
Abstract
MicroRNAs (miRNA) are small, noncoding RNAs with important regulatory roles in development, differentiation, cell proliferation, and death as well as the complex process of acquired drug resistance. The goal of this study was to identify specific miRNAs and their potential protein targets that confer acquired resistance to gemcitabine in urothelial carcinoma of the bladder (UCB) cell lines. Gemcitabine-resistant cells were established from 6 cell lines following exposure to escalating concentrations of the drug and by passaging cells in the presence of the drug over a 2- to 3-month period. Differential miRNA expression was identified in a microarray format comparing untreated controls with resistant cell lines, representing the maximum tolerated concentration, and results were validated via qRT-PCR. The involvement of specific miRNAs in chemoresistance was confirmed with transfection experiments, followed by clonogenic assays and Western blot analysis. Gemcitabine resistance was generated in 6 UCB cell lines. Microarray analysis comparing miRNA expression between gemcitabine-resistant and parental cells identified the differential expression of 66 miRNAs. Confirmation of differential expression was recorded via qRT-PCR in a subset of these miRNAs. Within this group, let-7b and let-7i exhibited decreased expression, while miR-1290 and miR-138 displayed increased expression levels in gemcitabine-resistant cells. Transfection of pre–miR-138 and pre–miR-1290 into parental cells attenuated cell death after exposure to gemcitabine, while transfection of pre–miR-let-7b and pre–miR-let-7i into the resistant cells augmented cell death. Mucin-4 was up-regulated in gemcitabine-resistant cells. Ectopic expression of let-7i and let-7b in the resistant cells resulted in the down-regulation of mucin-4. These results suggest a role for miRNAs 1290, 138, let-7i, and let-7b in imparting resistance to gemcitabine in UCB cell lines in part through the modulation of mucin-4. Alterations in these miRNAs and/or mucin-4 may constitute a potential therapeutic strategy for improving the efficacy of gemcitabine in UCB.
Introduction
Cancer cells frequently display resistance to chemotherapeutic drugs. Despite the fact that chemotherapeutics encompass a wide range of compounds that interfere with different aspects of the cells’ basic machinery, resistance still occurs. This resistance contributes to disease progression and has become a major obstacle in drug treatment. 1
Drug resistance can be intrinsic or acquired. Intrinsic resistance occurs without prior exposure, while acquired resistance develops during treatment in the absence of initial sensitivity. 1 The activation of drug resistance mechanisms can occur at the genetic level through gene amplification, the transcriptional level through epigenetic modifications, or the proteomic level through mutation or aberrant expression. Additionally, the faulty processing of microRNA (miRNA) coding genes, which then results in the altered function of a targeted miRNA,2,3 can also result in drug resistance.
miRNA are small, nonprotein coding RNA that are typically 20 to 22 nucleotides in length. miRNAs regulate gene expression at the posttranscriptional level by binding to complementary sequences on specific messenger RNA transcripts.4,5 It is suggested that miRNAs control about 60% of the human genome. 6 miRNAs can affect many pathways and are complicated in the respect that one miRNA can have many targets while many miRNAs can share the same target. 7 Interestingly, close to 50% of genes that are known to be regulated by miRNA are located in either cancer-associated genomic regions or fragile genomic areas. 8
Mutation and aberrant expression of many miRNAs have been found in cancer patients9,10 and have led to the study of miRNAs as oncogene regulators and tumor suppressors/promoters. The relationship between miRNA expression and drug resistance appears to be tissue specific, with different tumors demonstrating different profiles. 11 Members of the let-7 family have been found to be underexpressed in several cancers. 3 Restoring let-7 levels in an in vitro model has been shown to restore normalcy and inhibit cancer growth. 3
However, as with all chemotherapeutic agents, resistance does occur. Understanding the mechanism of this resistance through in vitro studies can potentially translate to improved clinical care. In this study, we evaluated the expression pattern of miRNAs between urothelial carcinoma of the bladder (UCB) parental cell lines and cell lines with acquired gemcitabine resistance. We validated a subset of these miRNAs and found that 4 miRNAs showed significantly different expression profiles between these 2 groups. Moreover, restoring these miRNAs to the levels of the parental or resistant cell lines attenuated or augmented cell death, respectively. The mechanism of sensitivity appears to be related, in part, to expression levels of mucin-4, a membrane-bound high molecular weight glycoprotein.
Results
Gemcitabine Sensitivity Profiles of Bladder Cancer Cell Lines
Clonogenic assays
Three poorly (RT4, RT112, CUBIII) and 3 highly (TCCSUP, UM-UC-3, J82) invasive bladder carcinoma cell lines were used in clonogenic assays to assess the effects of gemcitabine. As shown in Figure 1A and 1B, bladder cell lines were treated with various concentrations of gemcitabine, and the IC50 was recorded within a range of 25 to 175 nM. Although the treatment of gemcitabine caused a concentration-dependent inhibition of growth in all 6 of the bladder cell lines, RT4, J82, and TCCSUP cell lines tended to be the most sensitive, whereas UM-UC-3, RT112, and CUBIII cell lines were more resistant.
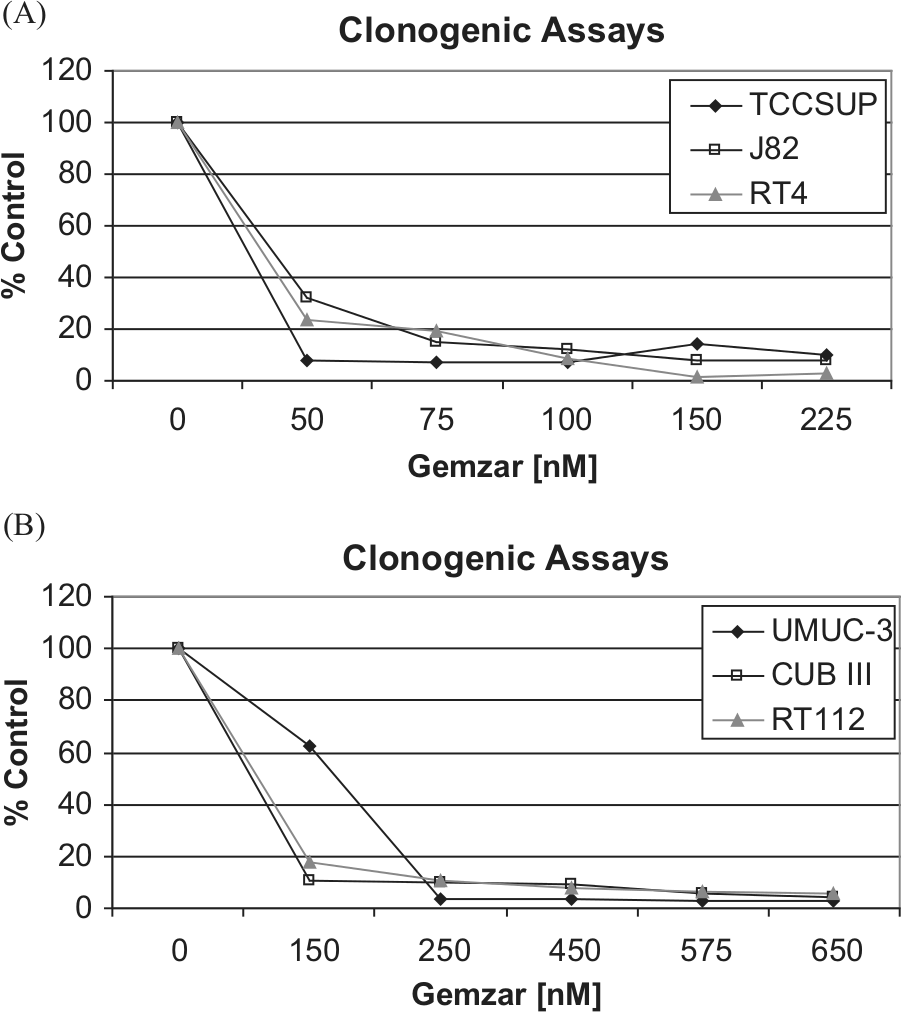
Clonogenic assay results with a panel of bladder carcinoma cell lines following exposure to different concentrations of gemcitabine. (
As no cell line displayed intrinsic gemcitabine resistance (>50% viability), resistant cell lines were established from each of the 6 bladder cell lines by continued exposure to gemcitabine, whose concentrations were serially increased. Stably resistant cells were established following passaging of cells in the presence of gemcitabine over a 2- to 3-month period. Acquisition of resistance to gemcitabine was considered successful when cells survived over multiple passages at a concentration exceeding the IC90 of the parental cell line. Gemcitabine resistance was generated to a maximum concentration of 100 nM in TCCSUP, 150 nM in J82 and RT4, 200 nM in CUBIII, and 450 nM in RT112 and UM-UC-3 cell lines.
Identification of miRNAs differentially expressed in gemcitabine-resistant and parental cell lines
To identify miRNAs differentially expressed between gemcitabine-resistant and -sensitive cell lines, we analyzed the cells that were resistant to the maximum dose of gemcitabine relative to the sensitive parental cells in a microarray format displaying 846 human miRNAs and 424 hsa-miRPlus sequences. The miRPlus sequences are licensed human sequences not yet annotated in the miRBase database. Figure 2 shows a heat map generated from the median normalized microarray data. The median normalized data represent the signal obtained when the replicates (n = 4), for the same miRNA, were averaged. The quantified signals (background corrected) were normalized using the global locally weighted scatterplot smoothing (LOWESS) regression algorithm. Only miRNAs with values above threshold across all samples were analyzed. miRNAs with median fold ratios across sample comparisons of greater than ±2.0 were included in the clustering analysis. Hierarchical clustering using Manhattan distance and average linkage revealed the cell lines cluster together into 2 major groups on the dendrogram. Interestingly, one group contained 2 of the 3 invasive cell lines, whereas the other consisted of all 3 noninvasive and 1 invasive cell lines. The microarray data identified 41 human miRNAs (Table 1) and 25 human miRPlus sequences that were differentially expressed between all of the sensitive and resistant cell lines.
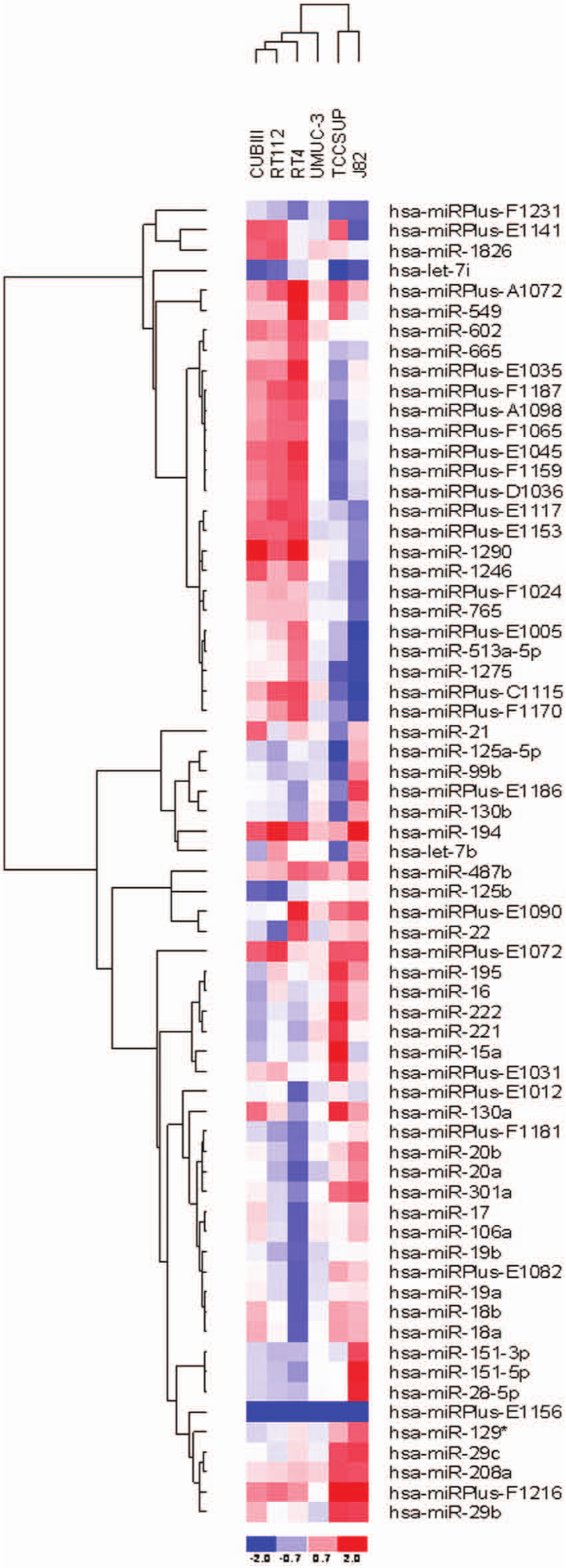
Heat map diagram demonstrating 2-way hierarchical clustering of miRNA and samples. Each column represents a sample, and the rows represent individual miRNA. The red color represents expression above the mean, and blue represents below the mean.
The 41 miRNAs Differentially Expressed across All Samples as Determined by Microarray Analysis of Resistant Cell Lines Relative to Sensitive Parental Lines

Note: Absolute value fold change >2.0. Ranked relative to the maximum fold change observed; * indicates a star sequence; originates from the opposite arm of the hairpin precursor, also known as the passenger strand.
Validation of microarray data by qRT-PCR assay
Expression levels of a small subset of the most highly differentially expressed miRNAs were assessed by qRT-PCR for the parental and resistant cell lines including both the maximally resistant lines and lines with resistance midway between the parental and maximally resistance lines. Figure 3 shows a representation of the expression profile of 4 individual miRNAs within the sensitive and resistant cell lines, confirming the differential expression of specific miRNAs. For most cell lines, both miR-let-7i (Fig. 3A) and miR-let-7b (Fig. 3B) exhibited decreased expression as the dose of gemcitabine was increased. Conversely, analysis of miR-1290 (Fig. 3C) and miR-138 (Fig. 3D) levels showed increased expression in the majority of the resistant cell lines.
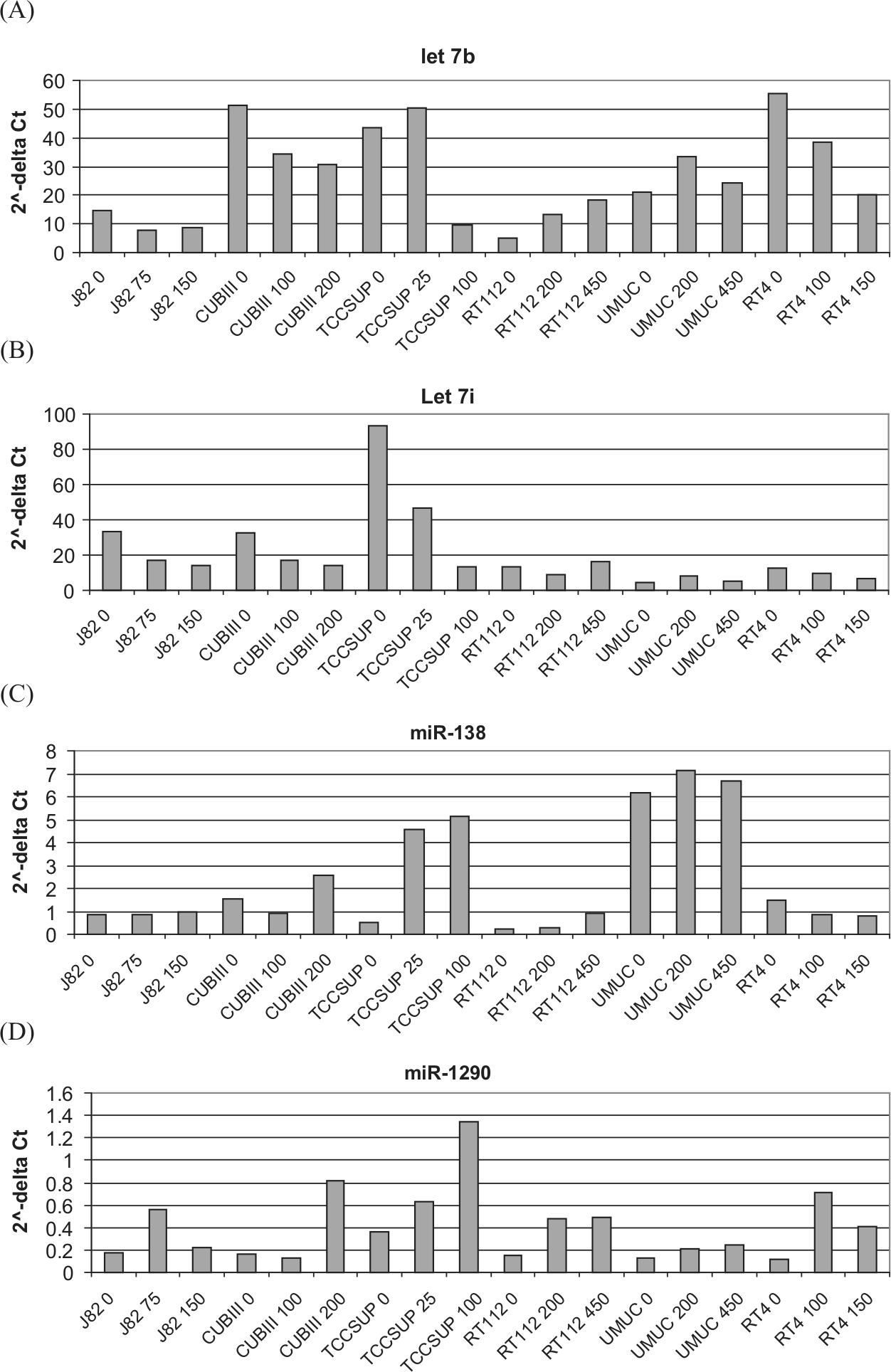
Fold change in the expression of (
miRNAs implicated in gemcitabine resistance alter cell survival
Transfection of pre–miR-138 and pre–miR-1290 into parental cells attenuated cell death after exposure to gemcitabine (Fig. 4A). Introduction of miR-138 and miR-1290 increased the survival 6-fold in RT112 cells. In TCCSUP and CUBIII cells, miR-138 expression led to a 4-fold and 2-fold increase in cell survival, respectively, whereas miR-1290 had no effect in either cell line. Transfection of pre–miR-let-7b and pre–miR-let-7i into the UM-UC-3 and TCCSUP cell lines with acquired resistance to gemcitabine resulted in increased sensitivity in contrast to CUBIII cells in which no difference was detected (Fig. 4B). Overexpression of let-7b resulted in a 20% and 40% decrease in survival in UM-UC-3– and TCCSUP-resistant cells, respectively.
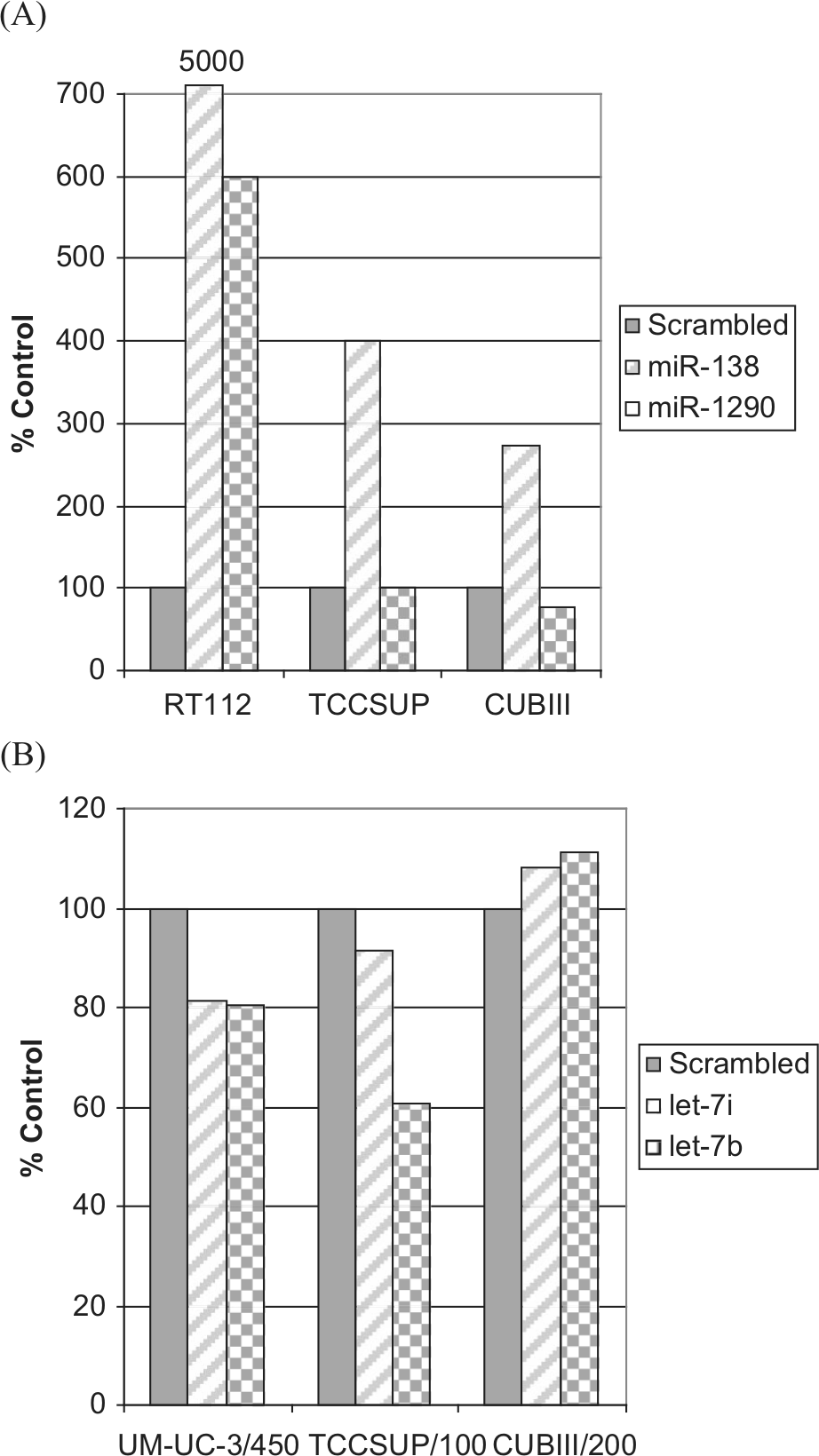
Change in cell survival in (A) parental cell lines transfected with a scrambled control, pre-miR-138, and pre-miR-1290 or (B) resistant cell lines transfected with a scrambled control, pre-miR-let-7i, and pre-miR-let-7b.
Mucin-4 expression in cell lines with acquired resistance to gemcitabine
Expression levels of mucin-4 were evaluated in UM-UC-3 and CUBIII parental and resistant cell lines. Figure 5A shows that mucin-4 was expressed at higher levels in the resistant cell lines as compared to the parental cells. Densitometry showed that the mucin-4 levels were nearly 2-fold higher for CUBIII-resistant cells and more than 10-fold higher for the UM-UC-3–resistant cells.
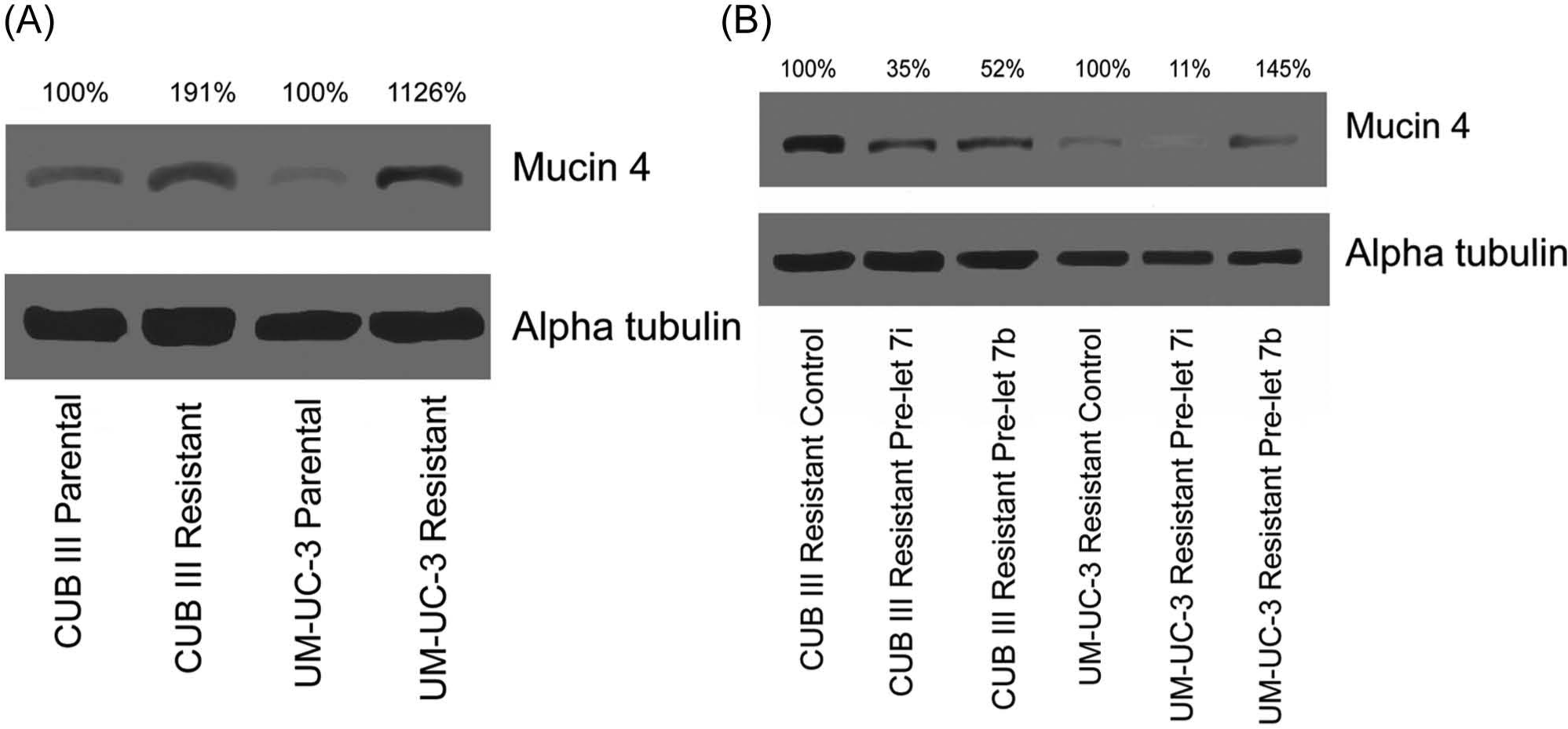
Western blotting of the predicted protein target, mucin-4. (
miRNA modulators of mucin-4 expression
To evaluate a possible role for let-7b and let-7i in the modulation of mucin-4, we have restored the expression of each miRNA in the resistant UM-UC-3 and CUBIII bladder carcinoma cell lines that constitutively express low levels of mucin-4. Figure 5B shows the expression levels of mucin-4 and α-tubulin following the introduction of pre–miR-let-7b, pre–miR-let-7i, and scrambled sequences as controls. Densitometry showed that in CUBIII-resistant lines, the incorporation of both let-7i and let-7b decreased the expression of mucin-4 to nearly one half and one third, respectively. In UM-UC-3–resistant lines, the incorporation of let-7i decreased the expression of mucin-4 nearly 10-fold, while let-7b slightly increased expression.
Discussion
In this study, we report the identification of miRNAs potentially involved in conferring gemcitabine resistance in UCB cell lines. Gemcitabine-resistant cell lines were established following exposure of the cells to escalating concentrations of the drug, and a panel of miRNAs was identified that discriminated between parental and resistant UCB cells. Further, we demonstrated a role for several of these miRNAs in imparting gemcitabine resistance.
The panel of miRNAs that is differentially expressed in resistant cells consisted of 2 members of the let-7 family (let-7b and let-7i), miR-138, and miR-1290. let-7i and let-7b were both found to be down-regulated, while miR-138 and miR-1290 were up-regulated in the resistant UCB lines. The relative importance of each of these miRNAs in conferring resistance was assessed, and we demonstrated that restoration of let-7 family members can modulate cell survival in some cell lines after gemcitabine exposure. This included a 20% to 40% increase in cell death from gemcitabine exposure following restoration of the expression of let-7b and let-7i in UM-UC-3– and TCCSUP-resistant cell lines. Previous studies have also shown that restoring let-7 expression levels inhibits growth by targeting pathways important in cancer cell proliferation.3,12-14 Li et al. 15 have shown that in pancreatic cell lines, several miRNAs including let-7b were correlated with gemcitabine resistance.
Altered expression of miR-138 has been reported in a variety of cancers16-18 as well as miR-1290.19-21 Specifically, Nordentoft et al. 22 reported that decreased levels of miR-138 increased cisplatin sensitivity in some bladder cancer cell lines. However, Wang et al. 23 demonstrated that up-regulation of miR-138 increased the sensitivity of a drug-resistant non–small cell lung cancer line to cisplatin and increased apoptosis. These studies along with our results demonstrate a role for miR-138 in chemoresistance and reiterate the cell-specific functions of miRNAs.
Intravesical therapy, agents that are placed directly into the bladder after tumor removal, has been shown to delay both recurrence and progression in patients with superficial bladder tumors at a high risk of recurrence. The 3 most common drugs used for intravesical chemotherapy are mitomycin C, Bacille Calmette-Guerin (BCG), and doxorubicin. The efficacy of these intravesical treatments is limited, and there is debate over both short- and long-term benefits in the recurrence rate in intermediate- risk tumors.24-26 Tumors can also become refractory to treatment, and surgical removal of the bladder (cystectomy) is often needed when current intravesical treatment regimes have been exhausted.
Gemcitabine (marketed as Gemzar by Eli Lilly and Co. [Indianapolis, IN]) is a nucleoside analog that can replace the nucleic acid cytidine during DNA replication and inhibits ribonucleotide reductase, thereby inhibiting replication and repair and promoting apoptosis. 27 Gemcitabine is used as a chemotherapeutic in various cancers including non–small cell lung cancer, pancreatic cancer, and breast cancer and is the first-line chemotherapeutic drug for metastatic bladder cancer and pancreatic adenocarcinoma.28,29 Additionally, in culture, gemcitabine is selectively toxic to UCB cell lines, suggesting that this treatment might specifically target bladder tumor cells and spare their normal counterparts, reducing overall toxicity.25-27,30,31
Several groups have evaluated the use of gemcitabine as an intravesical treatment for UCB. Bendary et al. 31 concluded that gemcitabine had a better safety profile than BCG and was equally as effective in preventing the recurrence and progression of nonmuscle invasive bladder cancer. Overall recurrence was 25% for gemcitabine-treated patients versus 30% for BCG-treated patients, and progression was equivalent between the groups. Di Lorenzo et al. 32 showed that in carcinoma in situ (CIS) patients with BCG failure, gemcitabine treatment resulted in a recurrence rate of 52.5% (v. 87.5% for BCG-treated patients) and a 2-year recurrence-free survival rate of 19% (v. 3% for BCG). There was no improvement in the mean time to first recurrence or disease progression. Cao et al. 33 also reported a 2-year tumor-free survival improvement for gemcitabine-treated patients with nonmuscle invasive cancer who failed first-line intravesical treatment versus those who received the original therapy (70.8% v. 45.8%, respectively). Borut et al. 34 found a 3-year progression-free survival rate of 81% in patients with invasive bladder cancer treated concurrently with gemcitabine and radiotherapy.
Previously, we have identified changes linked to an epithelial-to-mesenchymal transition (EMT) in UCB tumors in vivo. 35 The EMT phenotype has been shown to be involved in drug resistance to various agents including taxol, vincristine, oxaliplatin, and epidermal growth factor receptor–based therapy.36-38 As cancer cells shift towards a mesenchymal phenotype, they demonstrate lower levels of proteins such as E-cadherin and γ-catenin that promote cell-to-cell adhesion and higher levels of vimentin and N-cadherin.35,39-42 Expression of these mesenchymal markers increases the migratory capability and invasive potential of the cells.43-45 Interestingly, pancreatic cell lines resistant to gemcitabine also showed EMT characteristics, and re-expression of the miR-200 family resulted in a decrease in Zeb1, Slug, and vimentin, consistent with a reversal of EMT. 15 Depending on the resistant cell line tested, we saw similar results (data not shown).
Altered expression of mucin-4 has been reported in various cancers 46 and implicated in tumor cell proliferation.47-49 In previous studies, mucin-4 has been shown to protect pancreatic cancer cells from apoptosis after exposure to gemcitabine.50,51 Mucin-4 has been linked to the susceptibility of pancreatic cells to gemcitabine in numerous studies,50-53 and most recently, hCNT1 and the NF-κB pathway have been identified as mediators of this resistance. 52 Mucin-4 has also been identified as a potential protein target for the let-7 family in multiple miRNA protein target databases.
Based on the literature and database predictions, we evaluated the role of mucin-4 in imparting gemcitabine resistance in UCB cells. Mucin-4 levels were compared in resistant and sensitive cells for 1 invasive (UM-UC-3) and 1 noninvasive (CUBIII) cell line. While both resistant lines exhibited higher levels of mucin-4 compared to their parental (sensitive) cell line, the difference was much greater in the invasive cell line. Interestingly, mucin-4 has also been shown to induce EMT, which then results in an increased invasion potential in epithelial cancer.50,54 The mechanism behind mucin-4’s role in EMT has been shown to involve a cascade effect in which the disruption of the E-cadherin protein increases the activation of other proteins—FAK, MMK7, JNK1/2, and c-JUN—resulting in up-regulation of N-cadherin. N-cadherin then affects AKT, which affects MMP9, resulting in (or leading to) increased motility and invasiveness. 54
In this study, we recorded the elevated expression of mucin-4 in gemcitabine-resistant UCB cells, while transfection of let-7 into these cells results in reduced levels of mucin-4. Together, these results suggest that let-7 plays a role in gemcitabine resistance in UCB cells and that mucin-4 is a target of let-7. Whether it is a primary or secondary target, however, needs to be further elucidated.
Given that intravesical gemcitabine has shown promise in preventing or delaying disease progression, it is important to identify those patients who may benefit most from intravesical gemcitabine. The results from this study suggest a role for miRNAs 1290, 138, let-7i, and let-7b in imparting resistance to gemcitabine in bladder carcinoma cell lines, in part, through the modulation of mucin-4. Alterations in these miRNAs and/or mucin-4 may constitute a potential therapeutic strategy for improving the efficacy of gemcitabine in bladder cancer, and further validation in clinical material is warranted.
Materials and Methods
Cell Culture and Proliferation Assay
The human bladder carcinoma cell lines J82, UM-UC-3, TCCSUP, CUBIII, RT4, and RT112 were maintained in DMEM supplemented with 7.5% fetal bovine serum and penicillin/streptomycin. Gemcitabine-resistant cells were established from UCB cell lines following exposure to escalating concentrations of the drug. Stably resistant cells were established following passaging of cells in the presence of gemcitabine over a 2- to 3-month period.
The effect of each concentration of gemcitabine on the viability of UCB cells was determined by clonogenic assays. Cells were seeded at a density of 1 × 103 cells per 60-mm dish and allowed to attach. After 48 hours, cells were incubated with fresh medium containing an appropriate concentration of gemcitabine (0-600 nM). After 2 days of treatment, the media were removed, and cells were washed with phosphate buffered saline (PBS) and maintained for an additional 7 to 10 days in standard culture media. Cells were then washed 3 times with PBS, fixed for 20 minutes in 3.7% formaldehyde, and stained with crystal violet (0.2%). Following a 15-minute incubation with crystal violet, cells were rinsed in water and dried at room temperature. Surviving colonies with >50 cells were counted without optical aid. All experiments were repeated at least 3 times.
Western Blot Analysis
Subconfluent dishes of cells were washed in PBS followed by lysis in hot sample buffer (2× ESB, 0.08 M Tris, pH 6.8, 0.07 M SDS, 10% glycerol, 0.001% bromophenol blue) and sheared through a 26-gauge needle. Lysates were then assayed for protein concentration using the BSA method (Pierce, Rockford, IL). After determination of protein content, β-mercaptoethanol (1%) was added to each sample. Samples were boiled for 5 minutes, and protein was loaded in each lane of a 7.5% or 12.5% polyacrylamide gel. Proteins were transferred overnight onto nitrocellulose. Membranes were blocked in 10% milk in TBS with 0.05% Tween (TBST) and placed on a primary antibody overnight at 4°C. Antibodies to mucin-4 (Invitrogen, Carlsbad, CA) and α-tubulin (Sigma-Aldrich, St. Louis, MO) were used in Western blot analysis of cell lysates. Following incubation with the primary antibody, blots were washed in TBST 3 times for 15 minutes each, and a secondary antibody linked to horseradish peroxidase was incubated with the blots for 1 hour at room temperature. Blots were then washed as described above and developed with an ECL kit (Amersham, Arlington Heights, IL).
RNA Extraction
Total RNA was extracted from cell lines using the mirVana miRNA Isolation Kit (Ambion, Austin, TX) according to the manufacturer’s protocol. Total RNA was quantified using the Quant-iT RNA Assay Kit (Invitrogen) detected with the Qubit fluorometer (Invitrogen).
Microarray Analysis
Total RNA from cell lines was analyzed using miRCURY LNA Array microRNA Profiling (Exiqon, Vedbaek, Denmark). Briefly, after quality control was performed to determine the RNA integrity, concentration, and content of small RNAs, RNAs were labeled with Hy3 and Hy5 fluorophores using the miRCURY LNA Array Power labeling kit (Exiqon) and hybridized to the miRCURY LNA Array (Exiqon).
Data were obtained as a median of replicated fluorescent signal measurements of the same miRNA from each slide. Slides were analyzed using Exiqon’s protocol. In addition to the raw data for each data point, an expression matrix was performed containing the normalized log2-transformed Hy3/Hy5 ratios. Normalization included background subtractions with a global LOWESS regression algorithm. Spots were flagged and analyzed for signal quality (empty spot, spot signal less than background, less than optimal spot morphology, or saturated spot).
Real-Time qRT-PCR
qRT-PCR quantification of mature miRNAs was conducted using TaqMan miRNA quantification kits following the manufacturer’s protocol (Applied Biosystems, Foster City, CA). Normalization of miRNA expression was performed using the mean of RNU6B and RNU48.
Transfection of pre-miR Constructs
Cells (2.3 × 105) were transfected with 30 nM of pre–miR-138, pre–miR-let-7b, pre–miR-let-7i, pre–miR-1290, or a scrambled pre-miR negative control (Ambion) using siPORT NeoFX transfection reagent (Ambion) according to the manufacturer’s protocol. Seventy-two hours after transfection, cells were harvested for protein analysis and qRT-PCR to confirm the altered expression of individual miRNAs in transfectants.
Statistical Analysis
Comparison of miRNA values between control and resistant cell lines was performed using a t test.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.