
Abstract
Objectives
Infections caused by multidrug-resistant (MDR) Pseudomonas aeruginosa are a significant public health concern. Imperata cylindrica is a dietary plant used in traditional medicine to treat many health conditions, including microbial infections. This study aimed to evaluate the anti-pseudomonal activity, antibiotic resistance modulation potential, and the effect of dichloromethane/methanol extract of I. cylindrica rhizomes on the antioxidant system of MDR P. aeruginosa.
Methods
The microdilution method was used to assess the anti-pseudomonal and antibiotic resistance modulation activities of the extract. The effect of the extract on catalase activity and lipid peroxidation was evaluated by measuring the height of the foam and spectrophotometric quantification of malondialdehyde, respectively.
Results
The tested extract displayed variable antibacterial activity, with minimum inhibitory concentrations (MICs) ranging from 32 to 2048 µg/mL. Higher anti-pseudomonal activity was observed against P. aeruginosa D130 (MIC = 32 µg/mL) and K290 (MIC = 64 µg/mL). At its sub-inhibitory concentration (MIC/8), I. cylindrica extract enhanced the activity of the tested antibiotics, mainly aminoglycosides (streptomycin, amikacin, and gentamycin), by 2 to 258-fold. The extract also affected the antioxidant system of P. aeruginosa D130, by inhibiting catalase activity, as confirmed by lipid peroxidation.
Conclusion
These findings indicate that the dichloromethane/methanol extract of Imperata cylindrica rhizomes can serve as a potential candidate for the discovery of new antibiotics or antibiotic-resistant modulators to efficiently treat infections involving MDR Pseudomonas aeruginosa.

Introduction
Antimicrobial resistance stands as a significant global health menace of the 21st century, particularly with the emergence of multidrug-resistant (MDR) bacteria. 1 MDR is defined as the non-susceptibility to at least one antimicrobial agent across two or more distinct categories of antimicrobial agents. 2 Among the various challenges associated with AMR, the emergence and rapid dissemination of MDR Gram-negative bacteria, such as MDR Pseudomonas aeruginosa, represent a significant threat to healthcare systems. 1 P. aeruginosa is an opportunistic pathogen associated with various nosocomial and community infections, including chronic respiratory infections, urinary tract infections, meningitis, and bacteraemia. 3 Infections caused by MDR P. aeruginosa are a significant public health issue, with high mortality and morbidity rates, increased resource utilisation, and costs. 4 By means of the efflux systems and the β-lactamases’ expression, P. aeruginosa has developed a high resistance to several antimicrobial drugs, including aminoglycosides, β-lactams, polymyxins, and quinolones.5,6 Countermeasures are crucial since few antibiotics are effective against infections caused by MDR bacteria.
Bacteria, including P. aeruginosa, produce under stress conditions enzymes, such as catalases, which significantly protect them from oxidative stress caused by reactive oxygen species (ROS). 7 Without such a bacterial antioxidant system, ROS induce lipid peroxidation that causes changes in the bacterial membrane structure by altering its fluidity and damaging the membrane integrity. 8 Therefore, molecules targeting the antioxidant system of bacteria might serve as good drug candidates against MDR bacteria.
Medicinal plants have historically played a vital part in traditional use to cure a range of disorders, including bacterial infections. 9 This intensified focus on medicinal plants since they hold an immense reservoir of bioactive compounds that could lead to the discovery of novel antibacterial drugs10,11 and antibiotic resistance modulators.12,13 Imperata cylindrica, a plant of the Gramineae family, is a perennial rhizomatous plant that can grow on soils with a vast range of nutrients and moisture. 14 It is a plant widely distributed in southwestern Asia and is specifically native to the tropical and subtropical zones. I. cylindrica is a medicinal spice used in traditional medicine either alone or in combination with other herbal medicines to cure haematuria, jaundice caused by damp-heat pathogen invasion, emesis, oedema, haemorrhage, fever, pain, polydipsia, diarrhoea, and urinary tract infections associated with heat toxins.15-17 Besides, the I. cylindrica rhizome has high nutritional value. It contains crude fiber, carbohydrates, sugars, fatty acids, and trace elements, indicating its low-calorie and health-promoting properties. 16 The main phytochemicals isolated in I. cylindrica include terpenoids, saponins, coumarins, flavonoids, and phenols.16,17 These constituents contribute its pharmacological properties, including immunomodulatory, 17 antioxidant, 18 antibacterial,19,20 antifungal, 21 antihypertensive, 22 hepatoprotective, 23 anticancer,24,25 anti-inflammatory, 26 anti-helminthic, 27 and anti-trypanosomal 28 properties. Although the antimicrobial activity of some extracts from I. cylindrica has been previously evaluated against various pathogens, there is little information regarding their antibacterial property, antibiotic resistance modulation activity, and modes of action against multidrug-resistant P. aeruginosa. Based on these findings, we hypothesized that the dichloromethane/methanol extract of Imperata cylindrica rhizomes may exhibit anti-pseudomonal activity by inhibiting the antioxidant system and modulating antibiotic resistance in MDR P. aeruginosa. This study aimed to investigate the antipseudomonal activity, antibiotic resistance modulation potential, and effect of the dichloromethane/methanol extract of Imperata cylindrica rhizomes on the antioxidant system of MDR P. aeruginosa.
Materials and Methods
Plant Collection and Identification
Fresh Imperata cylindrica rhizomes were collected in November 2023 at Dschang, West Cameroon. The plant sample was identified at the National Herbarium of Cameroon, Yaoundé, where the voucher has been deposited under registration number 10084/SRF/Cam.
Preparation of I. cylindrica Extract
The dried Imperata cylindrica rhizomes were ground, and the obtained powder (200 mg) was macerated for 48 hours at room temperature (19-26°C) in 600 mL of a mixture of dichloromethane/methanol (1:1, v/v), with constant agitation. The macerated powder was filtered using Whatman filter paper No. 1, and the filtrate was concentrated under reduced pressure at 40°C using a rotary evaporator (BÜCHI R-200). The residual solvent was removed by drying the crude extract at 40°C in an oven. Finally, the prepared extract was kept at 4°C in a sealed container until further use.12,29 The extraction yield, calculated as the ratio of the extract weight to that of the dried plant material, was 4.73%.
Test Microorganisms and Their Culture Conditions
The test organisms consisted of 19 P. aeruginosa strains, including one strain from the American Type Culture Collection (ATCC27853) and 18 MDR clinical isolates (B026, B178, D100, D130, K033, K126, K130, K139, K142, K261, K272, K290, K294, T076, T106, T138, T201, and PA124). Their resistance profiles are presented in the supporting information (Table S, Supplemental material). These organisms were cultured and maintained at 37°C for 18 h in Petri dishes containing Mueller-Hinton agar (MHA, ReadyMED®, USA).
Inoculum Preparation
A bacterial suspension equivalent to 0.5 McFarland standard (1.5 × 108 CFU/mL) was prepared from a pure culture of the test organism. Briefly, 4-6 colonies of P. aeruginosa were mixed with sterile distilled water to obtain a suspension with an absorbance of 0.08–0.1 at 600 nm using a UV/VIS spectrophotometer (Biobase BK-D590). Thereafter, the obtained suspension was diluted with Mueller-Hinton broth (MHB, ReadyMED®, USA) in 10 mL sterile test tubes to obtain a concentration of 2×106 CFU/mL.30,31
Determination of the Anti-pseudomonal Activity of I. cylindrica Extract
The minimum inhibitory concentration (MIC) of the samples was determined by the microdilution method using iodonitrotetrazolium chloride (INT) as previously described.29,32 The test sample solution was prepared by dissolving samples in dimethyl-sulfoxide (DMSO, BDH Chemicals Ltd, Poole, England), followed by dilution in MHB to yield stock solutions of 4096 µg/mL for the extract and 1024 µg/mL for streptomycin used as reference antibacterial. A volume of 100 µL of each solution was serially diluted twice in a 96-well microplate containing 100 µL of MHB. Thereafter, 100 µL of inoculum (2×106 CFU/mL) was added to each well. Wells containing 2.5% DMSO served as the negative control. After 18 h of incubation at 37°C, the MIC of each sample was detected following the addition of 40 µL of INT solution (0.2 mg/mL) and incubation at 37°C for 30 min. MIC was defined as the lowest concentration of the sample that completely prevented bacterial growth (absence of pink color in the wells). The minimum bactericidal concentration (MBC) of each sample was determined by sub-culturing 50 μL of the bacteria suspension from the wells that did not show any growth after the MIC assay in a sterile 96-well microplate containing 150 μL of fresh MHB. Thereafter, the microplate was incubated at 37°C for 48 h. The lowest concentration of the sample that prevented the formation of the pink colour after the addition of INT, as mentioned above, was identified as the MBC. 29 Each sample underwent two tests, and the experiment was conducted three times.
The anti-pseudomonal activity of the extract was defined as follows: outstanding activity (MIC ≤ 32 µg/mL), excellent activity (32 < MIC ≤ 128 µg/mL), very good activity (128 < MIC ≤ 256 µg/mL), good activity (256 < MIC ≤ 512 µg/mL), average activity (512 < MIC ≤ 1024 µg/mL), weak activity or not active (MIC > 1024 μg/mL). 33
Evaluation of the Antibiotic Resistance Modulation Activity of the Extract
The antibiotic resistance modulation activity of the extract was performed as previously described.34,35 Briefly, the MIC values of amikacin, cefixime, cefotaxime, ciprofloxacin, gentamicin, levofloxacin, and streptomycin (Sigma-Aldrich) were determined in the presence or absence of the different extracts at their sub-inhibitory concentrations against six MDR clinical isolates, including B026, K033, K126, K294, T201, and PA124. After a preliminary assay with different sub-inhibitory concentrations of extract against PA124, the extract at MIC/8 was selected and tested for its antibiotic resistance modulation potential on other MDR isolates. Each antibiotic was subjected to serial dilution as described above, and each extract solution at sub-inhibitory concentrations was introduced into the wells. Following inoculation and incubation, as outlined above, the modulation factor was calculated as the ratio of the MIC of the antibiotic alone to that of the antibiotic in the presence of the extract. Columns that did not receive the extract were used to determine the MIC of the antibiotic, whereas those containing the extract were used to determine the MIC of the combination. A modulation factor ≥ 2 has been set as the threshold for the biological significance of the extract’s antibiotic resistance modulation activity. 36 Each assay was done in duplicate and repeated three times.
Evaluation of the Effects of the Extract on the Antioxidant System of MDR P. aeruginosa
Catalase Assay
The effect of the extract on the catalase activity of P. aeruginosa D130 was determined as previously described.37,38 Briefly, bacterial colonies were inoculated into 5 mL of MHB and incubated overnight at 37°C. Thereafter, 4 mL of the inoculum was cultivated with 1 mL of extract (MIC and 2xMIC) or ciprofloxacin (MIC), which was used as the reference antibacterial drug. Next, 100 µL of each sample aliquot was added to a test tube along with 100 µL of 30% (v/v) H2O2 and 100 µL of 1% Triton X-100. Each tube was kept at room temperature for 5 min after mixing, and the height of the foam (cm) was measured using a ruler. The catalase inhibition effect was based on the height of foam (cm) in the test tubes compared to that in the control. Tubes containing distilled water instead of the samples were used as controls. The assay was performed in triplicate.
Lipid Peroxidation Assay
The lipid peroxidation activity of the extract was assessed by measuring the concentration of malondialdehyde (MDA).
39
Briefly, fresh colonies of P. aeruginosa D130 were cultured overnight in 4 mL of MHB containing 1 mL of extract (MIC and 2xMIC) or polymyxin B (MIC). Subsequently, 1 mL of the test culture and 1 mL of trichloroacetic acid were introduced into a screw-cap test tube, mixed, and heated for 10 min at 100°C in a water bath. After cooling, the tubes were centrifuged for 20 min at 5000 rpm. The supernatants were collected, and their absorbance was measured at 535 nm against the control. The tubes without extract served as controls. The MDA concentration (µM) was calculated based on its millimolar absorbance coefficient (E0 = 156 cm−1. mM−1) using the following equation:

Data Analysis
Data were reported as mean ± standard deviation (mean ± SD) of three replicates. One-way analysis of variance (ANOVA), followed by Tukey’s post hoc multiple comparison test at a significance level of p < 0.05, was used to compare the means. GraphPad Prism software for Windows, version 8.4.2, was used for data analysis.
Results
Anti-Pseudomonal Activity of I. cylindrica Extract
MIC and MBC (µg/mL) of CH2Cl2/MeOH Extract of I. cylindrica Rhizomes and Streptomycin Against P. aeruginosa

MIC: minimal inhibitory concentration; MBC: minimal bactericidal concentration; values in bold represent outstanding (MIC ≤ 32 µg/mL) and excellent (32 < MIC ≤ 128 µg/mL) activities of the extract. 33
Antibiotic Resistance Modulation Activity of I. cylindrica Extract
Antibiotic Resistance Modulation Effect of CH2Cl2/MeOH Rhizome Extract of I. cylindrica Against MDR P. aeruginosa
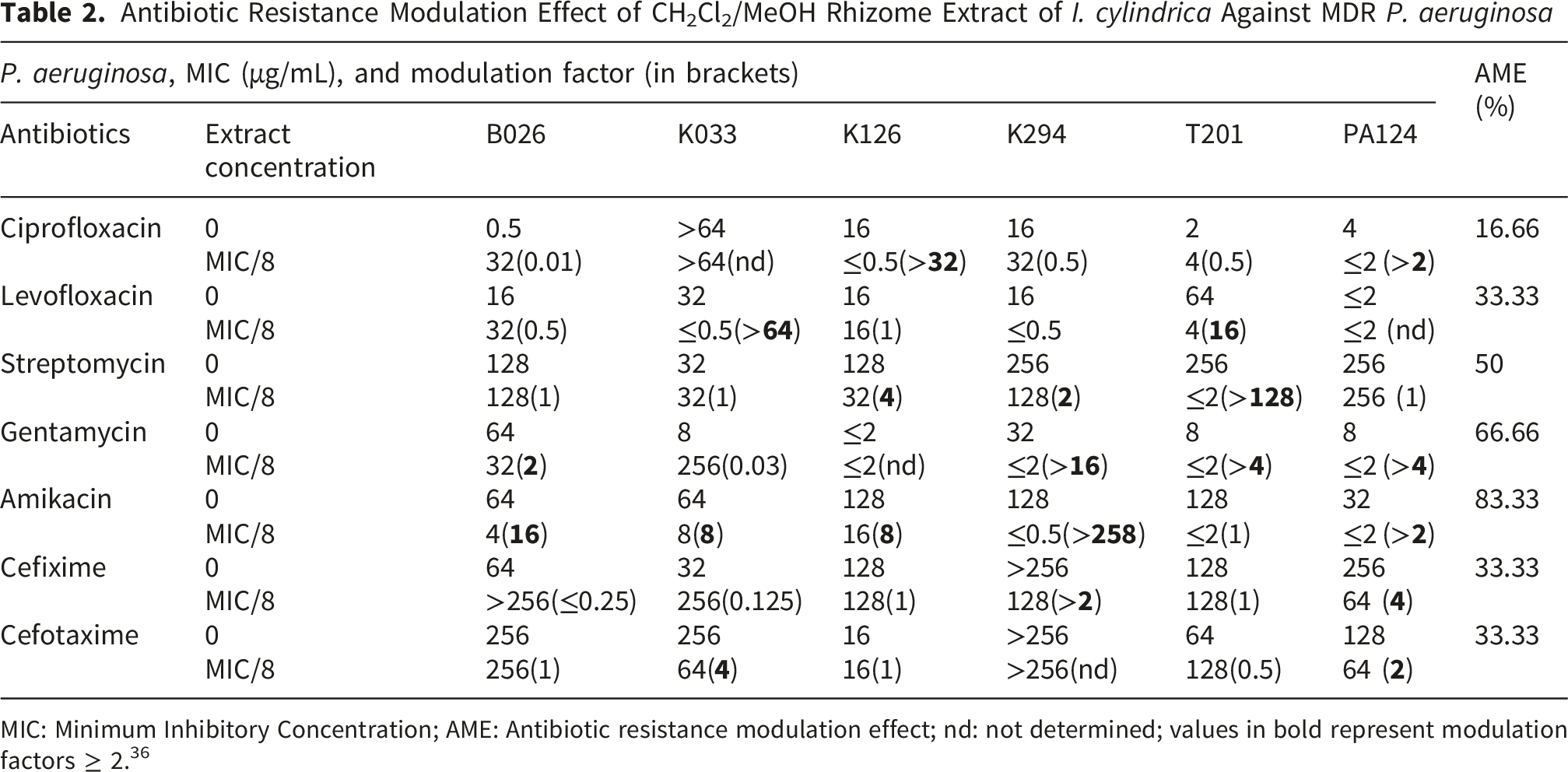
MIC: Minimum Inhibitory Concentration; AME: Antibiotic resistance modulation effect; nd: not determined; values in bold represent modulation factors ≥ 2. 36
Effect of I. cylindrica Extract on the Antioxidant System of P. aeruginosa
Effect of the Extract on Catalase Activity
In this study, we measured the impact of the dichloromethane/methanol rhizome extract of I. cylindrica on the catalase activity in P. aeruginosa D130. The extract (MIC and 2×MIC) significantly (p < 0.05) reduced catalase activity in P. aeruginosa D130 compared to that of the control (Figure 1). However, at 2×MIC, it had a more substantial effect, resulting in a foam height of 0.45 ± 0.07 cm. Ciprofloxacin was more active than the extract (0.35 ± 0.07 cm). Effect of the extract on catalase activity of P. aeruginosa D130. The MIC values of ciprofloxacin and the extract against P. aeruginosa D130 were 16 and 32 µg/mL, respectively. ** (p<0.01). IC: Imperata cylindrica; MIC: Minimal inhibitory concentration
Effect of the Extract on Lipid Peroxidation
Exposure of P. aeruginosa D130 isolate to the extract resulted in a significant (p < 0.05) and concentration-dependent increase in lipid peroxidation, resulting in high production of malondialdehyde (Figure 2). At 2×MIC, this extract resulted in a considerable increase in MAD concentration (0.934 ± 0.07 µM) compared to the control (0.339 ± 0.055 µM) and polymyxin B (0.574 ± 0.023 µM). Effect of the extract on lipid peroxidation in P. aeruginosa D130. The MIC value of polymyxin B and the extract against P. aeruginosa D130 was 32 µg/mL. IC: Imperata cylindrica; MIC: Minimal inhibitory concentration. * (p<0.05); ** (p<0.01); *** (p<0.001)
Discussion
Multidrug-resistance is responsible for treatment failure of many diseases, including infections caused by MDR P. aeruginosa.1,5 Therefore, the discovery of new treatment options for infections caused by MDR pathogens must be envisaged. Many plant extracts, including those from edible plants, have been shown to have antibacterial and antibiotic resistance modulatory activity against drug-resistant bacteria, including MDR P. aeruginosa.20,38,40,41 As part of our ongoing research, this study aimed to investigate the anti-pseudomonal activity, antibiotic resistance modulation potential, and effect of the dichloromethane/methanol extract of I. cylindrica rhizomes on the antioxidant system of multidrug-resistant P. aeruginosa.
This study showed that the dichloromethane/methanol rhizome extract of I. cylindrica displayed varying anti-pseudomonal activity. According to the MIC cut-off points established by Tankeo and Kuete (2023), the extract showed outstanding (MIC ≤ 32 µg/mL) to average (512 < MIC ≤ 1024 µg/mL) anti-pseudomonal activity against the test pathogens. 33 Several studies have reported the antimicrobial effects of extracts from the roots, leaves, and whole plant of I. cylindrica against many sensitive and resistant microorganisms. For instance, Voukeng et al (2012) have demonstrated the in vitro antibacterial activities of the methanol root extract of I. cylindrica against bacteria, including Enterobacter aerogenes, Enterobacter cloacae, Escherichia coli, Klebsiella pneumoniae, Providencia stuartii, and P. aeruginosa. 20 Moreover, a study had shown the in vitro anti-staphylococcal activity of the methanol root extract of this plant against a panel of methicillin-resistant (MRSA) and methicillin-susceptible Staphylococcus aureus (MSSA), with MIC values ranging from 512 to 2048 µg/mL. 42 Many other studies have shown the in vitro antibacterial activity of I. cylindrica against many other bacterial strains (Bacillus spp., Proteus spp., Salmonella spp., and Listeria monocytogenes) through the determination of the diameter of inhibition zones.43-46 The observed antibacterial activity may be due to the presence of secondary metabolites identified in I. cylindrica, such as terpenoids, flavonoids, and phenols, 17 known for their antimicrobial activity. 10 Some metabolites isolated from the root of I. cylindrica (cylindraucin, cylindricin A, cylindricin B, cylindracide A, 3′,4′, 5,5′,7- pentahydroxyflavanone, and mearnsetin) have been shown to have considerable antibacterial activities. 47 The findings of this study highlighted the antibacterial properties of I. cylindrica, particularly those of its dichloromethane/methanol rhizome extract.
Combination therapies are considered a promising approach to combat antibiotic-resistant bacteria. 48 For instance, it has been shown that natural substances, such as alkaloids and flavonoids, might enhance the activity of antibiotics when used in combination with them.49,50 In this study, I. cylindrica extract (at MIC/8) enhanced the activity of antibiotics, mainly aminoglycosides (amikacin, gentamicin, and streptomycin), by 2- to 258-fold. Given that these antibiotics act at the intracellular level by inhibiting protein synthesis and that efflux pumps represent the main resistance mechanism in P. aeruginosa, this finding suggests that the constituents of this extract may act as efflux pump inhibitors.51,52 This resistance mechanism is significantly involved in the aminoglycoside resistance in MDR P. aeruginosa. 52 Furthermore, the observed antibiotic modulation effect could be due to the action of the extract on different bacterial targets than those of antibiotics. This may include disruption of the bacterial membrane, thus facilitating the entry and action of these antibiotics. In a previous study, the methanol extract of I. cylindrica was shown to potentiate the activity of tetracycline, chloramphenicol, ciprofloxacin, and ceftriaxone by 2- to 16-fold in selected Gram-negative bacteria. 20 Therefore, our findings with the dichloromethane/methanol extract of I. cylindrica rhizomes highlight the potential of I. cylindrica extract as a source of antibiotic enhancers against MDR P. aeruginosa.
The development of antibacterial therapies that target previously unexpected pathways may be useful in the search for new antibacterial drugs. Enzyme antioxidant systems, such as catalase, are used by bacteria to protect themselves from ROS, which can cause oxidative stress through lipid peroxidation. 7 By blocking this enzyme, ROS will induce lipid peroxidation, which can cause cell death by altering the membrane integrity of the bacteria. 8 In this study, we evaluated the effect of the extract on the antioxidant system (catalase activity and lipid peroxidation) of P. aeruginosa. The extract affected the antioxidant system of P. aeruginosa D130 by inhibiting catalase activity, as confirmed by the observed lipid peroxidation effect. This could be due to the presence of phytochemicals such as phenols, whose antibacterial activity is attributed to their interaction with the bacterial cell wall and cell membrane, the generation of ROS, the inhibition of DNA replication, and protein denaturation, affecting enzymatic activity.37,53 The isolation of such compounds in I. cylindrica root extract can explain the observed action. 54 The induction of ROS by the extract may also affect its antibiotic resistance modulation effect. In fact, it has been demonstrated that substances that enhance ROS production or inhibit bacterial oxidative stress hold potential in overcoming antibiotic resistance and enhancing antimicrobial efficacy.55-57
This study is subject to several limitations, including the need for in-depth mechanistic studies, validation in both in vivo and clinical settings, and identification of bioactive constituents.
Conclusions
The findings of this study provide valuable insights into the potential of the dichloromethane/methanol extract of I. cylindrica rhizomes in combating infections caused by MDR P. aeruginosa. They also provide scientific credence to the traditional use of I. cylindrica for managing diseases, including bacterial infections. Finally, this study provides the first evidence of the antibiotic resistance modulation activity and inhibition of the antioxidant system of the dichloromethane/methanol extract of I. cylindrica against MDR P. aeruginosa. Therefore, to facilitate its clinical use, its safety and the identification of its active ingredients must be investigated.
Supplemental Material
Supplemental material - Anti-Pseudomonal Activity, Antibiotic Resistance Modulation Potential, and Effect of Dichloromethane/Methanol Extract of Imperata cylindrica (Gramineae) on the Antioxidant System of Multidrug-Resistant Pseudomonas aeruginosa
Supplemental material for Anti-Pseudomonal Activity, Antibiotic Resistance Modulation Potential, and Effect of Dichloromethane/Methanol Extract of Imperata cylindrica (Gramineae) on the Antioxidant System of Multidrug-Resistant Pseudomonas aeruginosa by Aimé Gabriel Fankam, Céline Brinda Sonfack, Brenda Ngueffo Tiwa, Victor Kuete in Natural Product Communications.
Footnotes
Acknowledgments
The authors thank the National Herbarium of Cameroon, Yaoundé, Cameroon, for the identification of the studied plant.
Author Contributions
AGF: Conceptualization, Methodology, Resources, Supervision, Validation, Data curation, Formal analysis, Writing – original draft, Writing – review & editing. CBS: Investigation, Resources, Data curation. BNT: Investigation, Writing – review & editing. VK: Critical review & editing. All authors approved the final manuscript.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
The datasets generated during and/or analyzed during the current study are available from the corresponding author on request.
Statement of Human and Animal Rights
This article does not contain any studies with human or animal subjects.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.