCR Array as described in Methods with total RNA isolated from pmiR-203 and pEV transfected glioma cells and control cells for genes involved in cell motility/migration and identified matrix metalloproteinase-9 (MMP-9) as a miR-203 regulated gene. ERKs contribute to the induction of MMP-9 expression, a molecule involved in glioma cell migration and invasion. Because our results from Figure 5 indicate that pmiR-203 suppressed phospho-ERK1/2 levels, we hypothesized that MMP-9 expression could play a role in miR-203 induced effects on glioma migration and invasion. Therefore, we next determined the levels of MMP-9 expression and mRNA levels in pmiR-203 transfected cells. Figure 6A shows that MMP-9 protein and transcript levels were suppressed by 55% to 65% in glioma cells transfected with pmiR-203 compared to mock and pEV transfected cells. In parallel experiments, we determined the effect of silencing MMP-9 expression using the MMP-9 siRNA (pMMP9-Si) plasmid on glioma cell migration. Figure 6B indicates that transfection with plasmid expressing MMP-9 siRNA suppressed MMP-9 levels by 65% to 70% in glioma cells compared to mock and scrambled vector (pSV) transfected controls. The matrigel invasion assay indicated MMP-9 suppression decreased the invasive capacity of glioma cells by 60% to 70% compared to mock and pSV treated controls (Fig. 6C). We next determined the role of MMP-9 in miR-203 mediated antimigratory effects. Figure 6D indicates that cotransfection with plasmid expressing MMP-9 cDNA (pMMP9-FL) in pmiR-203 transfected cells restored MMP-9 levels comparable to that of the controls in glioma cells. However, pMMP9-FL transfection did not alter the levels of Robo1 and phospho-ERK1/2 levels (Fig. 6D). Concomitantly forced expression of MMP-9 using pMMP9-FL along with pmiR-203 effectively reversed the inhibition of cell migration caused by pmiR-203 transfection alone (Fig. 6E). Similar results were also obtained with the spheroid migration assay (Suppl. Fig. S3). These results suggest that the miR-203/Robo1/ERK1/2 axis regulates glioma cancer cell migration by suppressing MMP-9 expression.
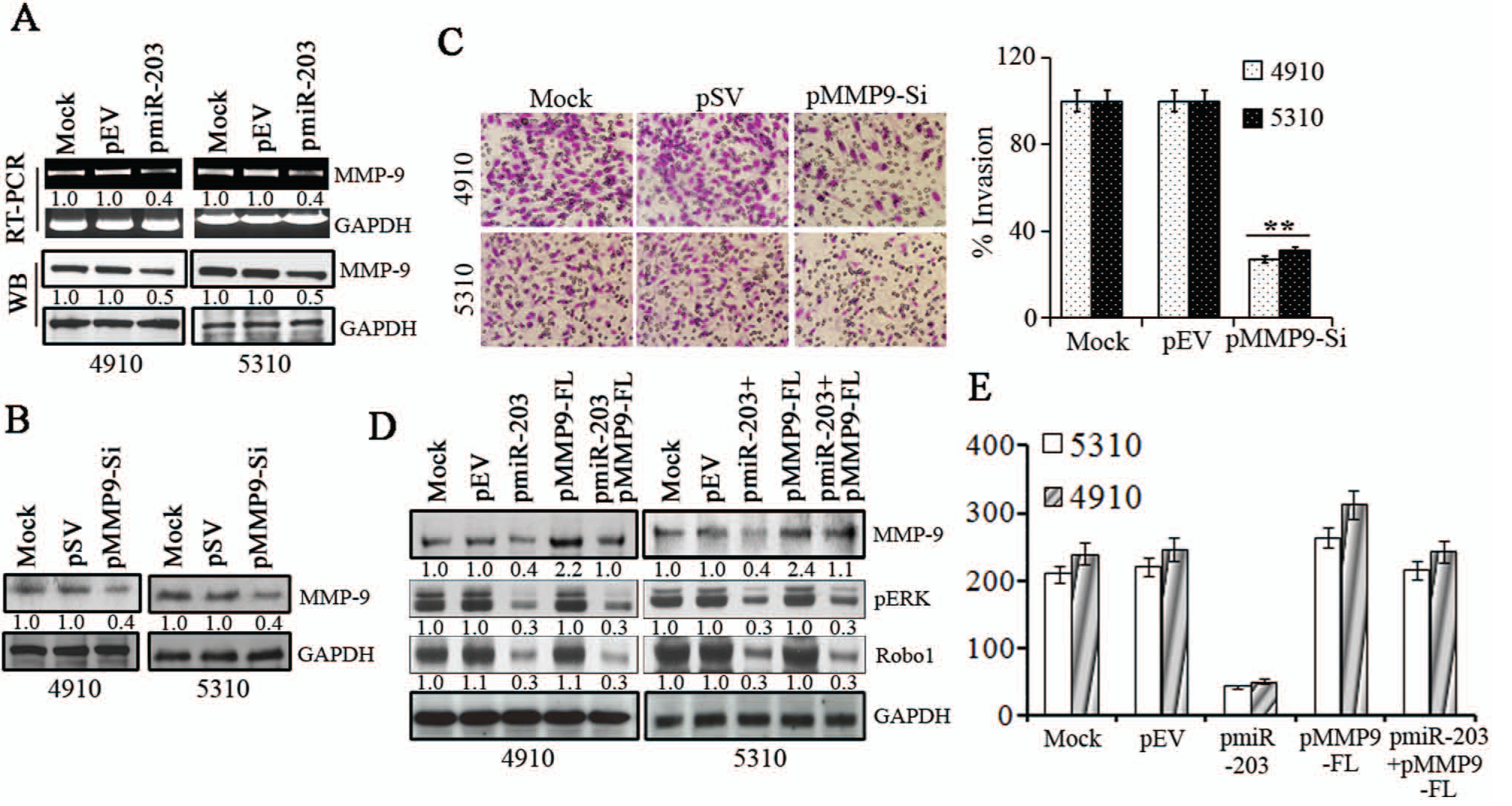
4910, 5310 cells were transfected with Mock or pEV or pmiR-203 and (
Discussion
MicroRNAs regulate mRNA stability and protein expression, and certain miRNAs have been demonstrated to act either as oncogenes or tumor suppressors. 29 Since miRNAs regulate multiple target genes simultaneously, they function as the critical control nodes in the existing tumor signaling network, making them a promising target for cancer treatment. 30 A potential role for miRNAs in malignancy has been suggested by the location of the genes for several miRNAs at sites of translocation breakpoints or deletions linked to a specific neoplastic state. 31 Allelic deletion on chromosome 14q plays an important role in the pathogenesis of GBM, and the site has been thought to harbor multiple tumor suppressor genes associated with GBM, a region that also encodes miR-203. Although previous reports have shown that the overexpression of miR-203 has an impact on growth in prostate, laryngeal, and breast cancer cell lines,32-34 its role in high-grade brain tumors (GBM) has not been elucidated. The TCGA Network comprehensively catalogued the molecular aberrations in 495 high-grade brain tumor, with much remaining to be elucidated regarding the miRNAs. Here, using TCGA data for GBM, RT PCR using glioma brain tumor samples, and in situ hybridization of glioma array, we surveyed the expression levels of miR-203. Collectively, these studies indicate that the expression of miR-203 was suppressed in human brain tumor tissues.
We also show that miR-203 expression suppressed the expression of Roundabout, an axon guidance receptor, homolog 1 (Robo1), a transmembrane receptor of the immunoglobulin family, and interacts with SLIT1 and SLIT2. 24 The SLIT/ROBO1 signaling pathway was described in the nervous system where it regulates axon guidance, branching, and neural migration. 35 Robo1 is regarded as a proto-oncogene and harbors migration-promoting activity. 24 Previous reports suggest that SLIT2 promotes migration in human umbilical vein endothelial cells through Robo1. 24 Furthermore, the engagement of Robo1 by SLIT2 degrades N-cad and enhances motility of neuronal cells. 35 SLIT-ROBO1 signaling may act through both paracrine and autocrine mechanisms to synergistically promote malignant transformation. For instance, a variety of cancer cells secrete SLIT2, which binds to Robo1 expressed on vascular endothelial cells and mediates tumor angiogenesis via a paracrine mechanism. 36 On the other hand, SLIT-ROBO signaling was demonstrated to act through autocrine mechanisms to synergistically promote the malignant transformation of colorectal epithelial cells, 37 suggesting Robo1 blockade could be a novel target as well as provide a molecular tool for cancer therapy.
We show that ERK phosphorylation is suppressed in miR-203 transfected glioma cells. ERK1/2 is a crucial mediator in the Raf/ERK/MAPK signaling axis maintaining diverse cellular functions including cell proliferation, apoptosis, as well as migration. 38 Constitutive activation of ERK1/2 signaling linked increasing cell motility. 39 Our studies also show that constitutive ERK1/2 abrogated miR-203 mediated antimigratory and anti-invasive effects without altering Robo1 levels, suggesting that ERK functions downstream of Robo1 signaling in glioma cells. These studies suggest that the miR-203 antimigratory effect was mediated, at least partially, by suppressing Robo1/ERK signaling. To unravel the mechanisms by which miR-203 affects glioma migration and invasion, microarray analysis of tumor cells transfected with a miR-203 precursor was used to reveal associations between miRNAs and biological functions. Our studies determined that miR-203 expression suppressed MMP-9. Matrix metalloproteinases are capable of digesting various components of the extracellular matrix (ECM) and play a role in cancer cell migration, cell growth, inflammation, and angiogenesis. 40 MMP-9 is highly expressed in human glioma tissues. 41 We show that knockdown of MMP-9 in glioma cells resulted in impaired migration and invasion. We showed that miR-203 suppressed ERK1/2 phosphorylation, a major upstream activator of MMP-9. ERK1/2 signaling is activated in the majority of gliomas, and its inhibition abolished its invasive potential indicating its oncogenic potency. 42 Thus, the down-regulation of multiple pro-tumorigenic genes, Robo1, ERK1/2, and MMP-9, in glioma cells by miR-203 may explain its suppressive effects on migration and invasion of glioma cells. Several studies demonstrate that miR-203 could be a potentially useful prognostic marker and a therapeutic target as the miR-203 regulatory network was shown to play a pivotal role in the progression and metastasis of various cancers. miR-203 can pleiotropically regulate a cohort of metastatic effectors that include ZEB2, Bmi1, and survivin in prostate cancer. 43 miR-203 represents a “stemness-inhibiting miRNA” as it was shown to control the properties of skin stem cells through inhibition of the stem cell factor p63.44,45 Another study showed that miR-203 was also involved in repressing stemness in pancreatic cancer. 46 miR-203 expression is attenuated in colon tumor tissues and overexpression of a miR-203 precursor (Pre-miR-203) reduced Hakai abundance, a direct target for miR-203, with the concomitant suppression of cell proliferation. 47
In summary, we show that the down-regulation of miR-203 contributes to the increased expression of Robo1 (a direct target of miR-203), ERK1/2, and MMP-9 in glioma cells, consequently enhancing glioma cell migration and invasion. Our findings also show that reintroduction of miR-203 in glioma cells suppressed Robo1/ERK1/2 signaling leading to suppression of MMP-9 expression and decreased migration and invasion (Figure 7).
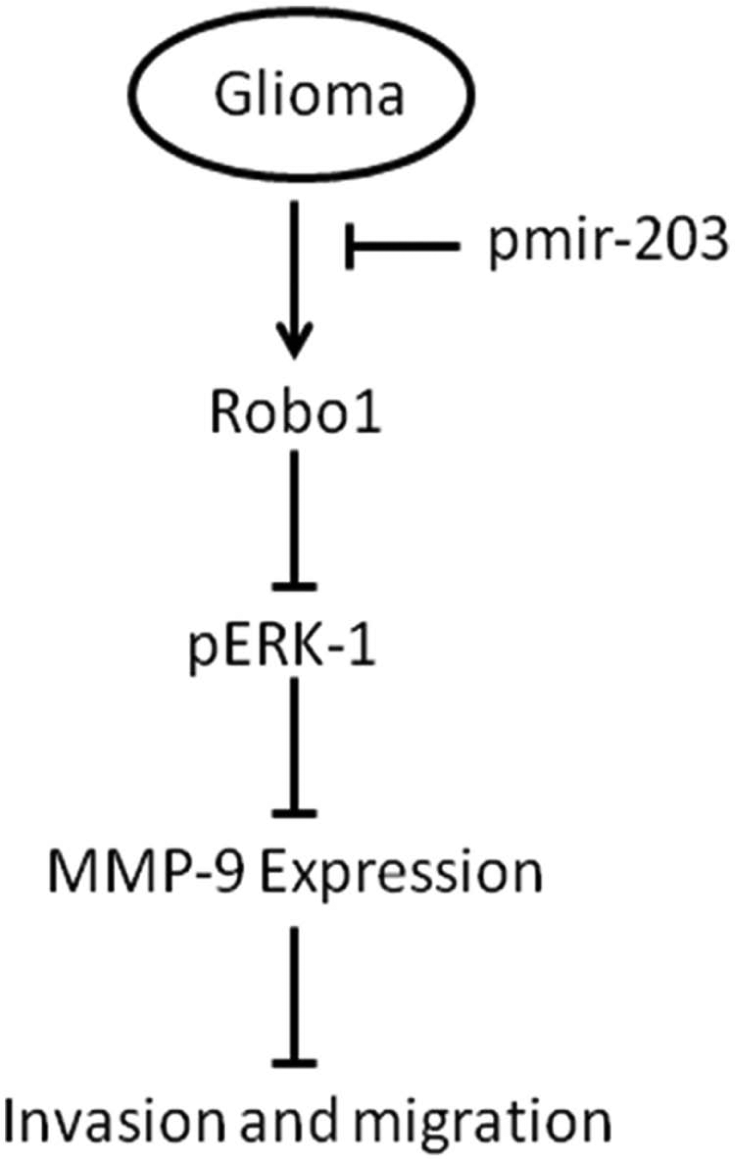
Schematic representation of the inhibitory effect miR-203 in glioma cell migration and invasion. This scheme delineates signaling steps identified in the present study. MicroRNA-203 suppressed Robo1/ERK1/2 signaling leading to suppression of MMP-9 expression and decreased migration and invasion.
Materials and Methods
Cell lines, growth conditions, and antibodies
The glioblastoma xenograft cell lines 4910 and 5310 (kindly provided by Dr. David James, University of California at San Francisco) were generated and maintained in mice and are highly invasive in the mouse brain. 48 Heterotopic tumors were induced in mice by injecting the cells subcutaneously, then the tumor cells were trypsinized and allowed to grow for 3 to 4 passages for consistent results. 4910 and 5310 cells were cultured in RPMI 1640 (Thermo Scientific, Waltham, MA); U251 cells were obtained from ATCC (Manassas, VA) and cultured in DMEM. Media supplemented with 10% fetal bovine serum, 50 mg/mL streptomycin, and 50 U/mL penicillin in a humidified atmosphere containing 5% CO2 at 37°C. All antibodies used in this study were obtained from Santa Cruz Biotechnology (Santa Cruz, CA).
Plasmid construction
PCR primers were obtained from IDT (San Jose, CA); pTarget PCR cloning and expression plasmid were obtained from Promega (Madison, WI). phCMV-C-Luc plasmid was obtained from Genlantis (San Diego, CA). The microRNA 203 precursor (hsa-miR-203) is amplified by PCR using the primers 203-Forward: 5′-AGCTCGGCGAACCGACGGTGTT-3′, 203-Reverse: 5′-CCTGACTGTGACTCTGACTCCA-3′, then cloned into pTarget TA cloning vector. White colonies were selected for screening with restriction enzyme that releases the insert, and the orientation is determined by sequencing. Robo1 3′UTR is amplified by Robo1 UTR-Forward: 5′-GACAACCAAGAGGCTTATGAGA-3′, Robo1 UTR-Reverse: 5′-TGATTATTGGATTTCAGTATGC-3′ primers and cloned into pLuc plasmid after the stop codon. Robo1 full-length cDNA was obtained from Open Biosystems, Thermo Fisher Scientific Inc., then sub-cloned into pCDNA 3.1A plasmid with suitable restriction sites. Robo1 ShRNA oligos Robo1 S 5′-AACCTTGGAATTCTTCCTCTGCGAGTCGTCGGCAGAGGAAGAATTCCAAGGTTTTTT-3′ and Robo1 AS 5′-AAAAAAACCTTGGAATTCTTCCTCTGCCGACGACTCGCAGAGGAAGAATTCCAAGGT-3′ were annealed and cloned into pTarget plasmid, then white colonies were screened, and orientation of insert confirmed by sequencing. MMP-9 Si plasmid was constructed previously. 49
Transfection methods
Cell lines were transfected with empty vector (pEV), scrambled vector (pSV), Robo1 cDNA in pTarget plasmid (pRobo1-FL) hsa-miR-203 micro RNA precursor in pTarget plasmid (pmiR-203), Robo1sh RNA pTarget plasmid (pRobo1 Si). AntagomiR 203 (oligo obtained from IDT, San Jose, CA). A total of 3 × 105 cells were seeded in a 100-mm Petri dish a day before and transfected with 7 µg of plasmid DNA with 21 µL of Fugene 6 (Promega, Madison, WI) in 500 µL of serum-free media (transfection complex) added to the cells in the plate containing 3.5 mL media. After transfection, cells were incubated in serum-containing medium for a minimum of 36 hours.
Western blot analysis
The transfected cells and untransfected Mock cells were collected and lysed in RIPA lysis buffer (50 mmol/mL Tris–HCl [pH 8.0], 150 mmol/mL NaCl, 1% IGEPAL, 0.5% sodium deoxycholate, 0.1% SDS) containing 1 mM sodium orthovanadate, 0.5 mM PMSF, 10 mg/mL aprotinin, 10 mg/mL leupeptin, and resolved on SDS-PAGE and then subjected to western transfer onto nitrocellulose membranes and kept overnight. Blots were blocked with 5% nonfat dry milk in TBST (tris buffer saline with 0.1% Tween-20). Blots were then incubated with primary antibody, followed by incubation with species-specific HRP-conjugated secondary antibody. Signal bands were visualized on HyGlo autoradiography film (Denville Scientific Inc.) after treating the membrane with Thermo Scientific Pierce ECL Western Blotting Substrate. Housekeeping genes: GAPDH or Tubulin (housekeeping genes) antibody was used to verify equal amounts of protein were loaded in all lanes.
Quantitative real-time PCR
Total RNA was extracted from cells using TRIZOL reagent (Invitrogen, Carlsbad, CA) as per standard protocol. Total RNA (10 ng) was used as a template for reverse transcription reaction (Taqman hsa-miR-203 Assay kit, Taqman MicroRNA Reverse Trascription kit, Life Technologies, Carlsbad, CA) in a 15 µL volume, followed by RT PCR with TaqMan Universal Master Mix II, no UNG with the Applied Biosystems (Carlsbad, CA) 7300/7500/7500 Fast Real-Time PCR System, and the results were analyzed by Vii7 RUO software (Invitrogen, Grand Island, NY).
Total RNA was extracted from cells using Trizol reagent (Invitrogen, Carlsbad, CA) as per standard protocol and cDNA synthesized using OligodT primer as per standard protocol. PCR was performed with the primers Robo1 R Forward: 5′-TCCACACAGCAATAGCGAAG-3′, Robo1 R Reverse: 5′-CCTGTAACATGGGCTGGAGT-3′; GAPDH Forward: 5′-TGAAGGTCGGAGTCAACGGATTTGGT-3′, and GAPDH Reverse: 5′-CATGTGGGCCATGAGGTCCACCAC-3′. Robo1 mRNA was quantified by densitometry.
RT2 profiler PCR array
Total RNA was isolated from 4910 glioma cells transfected with pEV and pmiR-203 for 24 hours, and cDNA was synthesized as described above. The human Cell Motility RT2 Profiler PCR Array was used to profile the expression genes related to cell motility according to the manufacturer’s instructions. The fold change of mRNA expression was calculated on the basis of the cycle threshold (Ct) values obtained from the RT PCR experiment. The scatter plot of test versus control samples indicated the validity of the experiment.
In situ hybridization
In situ hybridization was performed on formalin-fixed paraffin-embedded sections (5 µm thickness) of tumor biopsy specimens as previously described. 50 Briefly, after dewaxation, sections were treated with proteinase K (2 µg/mL) at 37°C for 15 minutes, washed, and prehybridized for 1 hour at 49°C. Hybridization with digoxygenin (DIG)-labeled miRCURY LNA probes (Exiqon, Woburn, MA) was performed overnight at 49°C. Slides were then washed at 49°C and incubated with alkaline phosphatase–conjugated sheep anti-DIG Fab fragments (1:1,500; Roche, Indianapolis, IN) for 1 hour at room temperature. The staining was visualized by adding BM purple AP substrate (Roche) according to the manufacturer’s instructions.
Scratch-wound assay
The xenograft cells 5310 and 4910 were seeded into each well of a 6-well plate at a density of 8.3 × 104. Then the cells were transfected with Mock or pEV or pmiR-203, pRobo1 Si, pRobo1-FL, or a combination of pmiR-203 + pRobo1-FL. Transfected cells were grown to confluence and a scratch was made with a sterile 200 µL pipette tip. The cells were kept in medium containing 1% FBS and photographed (Carl Zeiss Invertoscop 40C microscope) at different time points until the scratch was closed in controls. Migration efficiency was quantified by measuring the number of cells that migrated into the wound area after 36 hours of transfection.
Matrigel invasion assay
Matrigel invasion assay was performed using the BD BioCoat Matrigel Invasion Chamber (BD Bioscience, San Jose, CA). Briefly, 2.5 × 104 transfected (pEV or pmiR-203, MMP9 Si, or a combination of pmiR-203 + ERK1/2) or control cells in FBS-free medium were placed in the upper chamber of the insert with Matrigel (for invasion assay). Medium containing 10% FBS was added into the lower chamber. After 24 hours, the cells remaining on the upper membrane were removed by cotton buds, whereas the cells migrating or invading through the membrane were stained with crystal violet and counted under the microscope.
Spheroid migration assay
The xenograft cells 5310 and 4910 were seeded in agarose-coated (1% agarose) 96-well plates and allowed to grow for 48 hours in an incubator with shaking at 100 rpm in standard cell culture conditions. The spheroids formed were transferred to fresh 96-well plates and transfected with Mock or pEV or pmiR-203, pMMP9 Si, pmiR-203 + pMMP9-FL, pmiR-203 + pEV, or a combination of pmiR-203 + ERK1/2 and incubated for 48 hours, and then fixed and stained with Hema-3 stain. Photographs were taken from the control and treated plates and then migration of cells from the spheroids was measured and compared to see the effect of treatments on migration.
3′-UTR luciferase reporter assays
Firefly luciferase reporter plasmids (pHCMV-C-Luc) containing 3′-UTR of the Robo1 gene or empty luciferase vector obtained from Genlantis (San Diego, CA) were used for Luciferase Reporter Assay. 4910 and 5310 cells were cotransfected with the luciferase reporter plasmids together with Mock or pEV or pmiR-203 with and without Anti miR-203 using lipofectamine 2000. Luciferase activity was analyzed 24 hours posttransfection using the Dual Luciferase Reporter Assay System (Promega, Madison, WI) following the manufacturer’s instructions.
Statistics
Statistical significance for experiments was determined by Mann–Whitney U test or Student’s t test. Correlation between the expressions of different genes in the same samples was made using Pearson’s correlation test on log-transformed data. P values <0.05 were considered to be statistically significant.
Footnotes
Authors’ Note
The contents of this article are solely the responsibility of the authors and do not necessarily represent the official views of the National Institutes of Health (the funding agency).
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by the University of Illinois Cancer Center and National Cancer Institute Grant CA132853 (to SSL).