
Abstract
The management of assemblages of species across many taxa is a common concern in conservation. Consequently, the use of one or a few surrogate or indicator species to represent an entire assemblage has become an increasingly important tool in conservation science. However, conservation schemes based on the needs of one or two focal species often fail to account for individualistic responses of larger assemblages of species. Data from bird point counts along with vegetation characteristics from a coastal tropical dry forest in Kenya that is subject to elephant disturbance were used to explore the differential responses of bird species to environmental conditions in a forest reserve where wildlife management includes both endangered birds and mammals. Results revealed that even birds with similar foraging habits had idiosyncratic responses to both environmental traits and elephant disturbance. While overall species responded to important characteristics such as percent canopy cover and leaf litter depth, individualistic responses of different species trait diversity defied easy characterization of optimal forest management schemes. Taken together, our analyses highlight the difficulty in basing the development of management plans for entire assemblages of species on the response of a single or a few species. Implications for wildlife conservation in Arabuko-Sokoke Forest and similar forest reserves are discussed, emphasizing the need for a better understanding of individual species’ responses to forest conditions.
Introduction
Habitat loss and degradation are major drivers of biodiversity decline worldwide, and have been the focus of conservation research for decades (BirdLife International, 2005; Brooks & Thompson, 2001; Fahrig, 2003; Waltert, Bobo, Sainge, Fermon, & Mühlenberg, 2005; Wilcox & Murphy, 1985). The detrimental effects of habitat degradation are especially well-documented for birds in both temperate and tropical systems, and include declines in abundance and species richness (SR) as well as genetic diversity (Kettle, 2014; Monkkonen, Rajasarkka, & Lampila, 2014). However, in many cases, individualistic species responses to habitat degradation defy easy characterization of habitat-driven effects on SR and abundance (Lindenmayer, Franklin, & Fischer, 2006). Overall, some habitat structures and configurations can bolster some species, while having neutral or harmful effects on others. This presents challenges for conservation scientists and practitioners involved in both forest restoration and management, especially in cases where the focus is on protecting assemblages of species across taxa that rely on the same vegetation or habitat matrix (de Araújo, 2011; Lindell, Cole, Holl, & Zahawi, 2012). Many conservation schemes rely on choosing a few or a single species to represent an entire assemblage of species (Githiru, Lens, Bennun, & Matthysen, 2007). The response of the “surrogate” species to habitat conditions or disturbance is used to predict how other species in the assemblage will respond, and management protocols are developed based on the needs of the surrogate species. As such, the choice of surrogate species is often based on phylogenetic relatedness, based on the assumption that closely related species will share similar physiological attributes that will result in similar responses to habitat and environmental conditions (Banks, Ackleh, & Stark, 2010). However, relying on a surrogate species approach may fail to account for individualistic responses to certain environmental conditions, so that management prescriptions for an entire assemblage of species that are based on one or two focal species may be convenient but run the risk of poorly serving larger species assemblages (Caro & O’Doherty, 1999; Estrada & Rodríguez-Estrella, 2016; Wiens, Hayward, Holthausen, & Wisdom, 2008)—though these risks may be mitigated by the judicious selection of a suite of surrogates (Morelli, 2015).
Habitat features that may significantly affect forest bird populations include the composition and structure of the canopy (Ndang‘ang’a, Njoroge, & Githiru, 2013), density of understory vegetation (Matiku, Bennun, & Nemeth, 2000; Nemeth & Bennun, 2000), and the availability and abundance of food resources (e.g., insects and seeds). Birds that forage near or on the forest floor should be positively affected by more open understory. Likewise, insectivorous birds should respond positively to higher amounts of leaf litter and insect abundance (Banks et al., 2010; Yen, Thomson, Vesk, & Mac Nally, 2011). In East Africa, elephants are an important disturbance factor and they can significantly alter habitats through physical damage to trees and undergrowth. In such landscapes, we would expect elephant damage to indirectly negatively affect bird populations (Banks et al., 2010; Otieno, Ngala, & Mwalimu, 2014), especially for insectivorous or canopy-dwelling birds that depend on resources in leaf litter or in the canopy. Moreover, birds that are listed as species of concern in the IUCN red-list may be more sensitive to habitat degradation and elephant disturbance of vegetation—though that sensitivity may be a function of how dependent they are on forest habitat as well as their foraging habits (Bennun, Dranzoa, & Pomeroy, 1996). Hence, understanding the effects habitat features and disturbances have on both species diversity and species traits is essential for sound conservation planning.
SR and functional diversity (FD) are both metrics commonly employed to quantify the effects of habitat condition on bird communities (Tilman, 2001). FD describes the range of traits encompassed by an assemblage of species within a community and can shed light on the relationship between biodiversity and ecosystem functioning (Petchey & Gaston, 2002; Van Ruijven & Berendse, 2003). FD is typically better at capturing niche complementarity than SR across both plant (Flynn et al., 2009; Petchey & Gaston, 2006) and animal taxa (Gagic et al., 2015). Traditionally, the focus in bird studies has largely been on how SR changes in response to habitat change (Ding, Yuan, Geng, Link, & Lee, 2005; Jones, 2001), though some have explored bird traits in relation to environmental variables more explicitly. For example, Cleary et al. (2007) found bird traits such as feeding guild were sensitive to logging, while other traits such as threatened status were not. Luck, Lavorel, McIntyre, and Lumb (2012) found positive links between bird foraging behavior and tree cover that may bolster biological control in apple orchards. However, simply examining species responses based on individual traits may not capture important differences within species assemblages, as the interaction between traits may be crucial. Within forest-dwelling birds, for instance, some species show a preference for open understory space, which allows for short, local flight movements as well as unencumbered foraging behavior (Matiku et al., 2000; Nemeth & Bennun, 2000), whereas others do not (Villard, Trzcinski, & Merriam, 1999). These types of individualistic responses make developing management protocols for forest reserves, in which the protection of many taxa is the focus, more complicated. In forests that contain megafauna such as elephants, actively managing forest structure and vegetation condition so that birds and elephants and other species of concern often involves trade-offs necessary to accommodate the needs of several taxa (Banks et al., 2010).
We present here a study that investigates the ties between bird species’ trait interactions and habitat features, with an aim toward better understanding how species respond to environmental change and elephant disturbance. We also discuss the implications for conservation efforts, especially those focused on developing a surrogate species approach to reserve management. Our study illustrates the difficulty in making management decisions to conserve both endangered or declining species as well as broader assemblages of bird species using a case study from the Arabuko-Sokoke Forest (ASF), a tropical dry forest in coastal Kenya. Using data derived from point counts of birds along a dozen forest transects in one of the last large remnant forest tracts in East Africa, we explore the sensitivity of 53 bird species to a suite of forest characteristics pertaining to vegetation structure and disturbance by a resident herd of elephants.
Methods
Study Site
Data were collected in the ASF in the Coast Province of Kenya (Figure 1). ASF is an approximately 420 km2 forest reserve that encompasses the largest remaining patch of indigenous coastal forest in East Africa. It contains three distinct habitat types: (a) Brachystegia woodland (named for the species Brachystegia spiciformis Bentham), (b) Cynometra forest and thicket (named for the species Cynometra webberi Baker f.), and (c) Mixed forest, characterized by a densely packed diverse mixture of tree and shrub species (Nemeth & Bennun 2000). ASF is also home to nine species of birds on the IUCN red list of threatened species (IUCN, 2014), including the Spotted Ground Thrush Zoothera guttata (Endangered), the Sokoke Pipit Anthus sokokensis (Endangered), Sokoke Scops Owl Otus ireneae (Endangered), Amani Sunbird Hedydipna pallidigaster (Endangered), Clarke’s Weaver Ploceus golandi (Endangered), and the East Coast Akalat Sheppardia gunningi sokokensis (Near Threatened; Fishpool & Evans, 2001). Furthermore, the reserve is home to several other globally threatened species, including the Aders’ Duiker Cephalophus adersi (Critically Endangered) and Golden-rumped Elephant Shrew Rhynchocyon chrysopygus (Endangered) as well as a substantial population of African elephants, Loxodonta africana (Vulnerable). The reserve is home to approximately 150 elephants according to the last census conducted (Banks et al., 2010).
Map of study site in Arabuko-Sokoke Forest, Kenya.
Bird Diversity: Point Counts
Coordinates of Transect Sampling Areas in Arabuko-Sokoke Forest.

Vegetation or Habitat Characteristics
A suite of eight metrics pertaining to habitat and habitat condition were measured along each transect; to generate characteristic values, measurements were taken in circular plots of 22.6 m in diameter (encompassing an area of 400 m2) at points 300 m and 800 m from the start of each transect. Measurements taken were leaf litter depth, percent cover of grass, percent cover of shrubs, percent area comprised of short trees, percent area comprised of tall trees, percent canopy cover, and presence or absence of elephant trampling damage (scored as 0 [no elephant damage] or 1 [damage evident]). For five of the characteristics, namely percent cover of grass (ground vegetation <1 m in height), shrubs (1–3 m in height), short trees (woody plants 3–8 m in height), tall trees (woody plants >8 m in height), and canopy cover, four measurements (each corresponding to one quadrant of the circular sampling plot) were taken at each sampling point located at 300 m and 800 m along each transect; values at each sampling point were averaged, and then the mean of the values from both sampling points were calculated to produce a value for each characteristic for the entire transect. Similarly, leaf litter depth (cm) was measured once in each of the plots 300 m and 800 m along each transect and averaged. Finally, in order to measure understory visibility, the number of squares visible (unobscured by vegetation) on a 50 cm × 50 cm colored checkerboard (with 10 cm × 10 cm squares) held up at 1.25 m above the ground and at 5 m distance from the center of the plot were recorded in two different directions (right and left of the transect; MacArthur & MacArthur, 1961; Wilder, Brooks, & Lens, 1998). As with the other measurements, checkerboard values were measured at 300 m and 800 m points along each transect and were averaged to generate one value for each transect.
Bird Traits
Each bird species recorded in the study was classified according to their degree of dependence on forest habitat: those dependent on forest habitat (“Forest specialists” and “Forest generalists”) and those not dependent upon it (“Forest Visitors”). Forest specialists are species that thrive and generally breed in interior, undisturbed forest, and are only occasionally found in secondary or disturbed forest. Forest generalists are also sometimes found in forest interiors but are more commonly found in secondary or disturbed forest habitat (Bennun et al., 1996). Furthermore, birds were classified according to typical foraging position (Ground, Mid-Canopy, or Canopy) and feeding guild (feeding (a) primarily on insects, or insects plus nectar or bee wax, (b) primarily on fruit, or fruit plus insects, (c) seeds, or (d) other animals [predatory]). Finally, IUCN status and population trend (e.g., “Stable,” “Increasing,” or “Decreasing”), which can be related to assemblages of species traits (Jeppsson & Forslund, 2014; Kormann et al., 2015), was noted but not used to calculate FD. Thus, three classifications (Forest dependency, Feeding position, and Feeding guild) were used throughout the analyses as bird functional traits.
Statistical Analysis
We used analysis of covariance (ANCOVA) to test the effects of habitat variables on bird diversity per transect. Bird diversity was calculated as SR and functional richness (Petchey & Gaston, 2006). Functional richness is a measure of niche complementarity and it is calculated as the total branch length of a functional dendogram. To avoid multicollinearity and overfitting of the models, we reduced the habitat data set to three response variables (elephant damage, leaf litter, and canopy cover) after excluding highly related variables based on Hill and Smith (1976) ordination (Figure 2). Canopy cover was log transformed to meet the model assumptions. We validated normality and homosedasticity assumptions of the models by visually inspecting the model residuals.
Hill & Smith ordination, indicating collinearity among environmental factors measured.
To identify the bird traits responsible for the changes in bird communities along measured environmental gradients, we followed Redundancy Analyses (RDA)-sRTA methods described in Kleyer et al. (2012). First, we extracted species scores from RDA. The species scores correspond to the species responses to environmental gradients (Kleyer et al., 2012). We performed three separate partial RDAs where only one habitat variable was kept as constraining and the other two as conditioning variables.
Second, regression trees were used to select those traits that best predicted individual responses to environmental variables. This is a nonparametric approach that produces a binary tree that is built through recursive partitioning (Kleyer et al., 2012). For our analysis, functional traits were the predictors and the species responses (RDA species scores) were the predicted values. The trait that best distinguishes species responses splits the tree into two groups; then, within each subset, another trait splits the groups further. In this way, we could detect not only the main effects of traits but also important trait interactions. Each leaf of the tree had to contain at least four species. We used a pruned regression tree obtained by reducing a full-grown regression tree, with cross-validation (based on minimization of the cross-validated error “xerror”) determining the extent of reduction. All analyses were performed in R (R Core Team, 2015), using the “FD” (Laliberté, Legendre, & Shipley, 2014) and “rpart” package (Therneau, Atkinson, & Ripley, 2015).
Results
Fifty-three bird species were recorded in the bird surveys and included in the analyses (three species were excluded because they were clearly transient species and not normally found in forest habitat; Appendix A).
Hill–Smith Ordination revealed that species responses were clustered around several key environmental factors; some of those factors (such as checkerboard measurements and leaf litter depth, percentage of tall trees, and canopy cover) were redundant (Figure 2). Further analysis was conducted on the reduced set of three factors: elephant presence, leaf litter, and canopy cover. Habitat variables showed large variation among transects (e.g., leaf litter varied from 0 to 2.5 cm, and canopy cover from 28.12 to 66.25%).
SR was invariant across all of the investigated habitat variables, while bird FD declined with increasing canopy cover (−1.5782 ± 0.6019, p = .03, Figure 3).
Relationship between canopy cover and (a) species richness and (b) functional diversity..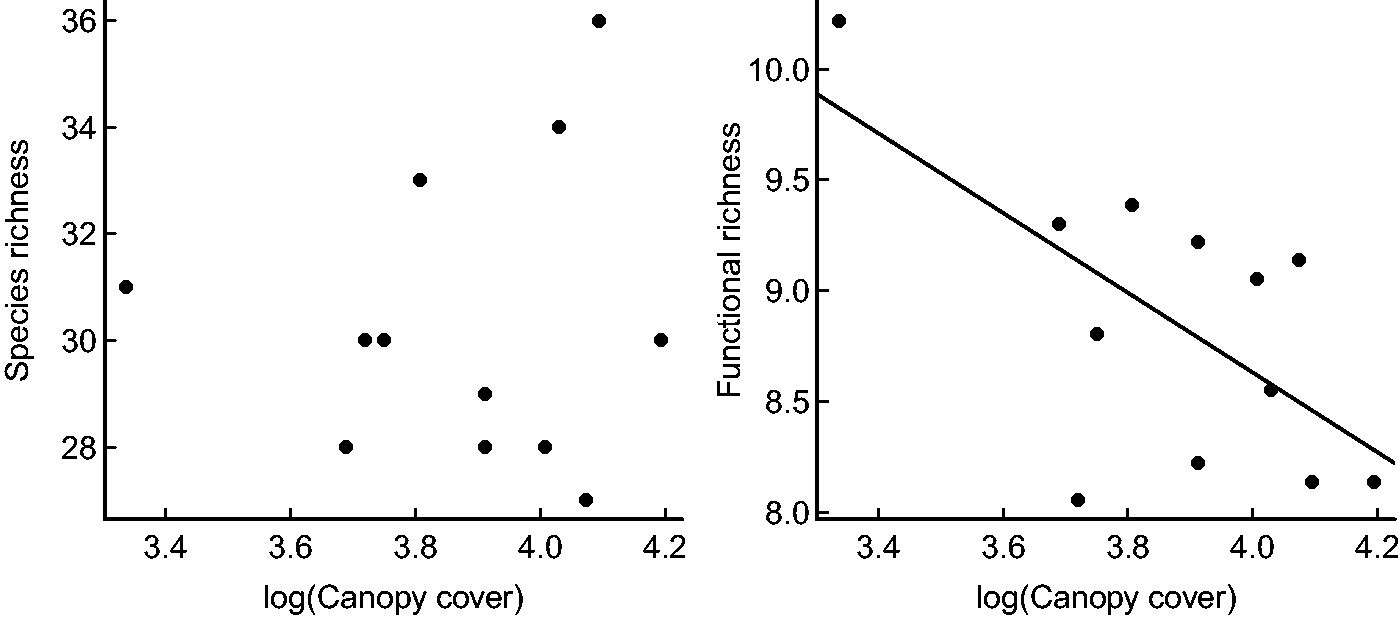
Each environmental gradient explained similar amounts of variation in species composition in the partial RDAs: canopy cover (8.50%), leaf litter (10.50%), and elephant damage (9.84%). The pruned regression tree focusing on the species response to leaf litter selected forest dependency as the best predictor (variable importance = 61) splitting species into two groups: species not dependent on forest (forest visitors, n = 17, deviance = 0.35, yval = 0.094) and species dependent on forest (forest generalists and specialists (n = 36, deviance = 0.49, yval = −0.016; Figure 4). Furthermore, forest visitors were split in two groups (variable importance = 37) based on feeding guild: feeding on insects or seeds (n = 13, deviance = 0.12, yval = 0.13) and feeding on fruits, mixed fruit and insect diet, or predators (n = 4, deviance = 0.14, yval = −0.03) resulting in three functional groups in total (Figure 4). The explained variation for the leaf litter tree with one split (forest dependency) was 14.28%.
Results of RDA analysis using pruned regression tree for leaf litter.
Pruned regression trees focusing on elephant damage and canopy cover revealed no good predictors among investigated traits. See Appendix B for the complete list of univariate RDA scores.
Discussion
Our results illustrate some of the challenges associated with establishing a consistent, systematic management protocol for protecting entire assemblages of species within a forest reserve. We show that although canopy cover had an effect on overall bird trait diversity (functional richness), individual bird species traits were more responsive to changes in leaf litter. Within the array of bird species studied, there were widely divergent habitat preferences among groups sharing the traits we tested. Important distinguishing traits were the degree of dependence on forest habitat and feeding guild, while typical foraging position was not selected as an important trait in any of the models investigated. Moreover, although forest dependence was an important trait separating groups that respond differently to habitat features, it exhibited a strong interaction with other traits, highlighting the fact that forest visitor responses to environmental variables vary as a function of feeding habits.
Different groups responded differently to leaf litter depth. Forest-dwelling species (Generalists and Specialists) responded negatively to leaf litter depth, as did forest visitors that have a mixed diet of fruits and insects. Conversely, forest visitor species who feed only on insects or seeds as well as predators were positively correlated with leaf litter depth. In a separate recent study, we discovered that beetles (primary food for many of the insectivorous bird species) in the Mixed Forest area of ASF were associated with higher leaf litter, habitat that is critically important for arthropods (Banks et al., 2010). In many cases, forest stewardship plans are aimed at effectively regulating understory conditions to bolster endangered or declining species, but may ultimately serve to diminish other species or guilds. The fact that some bird species with decreasing population trends that include insects in their diets are correlated positively with leaf litter (e.g., the Northern Brownbul, Phyllastrephus strepitans [forest visitor]; RDA1 species score = 0.16) while others are correlated negatively (e.g., the Green Barbet, Cryptolybia olivacea [forest specialist]; RDA1 species score = −0.15) highlights the difficulties inherent in prescribing effective “one-size-fits-all” habitat management protocols for wildlife conservation, even within guilds.
Other recent studies in ASF have illustrated that some species respond to a combination of vegetation preference and levels of disturbance (Virani, Njoroge, & Gordon, 2010). Disturbance created by African elephants is a critical driver of ecological processes in sub-Saharan Africa woodlands and savannahs (Baxter & Getz, 2005; Kerley & Landman, 2006; Landman, Kerley, & Schoeman, 2008). A closer look at the responses of individual endangered species in ASF from the current study reveals very disparate responses to elephant disturbance. For instance, the Sokoke Pipit (Anthus sokokensis) was strongly associated with the presence of elephants (RDA score = 0.22), whereas the East Coast Akalat (Sheppardia gunningi) was correlated with the absence of elephants (RDA score = −0.16). Hence, managing both elephants and avian species within a relatively small reserve requires extra vigilance and attention to population dynamics.
The fact that FD decreased with increasing levels of canopy cover suggests that less mature forest or forest exposed to some level of disturbance may boost niche complementarity. Increased canopy cover may also affect understory growth; such indirect effects make it more challenging to predict how different species will respond to environmental characteristics. Overall, our results highlight the difficulty inherent in determining a single species that can serve as a surrogate or an indicator for entire assemblages of species of concern in forest reserves such as ASF. A spate of recent studies (Banks, Stark, Vargas, & Ackleh, 2014; Dale & Beyeler, 2001; Estrada & Rodríguez-Estrella, 2016; Lindenmayer, Cunningham, & Fischer, 2005; Lindenmayer et al., 2006) have demonstrated that taking a surrogate species approach to the conservation of a larger assemblage of species is fraught with difficulties in many ecological settings and across myriad taxa. Although recent research has illustrated promising potential for the use of similar bird species as surrogates in nearby Taita Hills (Githiru et al., 2007), that work was restricted to pairwise species match-ups. Idiosyncratic responses to differences in vegetation condition serve as a foil to simple forest management prescriptions (Lindenmayer et al., 2002; Wiens et al., 2008). Further complicating the choice of suitable surrogates is the phenomenon that the response of rare vs. common species may differ if critical elements are either (a) met everywhere or (b) unmet everywhere (Pearman & Weber, 2007). In both cases, species’ distributions will appear to be invariant across the range of the environmental factor (Lennon, Beale, Reid, Kent, & Pakeman, 2011). Additionally, certain bird species may be more easily detectable than others, which can influence attempts to correlate their distribution with environmental variables—although the importance of detectability remains unclear (Lahoz-Monfort, Guillera-Arroita, & Wintle, 2014; Rota, Fletcher, Evans, & Hutto, 2011). Taken together, these issues cast substantial doubt on our ability to choose suitable surrogate or indicator species that accurately predict the fate of entire species assemblages.
Implications for Conservation
The management challenges faced by ASF are similar to those in many other reserve hot-spots found throughout the tropics, in that forest management is aimed at encompassing protection of not only a large assemblage of bird species but over half a dozen more endangered or threatened birds along with several mammals as well as the African elephant (Loxodonta africana). Available evidence suggests that some of the endangered bird species in ASF show strong preferences for some habitats over others; these preferences likely stem from a combination of vegetation composition and degree of disturbance due to both natural and anthropogenic forces (e.g., poaching; Banks et al., 2012; Otieno et al., 2014). Understanding the roles played by all of the wildlife in such reserves is clearly critical—this includes especially megafauna such as elephants, whose foraging behavior can negatively and positively affect different types of vegetation and wildlife (Banks et al., 2010; Cumming et al., 1997; Guidemond & Van Aarde, 2008; Herremans,1995; Otieno et al., 2014; Vogel et al., 2014). Larsen, Bladt, Balmford, and Rahbek (2012) note that supplementing bird biodiversity data with information about additional taxa greatly improves indicator or surrogate species approaches to conservation. Our data reaffirm that more in-depth studies of the effects of vegetation heterogeneity and individualistic responses of groups of species need to be undertaken in order to establish optimal protocols for wildlife management in forest reserves. In the case of elephant-dominated reserves, accurate census data within reserves is not enough—a better understanding of detailed spatial patterns of habitat use and dispersal is critical to prescribing effective management protocols for the conservation of species of concern across taxa.
Footnotes
Appendix
Acknowledgments
The authors thank Laura Sleeman for help with data organization and editing, and to anonymous reviewers for helpful suggestions.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The authors would like to thank KWS, KFS, and NatureKenya for providing support for this project with a Critical Ecosystem Profile Fund grant. J.E.B. was supported in part by the August T. Larsson Guest Researcher Program at the Swedish University of Agricultural Sciences (SLU) Uppsala during the preparation of this manuscript.