
Abstract
Given current urbanization trends, understanding the factors that affect local biodiversity is paramount for designing sound management practices. Existing evidence suggests that the assembly of urban communities is influenced by the environmental filtering of organisms based on their traits. Here, we investigate how environmental characteristics including isolation measurements affect the functional composition of avian assemblages in green spaces of Merida, Mexico, a Neotropical city. We sampled 22 sites, analyzed point-count data collected during fall migration, and characterized the habitat with regard to floristic and structural vegetation attributes, vegetation cover within green spaces, urban infrastructure, and isolation. We assessed the relationship between habitat descriptors and bird functional traits using RLQ and fourth-corner tests and compared trait–environment associations between resident and wintering species. Our results showed that functional composition of resident bird assemblages was linked to the environmental characteristics of the site, while the functional composition of wintering species was not. In particular, the degree of isolation revealed to be an important determinant of trait composition. Plant species richness, particularly native tree and shrub species, were critical for the functional composition of resident birds in green spaces. Our findings suggested shifts in body mass from less to more isolated green spaces. Specifically, we observed that large-bodied species predominated in isolated green spaces. This information is useful given the predicted increases in habitat isolation and transformation of green spaces due to urbanization.
Urban ecosystems are complex dynamic systems where humans are the dominant driving force (Alberti, 2008). Major human-induced transformations within urban areas include the clearing of vegetation, the introduction of non-native plant species, the installation of artificial structures, and the alteration of the quality and quantity of disturbances (Niemela, 2011; Parris, 2016) which can have significant effects on the spatial distribution of urban fauna (Fernandez-Juricic, 2002; González-Oreja et al., 2012; Ortega-Álvarez & MacGregor-Fors, 2010; White et al., 2005). In the face of global urbanization trends (Fragkias et al., 2013), understanding the factors that drive biodiversity patterns in urban areas has become paramount for both environmental science and policy.
Birds stand as one of the most common models to study wildlife responses to urbanization (Murgui & Hedblom, 2017). The majority of urban bird studies are conducted within vegetated green spaces due to their biodiversity conservation potential (Gallo et al., 2017). Urban green spaces can encompass sites that resemble natural habitats to a varying extent: from remnants of the local original vegetation to areas exclusively intended for human use. As a consequence, green spaces can markedly differ in size, can be subject to contrasting management practices, and can be used in distinct ways by visitors, all this variation affecting the conservation value of urban green spaces (Carbó-Ramírez & Zuria, 2011; Fernandez-Juricic, 2002; Tryjanowski et al., 2017).
Current understanding of the influence of green spaces characteristics on species richness and abundance is deep (Nielsen et al., 2014). However, it is equally important to gain an insight into the functional component of green spaces’ biodiversity (Pavoine & Bonsall, 2011). Furthermore, it has been proposed that the assembly of urban communities is determined, in part, by the interaction of environmental filters and species traits (Aronson et al., 2016). An increasing number of publications have documented the influence of bird species’ traits on their susceptibility to urbanization. Commonly assessed traits include trophic guild, migratory status, and body mass (Lees & Moura, 2017), while some authors have broadened the set of traits analyzed considering characteristics such as adult survival rate or innovative behavior (Meffert & Dziock, 2013). Recent works have quantified the functional diversity of urban avifauna through the use of indices (Morelli et al., 2017; Schütz & Schulze, 2015). Although such studies contribute to understand the effect of the filters on the distribution of traits, they do not allow to identify habitat associations with traits. We expect that if environmental factors prevent or favor the establishment of birds in green areas based on their traits, the distribution of species in surveyed green spaces will be heterogeneous, with species holding similar traits responding in a common fashion to habitat characteristics (Kraft et al., 2015).
Understanding how species’ traits are related to environmental characteristics of urban or urbanizing sites is paramount, especially in those areas experiencing or projected to experience elevated urbanization rates such as Mexico. Despite urban bird ecology in Mexico has experienced a rapid growth in recent years (Marzluff, 2017), most of the published information refers to urban areas within the Trans-Mexican Volcanic Belt (Nava-Díaz, 2016), while other important biogeographic regions remain unexplored not to mention. Although previous studies have assessed the responses of resident and migratory species at their breeding grounds (Huste & Boulinier, 2011), studies in their wintering grounds are uncommon (but see Wolff et al., 2018). Furthermore, functional traits information has been missing in urban bird ecology research in Mexico, despite it can contribute to disentangle the relationship between avian communities and urban-related habitat transformations (Silva et al., 2016). To fill an important information gap, we explored trait composition determinants during autumn migration in a Neotropical city. More precisely, this study was aimed (a) to explore how species and trait composition change in urban green spaces, (b) to identify traits that predict species response to habitat characteristics within green spaces, and (c) to compare trait–environment associations between resident and wintering species.
Methods
Study Area
Fieldwork was carried out in Merida (approximately 20.9° N, 89.6° W, 15 m a.s.l.), the main city of the Yucatan Peninsula (YP), southeastern Mexico with more than 1.1 million people (Consejo Nacional de Población, 2015). YP is one of the Mexican biogeographic regions with highest levels of bird species richness (Navarro-Sigüenza et al., 2014), and it holds considerable importance for wintering and transient Nearctic-Neotropical species (Calmé et al., 2015). The area was originally covered by seasonally dry tropical forest characterized by a dry season that may last between 7 and 8 months (Torrescano-Valle & Folan, 2015). Currently, more than 25% of the native species that conform the urban flora belong to the Fabaceae, Euphorbiaceae, and Poaceae families, while common exotic species include Flamboyant (Delonix regia), Golden rain tree (Cassia fistula), and Indian almond (Terminalia catappa) (Peraza-Contreras, 2011).
A total of 22 green spaces were surveyed within Merida Municipality limits (Figure 1). Herein, we use green spaces to refer to urban open spaces dominated by trees and shrubs that are used by humans for several purposes such as recreation, exercise, education, or others, and to which access can be unrestricted or restricted. For this work, we surveyed botanical gardens, public parks, an archaeological site, a zoo, a sport complex, and a reforested area within an industrial plant (Figure 2). Green space size ranged from 0.5 to 39.1 ha and distance to the nearest native vegetation patch ranged from 101 to 5685 m. Some of the surveyed sites harbor artificial waterbodies and were included in the sample to acknowledge the importance of these habitats for local bird diversity given that superficial waterbodies are scarce in YP (Torrescano-Valle & Folan, 2015).

Left: Study Area (Represented as the Red Dot on the Mexico Inset Map) and Overview of the 22 Surveyed Green Spaces Within Merida Municipality, Yucatan, Mexico. Right: Zoom to the Shadowed Area on Left.

Surveyed Green Spaces in the City of Merida Encompass a Wide Variety of Habitats.
Bird Surveys
Bird surveys were conducted during the fall of 2016 from September 27 to November 15. Birds were surveyed using 5-min fixed-width point counts (25 m) separated by at least 150 m. We chose a 25-m radius to increase the probability that all individuals would be detected in all the surveyed habitats (Hutto et al., 1986). The number of sampling points ranged from one to eight and corresponded to green space size. Counts were made during the first 4 hr after sunrise under suitable weather conditions (Ralph et al., 1996). Each point was visited thrice and bird data were collected by a single observer (R. N. D.). All detected birds were included in the avifauna description except Chaetura vauxi and Stelgidopteryx serripennis, since these species were flying over the plots and hence were unlikely to be using the habitat within the plot (Gates, 1997). Statistical analyses were performed with landbirds only since we did not measure the main habitat features that influence the distribution of aquatic birds (Rosa et al., 2003). Raptors were not included in statistical analysis because the count method is not suitable for estimating their numbers (Fuller & Mosher, 1981). For each sampling point, we pooled data from all three visits to get cumulative lists of detected species and generate a species presence/absence table. To determine whether our survey effort was enough to provide a representative sample of the bird community in the time surveyed, we computed the nonparametric incidence-based estimator Jacknife 1 (González-Oreja et al., 2010) using EstimateS ver. 9.1.0 (Colwell, 2013).
Habitat Characterization
We measured nine variables in the field to evaluate the habitat using 25-m-radius circular plots centered on each bird sampling point (Table 1). We considered five classes of environmental variables that could influence the distribution of birds in green spaces. Vegetation composition was evaluated by native tree species richness, exotic tree species richness, native shrub species richness, and exotic shrub species richness. Vegetation structure was assessed by maximum tree height, maximum basal area, maximum bush height, and maximum bush basal area. To quantify the extent of the urban infrastructure in green spaces, we counted the number of poles.
Descriptive Statistics of Environmental Variables Recorded in Green Spaces of Merida, Yucatan.

Variables’ names, abbreviation, description, and the corresponding predictor set are shown. SE = standard error.
Vegetation coverage within each green space was estimated using the Soil Adjusted Vegetation Index (SAVI), which is a modified version of the Normalized Vegetation Index (NDVI) (Huete, 1988). NDVI and SAVI are strongly correlated to several vegetation parameters including vegetation density and percent green vegetation cover (Huete, 1988; Purevdorj et al., 1998). In this study, we employed SAVI because it minimizes errors due to soil substrate optical properties (Huete, 1988). Using a Copernicus Sentinel-2 satellite image and an open source geographic information system (QGIS Development Team, 2020), we computed SAVI using the formula:

NIR = near-infrared band;
red = red band;
L = 0.5.
The quality of images acquired during the bird sampling season was low due to cloud cover so we used a satellite image acquired on January 25, 2017 (cloud cover percentage = 0.0%). The resulting SAVI layer had a 10-m spatial resolution. SAVI values were classified in 12 classes ranging from one (built environment) to 11 (very dense vegetation) and zero values represent water. High-resolution Google Maps images were used to assign SAVI classes that represent vegetation, considering from very scattered vegetation to highly dense vegetation. Once pixels were classified in 1 of these 12 classes, we obtained the total number of pixels for each class. Then, total counts were used to estimate vegetation cover.
Green space area can affect the probability of occupation of species in different fashion (Roberts & King, 2017), so we estimated green space size to introduce it in the models. Similarly, isolation of green spaces can influence the composition structure of urban bird communities (Charre et al., 2013; Fernandez-Juricic, 2002). In this study, we used two alternative approaches to quantify green space isolation. For the first one, we calculated the Euclidean distance from each green space to the closest continuous native vegetation patch. The second approach considered the fact that vegetation cover is not homogeneously distributed through the city and that vegetation cover adjacent to green spaces can influence bird richness and abundance (Shanahan et al., 2011). Therefore, we calculated the extent of vegetation cover in a 100-m width buffer around each green space. Estimations of vegetation cover adjacent to green spaces did not include estimations of vegetation cover within each green space. For this purpose, we employed the same SAVI classes used to identify vegetation cover within green spaces, but in the case of vegetation cover adjacent to green spaces, vegetation-related SAVI classes were grouped in two broad categories: scattered and dense vegetation.
Bird Trait Data
All species in our database were characterized based on three functional traits: diet, foraging strata, and body mass. Diet was expressed as the percentage use of each of the 10 food items categories considered (Table 2). Percentages of diet composition for each species sum to 100. Foraging strata trait was expressed as the estimated percentage use of each one of the seven strata considered. The values of the seven strata sum to 100. Functional traits were sourced from Wilman et al. (2014) and del Hoyo et al. (2018). In addition, residence status in the area was determined based on digital species distribution maps (BirdLife International, 2018) and considering four categories: residents, wintering, transient, and wintering/transient species.
Bird Traits Used in This Study.

Data Analyses
Here, we investigate the effect of potentially influential environmental factors on the functional composition of avian assemblages of urban green spaces through RLQ and fourth-corner tests, which allow to analyze trait-environment relationships (Dray et al., 2014). RLQ is a three-table ordination aimed to identify the main co-structures between an environmental table (R) and a trait table (Q) with the constriction of a species table (L). On the other hand, the fourth-corner approach quantifies and tests the significance of bivariate associations between traits and environmental variables (Dray et al., 2014). RLQ combines three separate ordinations which summarize the main structures of each table. In this way, RLQ relate species traits and environmental variables considering a sites-by-species table (ter Braak et al., 2012). In this study, we employed a binary species table (presence or absence). For the independent ordinations, we followed Borcard et al. (2018) to choose the ordination method based on the type of the variables and to assign rows and columns weights. To test the significance of the association between the environmental and trait tables, several permutation models have been proposed (Thioulouse et al., 2018; see details of permutation models in Borcard et al., 2018). We used a single global test that consists of two independent models whose null hypotheses are species compositions in the sites are not related to environmental conditions of the sites (Model 2) and species distribute according to their environmental preferences but irrespective of their traits (Model 4) (Borcard et al., 2018). The maximum p value of both permutation tests becomes the overall p value to attain a correct Type 1 error (ter Braak et al., 2012).
Measured environmental variables describe broad habitat features, so we grouped them in distinctive sets: vegetation composition (4), vegetation structure (4), vegetation cover (6), and urban infrastructure (1). The fifth set, patch extra descriptors (4) includes green space size and isolation measures. We performed an RLQ analysis including all the predictor sets (that total nineteen variables). In addition, we performed alternative RLQ analyses excluding one of the sets in each ordination since some variables can introduce noise and reduce significance. These RLQ ordinations with reduced environmental tables were obtained to explore the contribution of broad habitat characteristics in determining the functional composition of these bird assemblages. The sum of the correlation L metric of the first two RLQ axes was used as an indicator of the goodness-of-fit of the RLQ ordination (Bernhardt-Romermann et al., 2008). This metric serves to compare the correlation between the trait-based species scores and the environmental-based site scores generated by the RLQ ordination and the correlation of the sites and species scores of the separate ordination of the species table (Thioulouse et al., 2018). The ordinations with better fit are reported and used in the species grouping (further details afterwards), and displayed in the plots.
The fourth-corner analysis tests the relationships between species traits and environmental variables, one at each time (Thioulouse et al., 2018). In this study, we employed jointly fourth-corner analysis and output RLQ axes, which can be interpreted as either environmental gradients or trait syndromes (Dray et al., 2014). Since multiple tests are performed, p values need to be corrected. We set to 9,999 as the number of permutations and used the false discovery rate method to adjust p values in order to avoid Type 1 error (see details in Thioulouse et al., 2018). We decided to report results for which p value < .10 to increase the power of the test given the small sample size (Zar, 2014) and in order to detect likely associations.
To distinguish groups of species that share traits and respond in similar ways to environmental characteristics, we grouped species based on their resulting RLQ scores using Ward’s hierarchical clustering. To determine the optimal number of groups, we considered Calinski–Harabasz index (Borcard et al., 2018). Only those species that occurred in three or more green spaces were included in the analyses to reduce the disproportionately large effects of rare species (Legendre & Gallagher, 2001). We investigated patterns of response of two avian categories: resident and wintering species, so we performed RLQ analysis separately for these subsets of species. The experimental unit in all the multivariate analyses was the green space; hence, data from the sampling points were pooled for each green space. All statistical analyses were performed with R (R Core Team, 2018), applying functions from vegan package (Oksanen et al., 2017), and ade4 package (Dray & Dufour, 2007) and found in Kleyer et al. (2012).
Results
Surveyed green spaces were environmentally heterogeneous according to the measured variables (Figure 2 and Table 1). Some sites harbored exclusively exotic or native plant species, but green spaces with a predominance of native species were majority (∼65%). Considering vegetation structure, tree stratum in green spaces tended to be less than ten meters tall, while shrub stratum height was more evenly distributed, and tree basal area and shrub area were skewed toward low values. Green spaces ranged in size from 0.5 to 39 ha, but most of them (n = 16) were less than 10 ha, and they were scattered through the city, so the distance to native vegetation patches varied from circa 100 m to more than 5,000 m. Vegetation cover within and around green spaces was represented by six classes of SAVI values. Within green spaces, vegetation covered from 51% to 98% of their area, while vegetation cover in the vicinity of green spaces ranged from 1.8% to 51.4%.
Avifauna of Green Spaces in Merida
A total of 87 species from 16 orders and 32 families were detected in green spaces of Merida (Online Appendix 1). Species richness per green space ranged from 9 to 43. Results of the richness estimation indicate that our sample captured 78.4% of the species present in the surveyed space and time. There was a clear predominance of resident birds over migratory ones (Table 3). The most abundant species were Great-tailed Grackle (Quiscalus mexicanus) and White-winged Dove (Zenaida asiatica). Six species were widespread occurring in more than 80% of the green spaces, while 34 species such as Masked Tityra (Tityra semifasciata) or Vermilion Flycatcher (Pyrocephalus rubinus) were detected in just one green space. Aquatic birds, such as Blue-winged Teal (Spatula discors) or Little Blue Heron (Egretta caerulea), were present in only three green spaces and comprised 17 species including migratory ones. We recorded three species endemic to the YP (Calmé et al., 2015): Yucatan Woodpecker (Melanerpes pygmaeus), Yucatan Jay (Cyanocorax yucatanicus), and Orange Oriole (Icterus auratus) and two exotic species in the area: Rock Pigeon (Columba livia) and Eurasian Collared-Dove (Streptopelia decaocto).
Species Richness, Abundance, and Estimated Biomass of Species of Different Categories Recorded in Green Spaces of Merida, Yucatan.

RLQ Ordination for Birds of Urban Green Spaces
The correlation L score indicated that the optimal environmental set for resident and wintering species were similar (Table 4). The environmental table used in the RLQ analysis of resident birds included the vegetation composition, vegetation structure, urban infrastructure, and patch extra descriptor sets. For wintering species, the environmental table consisted of the same descriptor sets except for urban infrastructure. The first two axes accounted for most of the variability explained by the separate ordinations (92% and 81% for resident and wintering species, respectively), so the covariance between environmental variables and species traits was well described by RLQ analysis. Considering the first RLQ axis, variability was better captured for the environmental table (95% for resident birds, 92% for wintering birds) than for the trait table (63% for resident birds, 52% for wintering birds).
Summary of RLQ Ordinations for Resident and Wintering Species With Different Sets of Explanatory Variables.
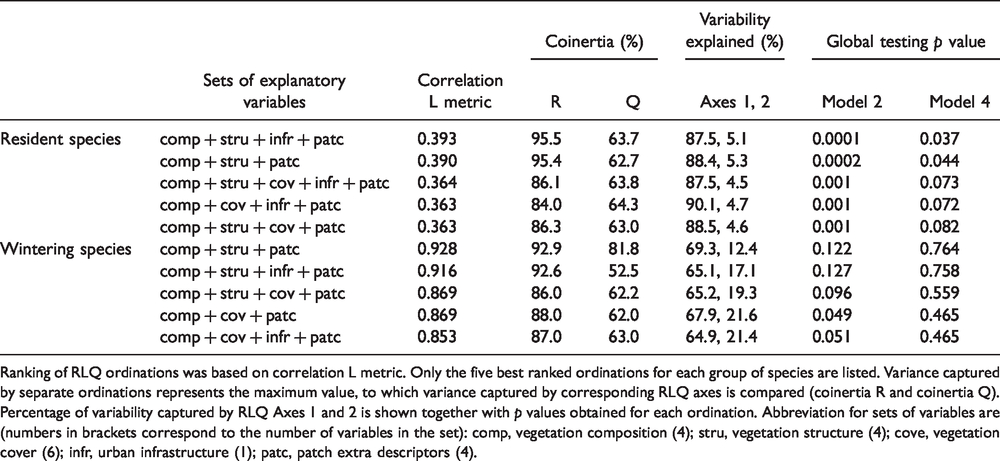
Ranking of RLQ ordinations was based on correlation L metric. Only the five best ranked ordinations for each group of species are listed. Variance captured by separate ordinations represents the maximum value, to which variance captured by corresponding RLQ axes is compared (coinertia R and coinertia Q). Percentage of variability captured by RLQ Axes 1 and 2 is shown together with p values obtained for each ordination. Abbreviation for sets of variables are (numbers in brackets correspond to the number of variables in the set): comp, vegetation composition (4); stru, vegetation structure (4); cove, vegetation cover (6); infr, urban infrastructure (1); patc, patch extra descriptors (4).
Drivers of Functional Composition: Resident Birds
RLQ Axis 1 extracted 87.5% of the co-inertia, and it defined a gradient of green space isolation, driven mainly by decreases in the amount of dense vegetation and increases in the distance to native vegetation patches (Figure 3A). The number of plant species, especially native species of both trees and shrubs, proved to be another relevant driver along RLQ axis, and it showed an opposite association with isolation (Figure 3B). The ordinations indicated that large-bodied species were common in sites far from native vegetation patches and scarcerlysurrounded by dense vegetation, while small-bodied species were present in green spaces that occupy an intermediate position along this axis. Furthermore, canopy-forager species were present in green spaces surrounded by relatively large amounts of dense vegetation and where native flora predominated (Figure 3C). This includes species such as Scrub Euphonia (Euphonia affinis) or Green Jay (Cyanocorax yncas) (Figure 3D). On the contrary, ground-foraging species whose diet is dominated by seed and plant material became more common in sites of reduced native flora richness. RLQ Axis 2 separated green spaces based on the number of exotic tree species, but it only extracted 5.1% of the co-inertia, so we do not consider it for further discussion.

RLQ Analysis Results for Resident and Wintering Bird Species (Left and Right Columns, Respectively). Plots show the ordination of (A) surveyed sites, (B) environmental descriptors, (C) species’ traits, and (D) species. Environmental descriptors and traits significantly associated to Axis 1 are shown with purple, while marginally significant associations are shown with orange labels. Bird species are grouped based on the output RLQ scores, using Ward’s hierarchical clustering. Codes for environmental variables are shown in Table 1.
Global testing showed that the link between traits and environment was significant for resident birds (Model 2 p value: 0.0001; Model 4 p value: 0.03), and this finding was supported by fourth-corner tests on RLQ axes. The joint approach of RLQ analysis and fourth-corner tests indicated that numerous environmental factors of green spaces were related to the functional composition of bird communities (Figure 3B). In particular, the amount of dense vegetation adjacent to green space showed the highest influence on trait composition. Regarding species’ traits, the association of body mass and aerial-foraging strategy with Axis 1 was marginally significant (Figure 3C). There was no evidence for significant associations for RLQ Axis 2. Results from the joint approach of RLQ and fourth-corner analysis are summarized in the ordinations plots (Figure 3B and C).
Drivers of Functional Composition: Wintering Birds
RLQ Axis 1 (69.3% of co-inertia) separated less isolated green spaces harboring a large number of shrub species, both native and exotic ones, from those green spaces with impoverished shrub richness, but with an elevated number of exotic tree species and far from native vegetation patches (Figure 3A). Body size was related to this axis together with the aerial-foraging strategy. Other traits, such as understorey-foraging and midhigh-foraging were not clearly associated with any particular characteristic of green spaces.
Global RLQ testing did not support a significant association between traits and environment (Model 2 p value: 0.12; Model 4 p value: 0.76). However, fourth-corner tests indicated few marginally significant associations (Figure 3B and C): dense vegetation extent and native shrubs richness were the characteristics of green spaces associated with trait composition, whereas body mass and the aerial-foraging strategy revealed an association with the environmental gradient defined by RLQ Axis 1.
Classification of Species
Differences between subsequent values of Calinksi–Harabasz criterion suggested the clustering of resident species in three groups. The ordination plot showed that Groups 2 and 3 clearly occupied opposite extremes of the environmental gradient extracted by Axis 1. Group 1 was the most numerous and included small-bodied species. This group comprised species that feed predominantly on the understorey and midhigh vegetation strata with a diet consisting mainly of invertebrates. On the contrary, Group 2 included large-bodied birds that feed on invertebrates and seeds. Species belonging to this group tend to forage on the ground and use to a much lesser extent other strata. The third group includes medium-sized species with a more diversified use of food items and foraging strata.
Regarding wintering birds, species were clustered in three groups. Species classified in Groups 1 and 3 were small-bodied birds. Although species in Group 1 were mainly insectivores and used the understorey and midhigh strata to forage, species belonging to Group 3 were characterized by the use of the ground, understorey, and canopy strata to forage. Group 2 comprised Summer Tanager (Piranga rubra) only, a large-bodied species with a wider use of foraging strata and a predominantly insectivore diet.
Discussion
This study provided evidence that the functional composition of bird assemblages in urban green spaces can be linked to the characteristics of the sites. More precisely, our study revealed that trait composition of resident bird communities was influenced by several characteristics of green spaces during fall migration. This finding reinforces the well-known idea about the differing sensitivity of bird species to human-induced alterations related to the possession of particular traits (Sacco et al., 2015). In the case of wintering species, our results do not support a significant relationship among the characteristics of green spaces and those species’ traits that we assessed.
The ordinations obtained for resident and wintering species point to the existence of a common environmental gradient along which resident and wintering species distribute. This gradient was defined mainly by the isolation of green spaces but it was also related to the richness of plant species. In the case of resident birds, this was the dominant gradient since it extracted most of the co-variance. Isolation effects have been described for bird species richness in fragmented habitats including urban areas (Charre et al., 2013; MacGregor-Fors et al., 2010; Martensen et al., 2012). Moreover, based on previous findings in forest landscapes and riparian forest parks, we propose that species-dependent responses to isolation in urban green spaces are likely, and these responses could be determined by traits combinations (Martensen et al., 2012).
Our findings suggest that the richness of native trees and shrubs could be a relevant factor for wintering birds. Indeed, the effect of plant richness on taxonomic bird diversity has been well documented for urban areas including green spaces (Nielsen et al., 2014). With reference to the relevant role of plant species for urban birds, here, we contributed with evidence that native trees and shrubs seem to be the component of plant richness associated with functional composition of resident birds during the nonbreeding season. This finding may be attributed to a larger complexity of the habitat in sites with a more diverse native flora, a favorable habitat condition for those species that require a wider diversity of resources such as food, shelter, and perches.
It is important to mention that exterior green spaces in the city of Merida tend to resemble more the original vegetation while inner green spaces usually consist of more landscaped sites. This urban landscape pattern may have important conservation implications since it may be indirectly driving the distribution of species based on their functional traits. There is evidence of the influence of the location of green spaces on its characteristics. For example, those green spaces located in the more urbanized regions of a city tend to be smaller and to include more exotic plant species (Useni-Sikuzani et al., 2018). Hence, we considered that the negative relationship between plant species richness and isolation of green spaces merits deeper examination. Although the inclusion of green spaces of different type may imply confounding factors, we believe our findings are valuable because they suggest the existence of an interplay of factors still scarcely understood.
With regard to vegetation cover within green spaces, the models selected for both resident and wintering birds did not support its contribution to the functional composition of the communities. This was an unexpected result given the existing evidence of vegetation cover as a determinant of several dimensions of bird diversity (Cristaldi et al., 2017; Harvey et al., 2006). We suggest that further research is needed to investigate the relative contribution of vegetation cover, especially shrub and tree cover to bird trait composition within the Neotropical realm, especially in tropical deciduous forests.
Contrary to our expectations, we did not identify clear trait syndromes related to environmental gradients, but our results showed that there was a shift in body mass at the community level along the more-to-less isolated gradient. Results suggest that resident species were filtered along this gradient based on their body size and the strata in which they forage: as green spaces became more isolated smaller birds became uncommon.
When studying the consequences of urban-driven habitat transformation on biodiversity, trait-based approaches allow to obtain more generalizable conclusions by using a set of traits, rather than organisms’ taxonomic identity (Dray & Legendre, 2008; McGill et al., 2006). In accordance with this conceptual approach, species’ trait levels that are subject to the filtering of the environment have been identified for urban birds (Lees & Moura, 2017; Lim & Sodhi, 2004). For instance, omnivorous and insectivorous species were the most frequently encountered species in urban parks of Porto Alegre, Brazil (Scherer et al., 2005). Although works that employ a descriptive approach are relatively common, studies that statistically test the link between traits and environment for urban bird communities are scarce (but see Sacco et al., 2015). Currently, RLQ and fourth-corner tests represent an integrated approach to analyze trait-environment relationships and to determine functional groups (Kleyer et al., 2012). We highlight the fact that RLQ and fourth-corner tests assess trait-environmental relationships considering either trait syndromes or environmental gradients, and not just single traits or environmental descriptors (Almeida et al., 2018; Gamez-Virues et al., 2015). Results obtained this way may deepen our understanding of the mechanisms underlying the sorting of species in the urban environment. So, future research on the assembly of urban bird communities should combine taxonomic and trait information so ecological knowledge advances toward a more general and predictive one (Webb et al., 2010).
Implications for Conservation
To our knowledge, this is the first published study about urban avifauna in YP. Considering urbanization trends, we believe that there is an urgent need to investigate the effects of urbanization in Mexico, especially in areas of evergreen and deciduous tropical forests, the vegetation types with the largest percentages of species richness in Mexico (Navarro-Sigüenza et al., 2014) and that differ from the temperate forest for which information is more abundant. Nearly one fifth (19.0%) of the estimated species richness for Yucatan was recorded in this study (Navarro-Sigüenza et al., 2014) together with 21.4% of the endemic species of YP. Lees and Moura (2017) registered a similar percentage of the regional species pool for the city of Belém, in the Brazilian Amazon. We highlight the relevance of this in the context of migratory species that rest and feed in the area either during the whole winter or during a brief period (Deppe & Rotenberry, 2005). Besides, we emphasize the contribution of waterbodies to bird diversity of the city. Although these waterbodies do not occur naturally, they provide habitat for both resident and migratory species, and this deserves attention considering the karstic origin of the YP and the scarcity of waterbodies in the area (Torrescano-Valle & Folan, 2015).
We strongly recommend to maintain urban green spaces of varying habitat characteristics that comprise from remnants of the original vegetation to landscaped sites. Our results show that habitat characteristics within green spaces and in their vicinity can affect functional composition of bird assemblages. We caution that severe alterations of the habitat can reduce the abundance of species that possess particular traits and this can affect ecosystem functioning (Bovo et al., 2018). Finally, we want to invite urban planners to acknowledge (a) the value that green spaces hold for biodiversity (Carbó-Ramírez & Zuria, 2011) besides its function as public spaces intended for people use, (b) the fact that urban green spaces may represent the only opportunity of locals to experience close contact with wildlife, and (c) that ecological knowledge should be applied to enhance biodiversity in green spaces. If green spaces are intended to conserve local biodiversity, local authorities need to issue guidelines and to set up mechanisms aimed to regulate the management of urban green spaces.
Supplemental Material
sj-pdf-1-trc-10.1177_1940082920923896 - Supplemental material for Drivers of Functional Composition of Bird Assemblages in Green Spaces of a Neotropical City: A Case Study From Merida, Mexico
Supplemental material, sj-pdf-1-trc-10.1177_1940082920923896 for Drivers of Functional Composition of Bird Assemblages in Green Spaces of a Neotropical City: A Case Study From Merida, Mexico by Remedios Nava-Díaz, Rubén Pineda-López and Alfredo Dorantes-Euan in Tropical Conservation Science
Footnotes
Acknowledgment
The authors thank Iriana Zuria Jordán and Luis Hernández Sandoval for their constructive criticism along the preparation of the paper.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Red Temática: Biología, Manejo y Conservación de la Fauna Nativa en Ambientes Antropizados—Consejo Nacional de Ciencia y Tecnología. This study forms part of a doctoral research for which R. N. D. received a scholarship from Consejo Nacional de Ciencia y Tecnología (No. 421382).
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.