
Abstract
Objective
Lamiaceae plants have high aromatic and medicinal qualities. This work reports the chemical composition of essential oils (EOs) from Thymus capitatus, T. fontanesii and Lavandula stoechas together with their inhibitory effects of selected enzymes and microbial virulence factors.
Methods
EOs were obtained by hydrodistillation and their chemical compositions determined by GC-FID/GC-MS. The enzyme inhibition of EOs on acetylcholinesterase (AChE), butyrylcholinesterase (BChE), α-amylase, α-glucosidase, urease and tyrosinase were investigated alongside antimicrobial, antibiofilm and quorum-sensing inhibitions (QSI) against a range of bacteria and fungi.
Results
In EOs from T. capitatus, T. fontanesii and L. stoechas, 36 (100%), 27 (100%) and 42 (100%) compounds were identified respectively. High amounts of oxygenated monoterpenoids were identified in T. capitatus (83.11%), T. fontanesii (72.77%) and L. stoechas (96.53%). At 200 µg/mL, L. stoechas EO exhibits the highest anticholinesterase activity of 56.63 ± 0.79% (AChE) and 71.45 ± 0.37% (BChE), compared to galantamine with 92.22 ± 0.18% (AChE) and 80.37 ± 0.27% (BChE). At 200 µg/mL, T. capitatus EO exhibits good inhibition of α-glucosidase (64.89 ± 0.82%) and α-amylase (62.44 ± 0.78%) compared to acarbose with inhibitions of 67.30 ± 0.45% (α-glucosidase) and 80.56 ± 0.52% (α-amylase). Urease and tyrosinase inhibitions were low. The EOs demonstrated strong antimicrobial activities with inhibition diameters reaching 45.0 ± 0.5 mm and minimal inhibitory concentrations (MIC) as low as 0.007 ± 0.32% v/v. At MIC and sub-MIC concentrations, the EOs inhibited biofilm formation in a concentration-dependent manner. Gram-positive and candida biofilms were relatively more susceptible. EOs inhibited violacein production against C. violaceum CV12472, with T. fontanesii EO being most potent. QSI zones against C. violaceum CV026 were large and L. stoechas EO was most active. EOs inhibited swimming and swarming motilities against P. aeruginosa PA01.
Conclusion
T. capitatus, T. fontanesii, and L. stoechas are rich in bioactive EOs which can be used to mitigate enzymes related ailments such as diabetes and Alzheimer's disease and attenuate virulence factors in pathogenic bacteria.
Keywords
Introduction
As defined in ethnopharmacology, traditional or herbal medicine involves the use of plant species for therapeutic purposes in humans and/or animals. Despite the advancements of modern medicine, there is universal persistence in the use of medicinal plants in supporting primary healthcare, especially where access to conventional healthcare is limited. 1 Approximately 14% to 28% of higher plant species have therapeutic uses based on ethnomedical practices and over 74% of bioactive compounds have been discovered from them. 2 The WHO recognizes the importance of medicinal plants and have documented over 20 000 species to date, demonstrating that over 80% of the world's population depend on them for primary healthcare. 3 Research on medicinal plants is multidisciplinary and spans across botany, plant biology, phytochemistry and pharmacology so as to ensure the quality, efficacy and safety of herbal medicines. 4 Plant extracts, essential oils and phytochemicals play a crucial role in therapeutic treatments, harnessing the antimicrobial properties of many plants.5‐9 Recent studies include the validation of the effectiveness of traditional medicines based on suitable methods of extraction, isolation of bioactive compounds, essential oils and bioassays. 10
Natural products are used to remedy a range of ailments that results from microbial infections and inappropriate enzymes expression. Neurological disorders like Alzheimer's diseases occur as a result of low levels of acetylcholine neurotransmitter, which is hydrolyzed by acetylcholinesterase (AChE) and butyrylcholinesterase (BChE) and leading to dementia and death in some cases. 11 Diabetes another leading metabolic diseases occurs from high amounts of blood glucose levels leading to hyperglycemia. Type 2 diabetes occurs when the body does not make enough insulin or the insulin produced does not work properly and can be cured by inhibiting the enzymes α-glucosidase and α-amylase which breakdown polysaccharides to produce absorbable sugars into the blood stream, causing postprandial glycaemia and advanced glycation end products.12‐14 Tyrosinase and urease are implicated in skin hyperpigmentation and gastric ulcers respectively, and their inhibition, particularly by natural products, is being explored as a potential treatment to lower the incidence of skin cancer and skin spots in the case of tyrosinase and to lower the infections from ureolytic bacteria like helicobacter in the case of urease.15,16 Antibiotics that aim to kill bacteria are becoming less effective over time as a result of resistance especially from biofilm colonies whose infections persists and cause surface contamination, nosocomial infections, deadly food poisoning, food deterioration, and other numerous health risks. 17 Antibiotic resistance relies on a number of virulence factors, including bacterial biofilms, swarming motility, and cellular quorum-sensing (QS) activities.18,19 Through the use of a network of cell-to-cell communication, QS allows bacterial colonies to monitor their environment and regulate the production of signal molecules including violacein, toxin, and motility during swarming. 20 In response to the continuous search of alternative medicines for microbial virulence and enzymes-related sicknesses, essential oils are being investigated. Essential oils are rich in low molecular weight compounds mostly terpenoids and phenylpropanoids with medicinal properties including antidiabetic and antimicrobial effects.21,22 This shows the great potential of essential oils as aromatherapeutic agents, capable of addressing problems linked to Alzheimer's disease, diabetes, hyperpigmentation, melanogenesis, ureolytic bacterial infections, microbial virulence and resistance.
Lamiaceae plants (mints or sage plants) are one of the most important sources of essential oils. They are consumable, ornamental and medicinal herbs which are easy to cultivate. They are among the over 3164 plant species within Algeria's floristic diversity, ranging from the Mediterranean in the north to the Sahara in the south. 23 The Lamiaceae in Algeria comprises 183 taxa, of which 19 are endemic, representing a total of 28 genera and 146 species.24,25 Species from this family are used in both traditional and modern medicine. 26 The genus Thymus is considered one of the prominent genera within the Lamiaceae family, encompassing approximately 215 species that have evolved in the Mediterranean basin. 27 Thymus plants have considerable economic importance and are used as aroma in cosmetics or antimicrobial agents and preservatives in food products.28,29 The examples that occur in Algeria include Thymus algeriensis Boiss. & Reut, T. capitatus Hoffmanns. & Link, T. ciliatus Lam., T. commutatus Batt., T. dreatensis Batt., T. glandulosus Reg., T. guyonii de Noé, T. hirtus Banks. & Sol., T. lanceolatus Desf., T. numidicus Poir., T. pallidus Coss. ex Batt., and T. fontanesii Boiss. & Reut.25,30 Several studies have established a link between the chemical composition of Thymus essential oils and their antimicrobial activities.31,32 In Algeria, Thymus species are widely used several medicinal effects. 33 Lavandula, another widely distributed genus in Algeria, comprises of six species namely; L. stoechas L., L. multifida L., L. coronopifolia Poir., L. pubescens Dec., L. dentata L., and the recently added L. antineae Maire.34,35 Lavandula is a rich source of various secondary metabolites, including essential oils, coumarins, and phenolic compounds. 36 It is commonly used in the treatment of superficial wounds and burns, exhibiting sedative, antibacterial, antifungal, antidepressant, and anti-inflammatory properties. 37 The plants selected for this study are T. capitatus, T. fontanesii and L. stoechas which are used in Algerian folk medicine as antiseptic and stimulating agents. 38 Additionally, in Algerian cuisine, these plants are used in herbal teas, couscous as well as in flavoring and sanitizing meats.35,39
In the context of our ongoing efforts to characterize and valorize Algerian aromatic and medicinal plants, this work is focused on the comprehensive study of chemical composition of selected Lamiaceae plants from Algerian flora and evaluation of their inhibitory effects on certain enzymes such as acetylcholinesterase (AChE), butyrylcholinesterase (BChE), α-amylase, α-glucosidase, urease and tyrosinase. The antimicrobial effects of the essential oils are also reported as well as their ability to disrupt biofilm formation and quorum-sensing in bacteria.
Materials and Methods
Plant Material
Three plants (T. capitatus, T. fontanesii; L. stoechas L.) belonging to the Lamiaceae family, renowned for their therapeutic properties, were collected and investigated. The aerial parts of these plants were harvested at the full flowering stage, between March and June 2022. They were collected from wild populations located in different regions of Tlemcen.
Preparation of Essential Oils
The extraction of essential oils (EOs) was carried out following the guidelines of the European Pharmacopoeia through hydrodistillation using a Clevenger-type apparatus.
40
Briefly, 250 g of each pulverized plant material were separately mixed with 800 mL of distilled water and boiled for 3 h. The water vapour loaded with the essential oil was condensed and separated using a separatory funnel. The distillation process was performed three times for each plant and the essential oils combined and dried using anhydrous magnesium sulphate. The obtained essential oils were stored at a temperature of 4 °C, in the absence of light. The yield (%) of each essential oil was determined using the following formula below:
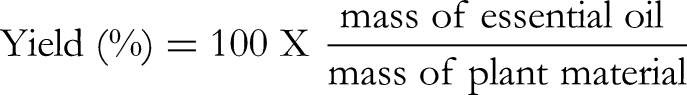
Characterization of Essential Oils
The chemical composition of the EOs were characterized using Gas chromatography-flame ionization detector (GC-FID) and gas chromatography-mass spectrometry (GC-MS) analyses, conducted according to established methods.41,42 GC analyses utilized a Rxi-5Sil MS fused silica capillary non-polar column (30 m × 0.25 I.D., film thickness 0.25 µm) and FID. The injector and detector temperatures were set to 250 °C and 270 °C respectively. Helium served as the carrier gas with a flow rate of 1.4 mL/min, and a sample injection volume of 0.2 µL with a split ratio of 20:1 was used. Percentage composition of the essential oils was determined using a Class GC10 GC computer program. The initial oven temperature was maintained at 60 °C for 5 min, followed by a gradual increase to 240 °C at a rate of 4 °C/min, and then held at this temperature for 10 min.
For GC-MS analyses, an ion trap MS and a Rxi-5Sil MS fused silica non-polar capillary column (30 m × 0.25 mm I.D., film thickness 0.25 µm) were employed. Helium was used as the carrier gas at a flow rate of 1.4 mL/min. Injector and MS transfer line temperatures were set at 220 °C and 290 °C, respectively. The ion source temperature was maintained at 200 °C, and a sample injection volume of 0.2 µL with a split ratio of 20:1 was utilized. Electronic ionization-mass spectrometry (EIMS) measurements were performed at 70 eV ionization energy, with a mass range from m/z 28 to 650 amu and a scan time of 0.5 s with 0.1 s interscan delays. The oven temperature program was similar as that of the GC analyses. Identification of EOs components was achieved through co-injection with standards where possible, along with GC retention indices determined using a homologous series of C7-C30 alkanes (Supelco), and computer matching with the Wiley, TRLIB, and NIST08 libraries. Additionally, comparison of fragmentation patterns documented in the literature was conducted. Quantification of individual constituents of the essential oils was based on internal normalization.
Anticholinesterase Activity
The inhibitory activity of acetylcholinesterase (AChE) and butyrylcholinesterase (BChE) was assessed spectrophotometrically using Ellman's method, with some minor modifications.43,44 For this, AChE from electric eel or BChE from horse serum were used, using the substrates acetylthiocholine iodide and butyrylthiocholine chloride respectively. Cholinesterase activity was measured using 5,5′-dithio-bis(2-nitrobenzoic acid) (DTNB). Galantamine was used as the reference compound in these tests. Each experiments were done in triplicates and optical densities were read on a 96-well microplate reader (SpectraMax, Molecular Devices). Results are expressed as percentage inhibition at highest test concentration of 200 μg/mL of EO.
Antidiabetic (α-Amylase and α-Glucosidase Inhibition) Assay
The α-amylase inhibitory activity was evaluated using starch-iodine method with some modifications.45,46 50 µL of α-amylase from porcine pancreas in pH 6.9 phosphate buffer prepared with 6 mM NaCl and 25 µL of EOs solutions were mixed in a 96-well microplate. The mixture was pre-incubated for 10 min at 37 °C. Then, 50 µL of starch solution (0.05%) was added and incubated for 10 min. at 37 °C. Following incubation, the reaction was completed by adding HCl (0.1 M, 25 µL) and Lugol (100 µL) solutions, and the absorbance was recorded at 630 nm. Acarbose was used as standard and each experiment was repeated three times and results expressed as percentage inhibition at 200 μg/mL of EO.
The α-glucosidase inhibitory activity was evaluated according to the method described previously.46,47 50 µL of glutathione, 50 µL of EO solution, 50 µL of α-glucosidase from Saccharomyces cerevisiae in phosphate buffer (0.01 M pH 6.8) and 50 µL of PNPG (4-N-nitrophenyl-α-D-glucopyranoside) in phosphate buffer (0.01 M pH 6.8) were mixed in a 96-well microplate. Then the solution was incubated for 15 min. at 37 °C. The reaction was then stopped with the addition 50 μL of sodium carbonate (0.2 M) and absorbances were read at 400 nm. Acarbose was used as standard compound and each experiment was repeated three times. Results were given as percentage inhibition (%) at 200 µg/mL EO.
Anti-Urease Activity
The anti-urease activity of EOs was determined by the indophenol method by measuring ammonia production. 48 A mixture containing sodium phosphate buffer (100 mM, pH 8.2), urease enzyme (25 μL) and urea (100 mM, 50 μL) was incubated at 30 °C for 15 min with 10 μL of each EO. After incubation, phenol and alkaline reagents were added, and absorbance was measured at 630 nm after further incubation. Thiourea was used as standard and each experiment was done three times. Results are expressed as percentage inhibition at a concentration of 200 μg/mL of EO, compared with standard thiourea.
Anti-Tyrosinase Activity
Anti-tyrosinase activity was assessed as described by. 41 Briefly, after 10 min incubation at 37 °C of a mixture containing 150 μL of sodium phosphate buffer (pH 6.8, 100 mM), 10 μL of EO solution and 20 μL of tyrosinase solution in buffer; 20 µL of L-DOPA was added. Kojic acid was used as a reference compound. Absorbances were read at 475 nm at 37 °C in a 96-well microplate. Each experiment was done in triplicate. Kojic acid was used as standard compound and results are expressed as percentage inhibition at a concentration of 200 μg/mL of EO.
Tested Microbial Strains
The antimicrobial and antibiofilm activity of essential oils was evaluated against six strains of various types of bacteria; including standard Gram-positive bacteria: Staphylococcus aureus (ATCC 25923), Bacillus subtilis (ATCC 6051), Bacillus cereus (ATCC 11778), Enterococcus faecalis (ATCC 29212), and Gram-negative bacteria: Escherichia coli (ATCC 25922), Klebsiella pneumoniae (ATCC 70603), Pseudomonas aeruginosa (ATCC 25853), P. aeruginosa Pa01 as well as a standard yeast: Candida albicans (ATCC 10231). Chromobacterium violaceum CV12472 and Chromobacterium violaceum CV026 were used for the violacein inhibition assay and quorum-sensing inhibition respectively. Pseudomonas aeruginosa PA01 was used in the swimming and swarming motility tests. The microorganisms were provided by two sources: Laboratory of Applied Microbiology in Food, Biomedical and Environment (LAMAABE) in Tlemcen, Algeria, and Food Quality Control and Analysis Program, Ula Ali Kocman Vocational School, Mugla Sitki Kocman University, Ula Mugla, Turkey.
Disc Diffusion Method
Inhibition diameter zones were measured using the disc diffusion method. Microbial strains were incubated at 37 °C in Müller-Hinton broth MH (Fluka Bio Chemika, Spain) for 24 h. Densities were adjusted to an optical density range of 0.08 to 0.13, considered the equivalence of 108 CFU/mL, representing 0.5 McFarland standard. Subsequently, these standardized bacterial suspensions were streaked onto Müller-Hinton agar plates (Fluka BioChemika, Spain) using standardized swabbing techniques. A sterile Whatman No. 03 paper disk, 6 mm in diameter, saturated with 10 μL of pure EO, was then positioned on the agar surface. 49 After a 24-h incubation period, the diameters of inhibition zones around the disks were meticulously measured in millimeters using a calibrated ruler. Each EO was subjected to individual testing, and the experimental procedure was repeated three times to validate result consistency.
Determination of the Minimal Inhibitory Concentration (MIC)
MIC was determined using microdilution method with 96-well microtitration plates, as described previously,50,51 with some adjustments. The MIC was defined as the lowest concentration of EO not producing visible bacterial growth. The stock solution of the EO was prepared with a concentration of 40% v/v. Variable concentrations of the EO were obtained through successive dilutions in MH broth, with the addition of 1% (v/v) Tween 80 to ensure complete dispersion of the essential oils. A complementary solution containing MH broth and 1% Tween 80 was prepared to maintain a constant concentration of 1% Tween 80 in the different concentrations of the EO. The bacteria, initially at a concentration of 108 CFU/mL, were diluted to achieve a final concentration of 105 CFU/mL. In the microplate, 180 μL of the diluted bacterial suspension were placed in the wells. Then, 20 μL of the EO solution were added. The final concentration of Tween 80 was maintained at 0.1% (v/v) in each well, with essential oil concentrations ranging from 4% to 0.0078%. This methodical approach allowed for a precise evaluation of the MIC for different bacterial strains.
Determination of Biofilm Inhibition
The ability of the EOs at MIC and sub-MIC concentrations to inhibit biofilm of microorganisms were evaluated using a microplate biofilm assay.
52
Briefly, 1% of overnight grown cultures of isolates were added into 200 μL of fresh Tryptose-Soy Broth (TSB) supplemented with 0.25% glucose and grown with or without EO for 48 h at 37 °C. The planktonic colonies were gently removed and wells rinsed with distilled water. Biofilm colonies on walls of the wells were stained by filling wells with 200 μL of 0.1% crystal violet solution and allowing for 10 min. The unabsorbed crystal violet were emptied from the wells and 200 μL of 33% glacial acetic acid (for Gram positive bacteria) or ethanol 70% (for Gram negative bacteria or fungi) were introduced into the wells and optical densities (OD) of each well was measured at 550 nm (Thermo Scientific Multiskan FC, Vantaa, Finland). Experiments were done in triplicate and percentage of inhibition of biofilm by EOs was calculated using the formula:

Violacein Inhibition Assay Against C. violaceum CV12472
The inhibitory effect of violacein produced by C. violaceum CV12472 was assessed through quantitative analysis following a previously described protocol.
41
In this experiment, 10 μL of overnight cultures of C. violaceum CV12472 (with an optical density of 0.4 at 600 nm) were combined with 170 μL of LB broth in sterilized microplates, along with 20 μL of minimum inhibitory concentration (MIC) and sub-MIC concentrations of EOs. A control assay without EOs (LB broth and C. violaceum CV12472) was included as the control. Following incubation at 37 °C for 24 h, absorbance readings at 585 nm were taken using a 96-well plate microplate reader (SpectraMax, Molecular Devices) to measure any reduction in violacein pigment compared to the control. Experiments were repeated three times and the percentage inhibition of violacein was calculated using the following formula:

Quorum-Sensing Inhibition (QSI) Activity Against C. violaceum CV026
The anti-quorum sensing properties of the three EOs were assessed using a modified version of a previously documented technique.53,54 Initially, a mixture of 5 mL Soft Top Agar, comprising 1.3 g agar, 2.0 g tryptone, and 1.0 g sodium chloride dissolved in 200 mL deionized water, was prepared. To this mixture, 100 μL of an overnight culture of C. violaceum CV026 and 20 μL of 100 μg/mL acylhomoserine lactone (AHL) as an exogenous source of signaling molecules were added. The resulting blend was gently poured onto the surface of a solidified LB agar plate to create an overlay. Once the overlay solidified, wells with a diameter of 5 mm were created on each plate and filled with 50 μL of EO at concentrations corresponding to the MIC and sub-MIC. Each experiment was conducted in triplicate, and the plates were then incubated at 30 °C for three days. The diameter of the inhibition zone, characterized by a white or cream-colored halo surrounding the well against a purple lawn of activated C. violaceum CV026 bacteria, was measured in millimeters.
Swarming and Swimming Motility Assay
To assess the effect of EOs on both swimming and swarming motility of P. aeruginosa PA01 strain, culture were inoculated at the center of their respective media and incubated overnight. For swarming motility, the medium consisted of 1% w/v peptone supplemented with 0.5% w/v each of D-glucose, agar, and NaCl, along with sub-MICs of EO, followed by 24-h incubation at 37 °C. 55 The swimming assay followed similar procedures, except for the composition of the medium, which included 1.0% tryptone, 0.5% NaCl, and 0.3% agar. 56 Swarming and swimming extent were quantified by measuring the swarming area (µm²) using ImageJ software and compared to controls. Plates without EO served as untreated controls.
Statistical Analysis
Experiments were done in triplicate and results were expressed as mean value ± standard deviation. The statistical differences between the test samples were analyzed through one-way ANOVA. The differences with P < .05 were considered statistically significant.
Results
The hydro distillation of the aerial parts of studied plants resulted in yellowish oils. The yields of EOs were 0.5% for L. stoechas, 1.62% for T. capitatus, and 1.64% for T. fontanesii. GC-FID and GC-MS were used to determine the chemical composition of the EO from T. capitatus, T. fontanesii, and L. stoechas, and the results are reported in Table 1. The total number of identified compounds was 36 (100%), 27 (100%), and 42 (100%) for T. capitatus, T. fontanesii, and L. stoechas, respectively. The EOs were rich in monoterpene hydrocarbons, comprising 14.99% of the detected components in T. capitatus, 26.54% in T. fontanesii, and 2.53% in L. stoechas. High amounts of oxygenated monoterpenoids were also identified in T. capitatus (83.11%), T. fontanesii (72.77%), and L. stoechas (96.53%). Sesquiterpene hydrocarbons formed 1.44% and 0.58% of the EOs of T. capitatus and T. fontanesii, respectively, and oxygenated sesquiterpenoids 0.22% and 0.11%, respectively. The most abundant compounds in T. capitatus were carvacrol (81.07%), p-cymene (9.15%), and γ-terpinene (4.11%), while in T. fontanesii they were carvacrol (49.08%), γ-terpinene (13.80%), and p-cymene (11.76%). In the EO of L. stoechas, the most abundant compounds were fenchone (44.63%), 1,8-cineole (33.20%), and camphor (14.82%).
Chemical Composition of the Essential Oils from Thymus capitatus, Thymus fontanesii and Lavandula stoechas.

: trace. bold type : composition for major component.
Retention index experimentally determined using homologous series of C7-C30 alkanes on Rxi-5Sil MS fused silica column.
Linear retention index taken from Adams (2007) and/or NIST 08 (2008).
Percentage concentration, Identification methods: Co−GC: Co-injection with authentic compounds, RI: based on comparison of calculated with those reported in ADAMS and NIST 08, MS: based on comparison with WILEY, ADAMS and NIST 08 MS databases.
Enzymes Inhibition Activity of Essential Oils
The study of the anticholinesterase, antidiabetic, anti-urease, and anti-tyrosinase activities of EOs from T. capitatus, T. fontanesii, and L. stoechas reveals significant variations in their respective inhibitory properties as shown on Figure 1. For acetylcholinesterase (AChE) inhibition, T. capitatus EO shows 44.20 ± 0.82% inhibition, and 60.01 ± 0.43% for butyrylcholinesterase (BChE), while T. fontanesii EO displays 38.41 ± 0.77% and 51.20 ± 0.65%, respectively. L. stoechas EO exhibits the highest anticholinesterase activity with 56.63 ± 0.79% inhibition for AChE and 71.45 ± 0.37% for BChE, compared to galantamine, a standard, which showed 92.22 ± 0.18% (AChE) and 80.37 ± 0.27% (BChE) inhibitions. Regarding antidiabetic activity, T. capitatus EO inhibits α-glucosidase at 64.89 ± 0.82% and α-amylase at 62.44 ± 0.78%, surpassing T. fontanesii EO with inhibitions of 58.33 ± 0.39% and 47.68 ± 0.57%, and L. stoechas EO with 51.05 ± 0.90% and 46.70 ± 0.73%. In comparison, acarbose, a standard, inhibits α-glucosidase and α-amylase at 67.30 ± 0.45% and 80.56 ± 0.52%, respectively. For urease inhibition, T. capitatus EO shows 42.71 ± 0.28%, T. fontanesii EO 35.80 ± 0.73%, and L. stoechas EO 32.12 ± 0.46%, compared to thiourea with 89.30 ± 0.24%. Finally, tyrosinase inhibition is 20.57 ± 0.65% for T. capitatus EO, 17.86 ± 0.47% for T. fontanesii EO, and 33.80 ± 0.50% for L. stoechas EO, while kojic acid, used as a standard, shows an inhibition of 82.84 ± 0.67%.

Anticholinesterase, antidiabetic, anti-urease and anti-tyrosinaseactivities of essential oil from Thymus capitatus, Thymus fontanesii and Lavandula stoechas (percentage inhibition at 200 µg/mL).
Antimicrobial Activity
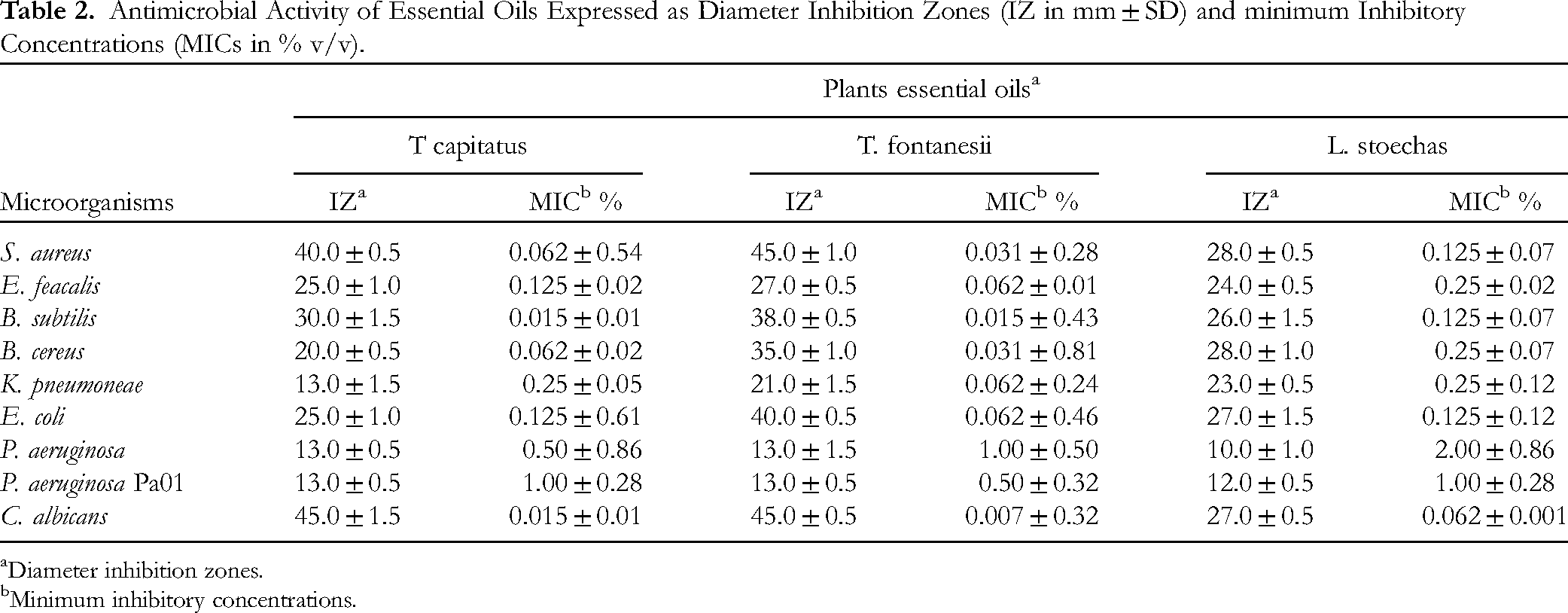
The EOs of T. capitatus, T. fontanesii, and L. stoechas were studied for their antimicrobial activity against several Gram-positive, Gram-negative microorganisms and C. albicans. As shown in Table 2, the diameters of the inhibition zones (IZ in mm ± SD) and the minimum inhibitory concentrations (MICs in % v/v) revealed significant variations depending on the bacterial species and strains tested. For Gram-positive bacteria, T. fontanesii EO demonstrated the strongest activity against S. aureus, with an IZ of 45.0 ± 1.0 mm and a MIC of 0.031 ± 0.28%, followed by T. capitatus EO with an IZ of 40.0 ± 0.5 mm and a MIC of 0.062 ± 0.54%. In contrast, L. stoechas exhibited an IZ of 28.0 ± 0.5 mm and a MIC of 0.125 ± 0.07%. Similarly, E. faecalis was more sensitive to T. fontanesii EO (IZ of 27.0 ± 0.5 mm, MIC of 0.062 ± 0.01%) compared to T. capitatus EO (IZ of 25.0 ± 1.0 mm, MIC of 0.125 ± 0.02%) and L. stoechas EO (IZ of 24.0 ± 0.5 mm, MIC of 0.25 ± 0.02%). For B. subtilis, T. fontanesii EO exhibited an IZ of 38.0 ± 0.5 mm and a MIC of 0.015 ± 0.01%, while T. capitatus EO showed an IZ of 30.0 ± 1.5 mm with the same MIC, and L. stoechas EO had an IZ of 26.0 ± 1.5 mm with an MIC of 0.125 ± 0.07%. Regarding B. cereus, the inhibition zones were 35.0 ± 1.0 mm for T. fontanesii EO (MIC of 0.031 ± 0.81%), 20.0 ± 0.5 mm for T. capitatus EO (MIC of 0.062 ± 0.02%), and 28.0 ± 1.0 mm for L. stoechas EO (MIC of 0.25 ± 0.07%).
Antimicrobial Activity of Essential Oils Expressed as Diameter Inhibition Zones (IZ in mm ± SD) and minimum Inhibitory Concentrations (MICs in % v/v).
Diameter inhibition zones.
Minimum inhibitory concentrations.
Although Gram-negative bacteria are generally less sensitive, the EOs of T. capitatus and T. fontanesii demonstrated significantly greater antimicrobial activity than EO of L. stoechas. Against some Gram-negative bacteria, such as E. coli and K. pneumoniae, the Eos showed significant inhibition zones especially T. fontanesi. K. pneumoniae was more sensitive to T. fontanesii EO, with an IZ of 21.0 ± 1.5 mm and an MIC of 0.062 ± 0.24%, followed by L. stoechas EO (IZ of 23.0 ± 0.5 mm, MIC of 0.25 ± 0.12%) and T. capitatus EO (IZ of 13.0 ± 1.5 mm, MIC of 0.25 ± 0.05%). Against E. coli, an IZ of 40.0 ± 0.5 mm was exhibited by T. fontanesii (MIC of 0.062 ± 0.46%), followed by T. capitatus EO (IZ of 25.0 ± 1.0 mm, MIC of 0.125 ± 0.61%) and L. stoechas EO (IZ of 27.0 ± 1.5 mm, MIC of 0.125 ± 0.12%). IZ were modest against P. aeruginosa as follows: T. capitatus EO (IZ of 13.0 ± 0.5 mm, MIC of 0.50 ± 0.86%), T. fontanesii EO (IZ of 13.0 ± 1.5 mm, MIC of 1.0 ± 0.5%), and L. stoechas EO (IZ of 10.0 ± 1.0 mm, MIC of 2.00 ± 0.86%). The EOs of T. capitatus and T. fontanesii also showed strong activity against C. albicans, with IZs of 45.0 ± 0.5 mm and 45.0 ± 1.5 mm respectively, and MICs of 0.007 ± 0.32% and 0.015 ± 0.01%. L. stoechas exhibited an IZ of 27.0 ± 0.5 mm and an MIC of 0.062%.
Antibiofilm Activity
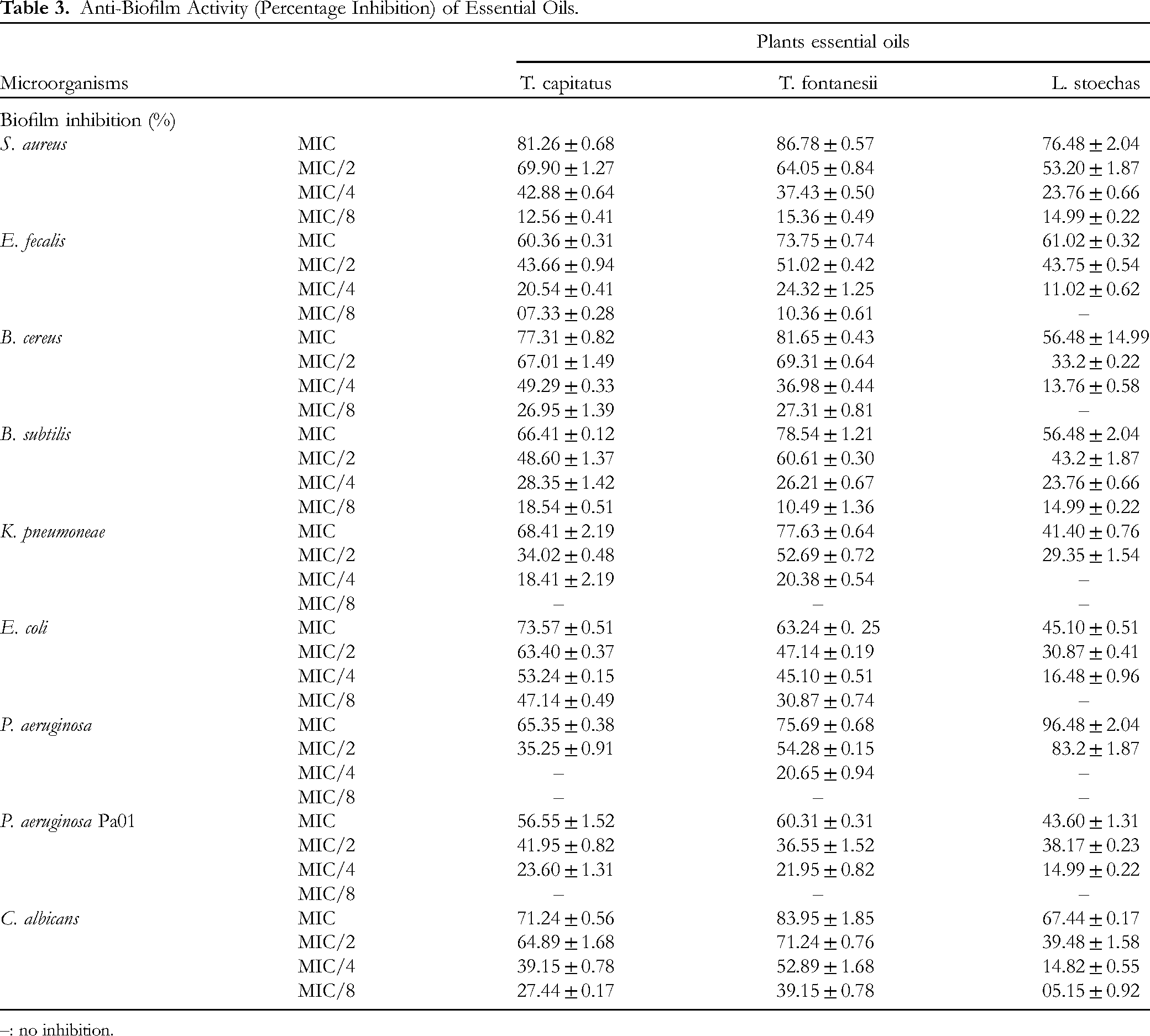
The EOs of T. capitatus, T. fontanesii, and L. stoechas demonstrated varying capacities to inhibit biofilms against different microbial strains, in a concentration-dependent manner as shown in Table 3. At the MIC, T. capitatus EO significantly inhibited biofilms, reaching up to 81.26% inhibition against S. aureus and exceeding 56% for other strains. T. fontanesii EO surpassed T. capitatus EO against many strains, with inhibitions ranging from 63.24% for E. coli to 86.78% for S. aureus. L. stoechas EO showed moderate efficacy, though with notable inhibition against P. aeruginosa (96.48%). At MIC/2, a reduction in the efficacy of the EOs was observed, but they EOs remained active. T. capitatus EO showed significant inhibitions, notably 69.90% against S. aureus, while T. fontanesii EO exhibits good activity notably with inhibition of 64.05% against S. aureus. L. stoechas EO had lower activity but with a notable inhibition against P. aeruginosa (83.20%). At MIC/4 and MIC/8, inhibitions further decreased. T. capitatus EO and T. fontanesii EO exhibited appreciable activities though with percentage inhibitions below 50%. In conclusion, T. fontanesii EO proved to be the most effective at inhibiting biofilms, even at reduced concentrations, with greater activitiy than EOs from T. capitatus and L. stoechas.
Anti-Biofilm Activity (Percentage Inhibition) of Essential Oils.
–: no inhibition.
Inhibition of Violacein Production
The MICs of the EOs were pre-determined so that violacein, QS and motility inhibitions can be measured at MIC and sub-MIC concentrations which are lower enough to avoid excessive pressure of bacteria and also eliminate possibility of cell death. For the inhibition of violacein production in C. violaceum CV12472, T. capitatus EO and T. fontanesii EO showed complete inhibition (100%) at the MIC (0.031%), with a gradual but substantial reduction down to 20.47% and 32% respectively at MIC/32. In contrast, L. stoechas EO, with an MIC of 0.25%, showed almost complete inhibition at MIC (96.48%), but this efficacy decreased more rapidly, reaching 7.95% at MIC/16 and no inhibition at MIC/32. Detailed results of violacein inhibition are presented in Figure 2.
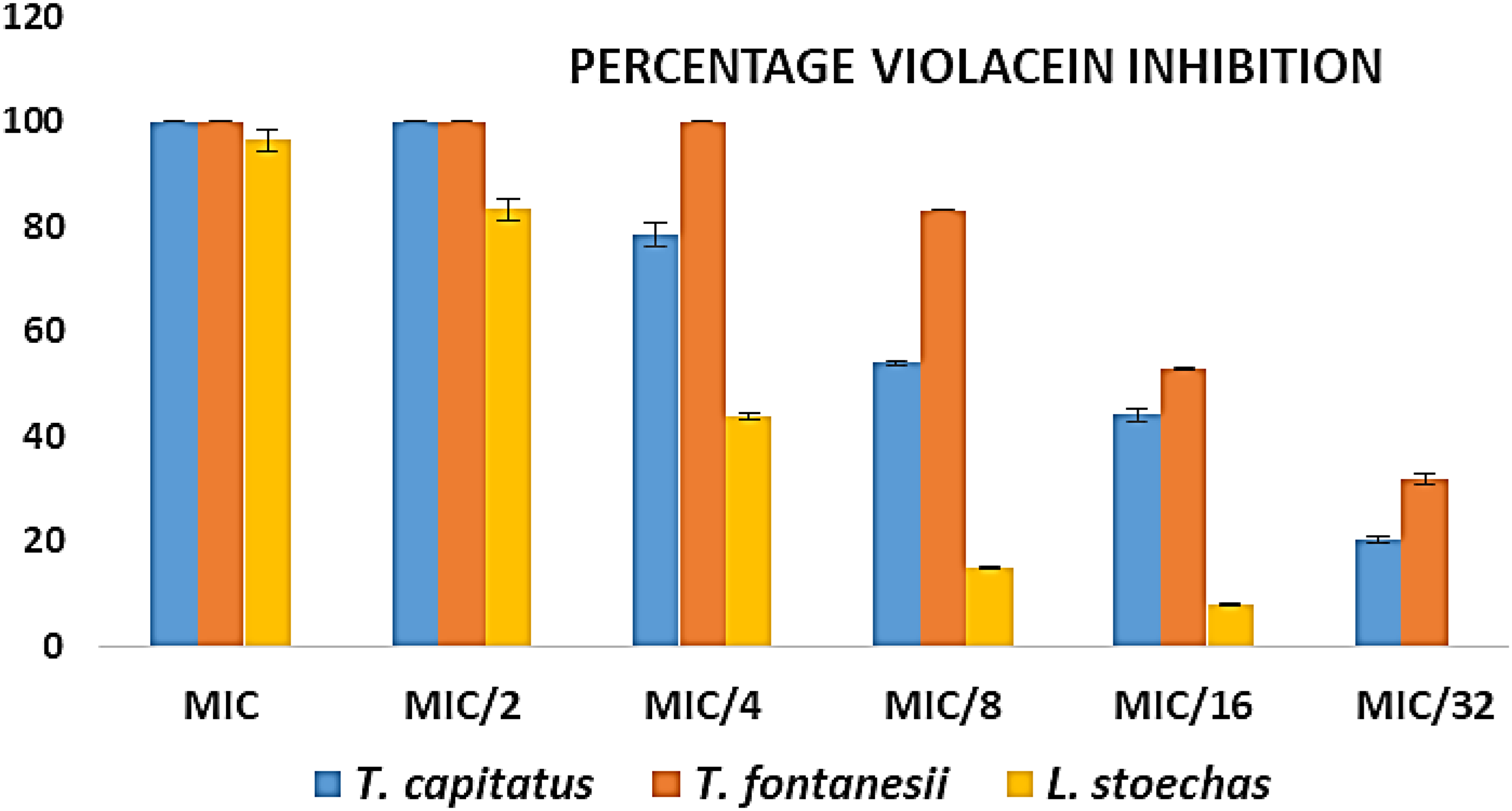
Inhibition of violacein production against C. violaceum CV12472 by essential oils from T. capitatus, T. fontanesii and L. stoechas.
Quorum-Sensing Inhibition
The evaluation of QS inhibition zones with C. violaceum CV026 strains shows that T. capitatus EO and T. fontanesii EO have similar effects as shown in Figure 3, with inhibition zones of 25.21 mm and 28.00 mm at the MIC respectively, which reduces to 8.41 mm (T. capitatus EO) and 8.02 mm (T. fontanesii EO) at MIC/4, with no notable inhibition at lower dilutions. L. stoechas EO, although having a higher MIC (0.5%), produced inhibition zones of 30.04 mm at the MIC, which decreased to 7.99 mm at MIC/8.

Quorum-sensing inhibition zones against C. violaceum CV026 by by essential oils from T. capitatus, T. fontanesii and L. stoechas.
Swimming and Swarming Motility Inhibition
Regarding the inhibition of swimming and swarming motilities of P. aeruginosa PA01, the results (Figure 4) show strong inhibition at all tested concentrations. T. capitatus EO showed 95.05% and 93.32% inhibition of swimming and swarming at the MIC, decreasing to 34.84% and 29.84% at MIC/8, respectively. T. fontanesii EO was slightly more active with 98.54% (swimming) and 96.59% (swarming) inhibition at the MIC, reduced to 25.61% (swimming) and 34.75% (swarming) at MIC/8. L. stoechas EO demonstrated inhibition rates of 85.53% (swimming) and 83.64% (swarming) at the MIC, but these rates dropped significantly to 7.05% (swimming) and 19.95% (swarming) at MIC/8.
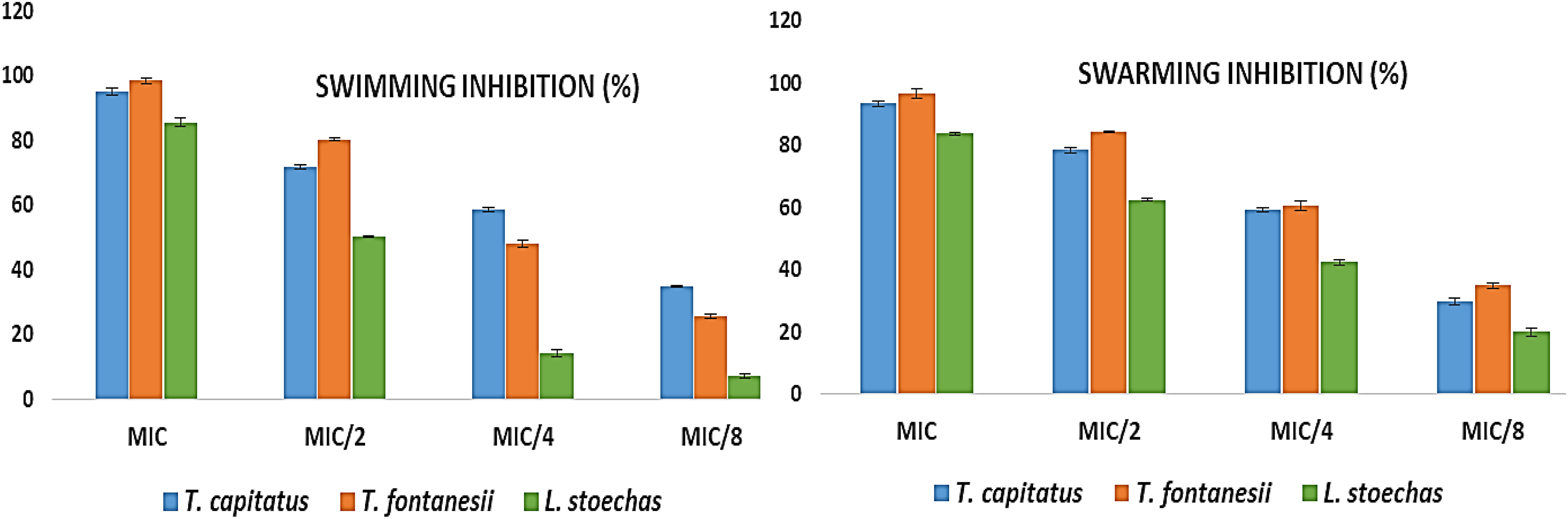
Swimming and swarming motility inhibition against P. aeruginosa PA01 by by essential oils from T. capitatus, T. fontanesii and L. stoechas.
Discussion
Our results from the chemical analysis of the EO of T. fontanesii revealed both similarities and significant differences compared to previous studies. Bekhechi et al 57 examined five EO samples of T. fontanesii, finding higher levels of carvacrol (66.7 to 69.5%) and lower amounts of p-cymene and γ-terpinene, with minor variations for other constituents such as linalool and α-pinene. Nabet et al (2017) obtained results similar to ours regarding the proportions of hydrocarbon monoterpenes (27.2%) and oxygenated monoterpenes (71.8%), but with a higher concentration of carvacrol (67.5%). 58 Benyoucef et al (2018) reported a composition dominated by monoterpenoid phenols (84.7%), with thymol (76.6%), as the main component, which represents a great difference with this study where thymol is almost absent. 59 Finally, Sidali et al (2020) identified carvacrol as the main compound (54.7% to 63.9%), with similar proportions of p-cymene and γ-terpinene, although the amount of carvacrol was slightly lower than our results. 60 For T. capitatus EO, GC-MS analysis revealed that monoterpene hydrocarbons represented 14.99% of the detected compounds. Oxygenated monoterpenoids were predominant, constituting 83.11% of the total composition. Sesquiterpene hydrocarbons accounted for 1.44%, and oxygenated sesquiterpenoids 0.22%. The most abundant compounds in T. capitatus EO were carvacrol (81.07%), p-cymene (9.15%), and γ-terpinene (4.11%). T. capitatus is endemic to the Mediterranean region, and several studies have characterized its chemical composition. For example, a study by Bounatirou et al (2007) in Tunisia showed that the EO of T. capitatus, collected at different development stages in three distinct locations, was mainly composed of carvacrol (61.6% to 83%), p-cymene (5% to 17%), γ-terpinene (2% to 14%), and β-caryophyllene (1% to 4%). 61 This composition varies depending on the location and growing season. Similarly, carvacrol was the major compound in 29 samples of T. capitatus from Sicily, varying between 49% and 83%, according to Napoli et al (2010). 62 The EO of T. capitatus in Albania 63 and Portugal 64 also showed high levels of carvacrol. Another study by Casiglia et al (2019) demonstrated that the EO of T. capitatus, growing wild in Salinelle, Sicily, and collected at three different vegetative stages, was characterized by high levels of carvacrol (14.2% to 81.2%), γ-terpinene (2.6% to 34.4%), and p-cymene (5.0% to 22.8%) of the total oil content. 65 The yield and composition of the EO vary throughout the plant's growing period. More recently, Tagnaout et al (2022) revealed that the EO of T. capitatus was rich in carvacrol (75%), p-cymene (10.58%), linalool (2.91%), (E)-caryophyllene (1.61%), and epoxy caryophyllene (1.5%). 27 However, it is important to note that some authors have identified a thymol chemotype in the EO of T. capitatus, as shown in the study by Goudjil et al (2020) in Algeria, where thymol represented 51.22% of the total EO content, followed by carvacrol (12.59%) and γ-terpinene (10.3%). 66 Similarly, the EO of T. capitatus from Sardinia, Italy, was dominated by thymol (29.3%) and p-cymene (26.4%), while carvacrol accounted for only 10.8%. 67 The EO of T. capitatus from Tizi Ouzou, Algeria, had a composition of thymol (25.82%), linalool (23.40%), and geraniol (14.22%). 68 Our chemical analysis of the EO of L. stoechas revealed that the most abundant compounds were fenchone (44.63%), 1,8-cineole (33.20%), and camphor (14.82%). These results are consistent with numerous previous studies that have also identified fenchone, camphor, and 1,8-cineole as the main compounds of this species. For example, Angioni et al (2006) showed that the fenchone/camphor chemotype predominates in the EOs of L. stoechas in Sardinia, with fenchone contents ranging from 52.60% to 66.20% and camphor from 13.13% to 27.08%. 69 Additionally, Benabdelkader et al (2014) observed similar variations in Algeria, with fenchone concentrations from 11.27% to 37.48% and camphor from 1.94% to 21.8%. 35 La Bella et al (2014) also found high levels of fenchone (52.8% to 71.1%) in Sicilian populations. 70 However, some studies have highlighted regional differences in the composition of L. stoechas EOs. Gursoy et al (2009) reported the presence of cymene in Turkish samples, a compound rarely found elsewhere. 71 Baali et al (2019) found a predominance of fenchone (50.29%) and camphor (14.02%) in Algerian samples, but also a notable presence of bornyl acetate (5.60%), a compound absent from our analysis. 72 Differences in chemical composition can be explained by various geographical origins, climatic and seasonal conditions, collection periods, stages of maturity, and extraction methods.
The enzyme α-amylase hydrolyzes starch into simple sugars, while α-glucosidase catalyzes the breakdown of glucose from disaccharides, and both processes contribute to the development of diabetes. Inhibiting these enzymes can slow carbohydrate digestion and reduce glucose absorption. Therefore, α-amylase and α-glucosidase inhibitors are used to lower postprandial plasma glucose levels, particularly in the management of type 2 diabetes and obesity. In our enzyme inhibition assays, T. capitatus exhibited α-glucosidase inhibition at 64.89 ± 0.82% and α-amylase inhibition at 62.44 ± 0.78%. These results surpass those of T. fontanesii, (P < .05) which showed inhibitions of 58.33 ± 0.39% for α-glucosidase and 47.68 ± 0.57% for α-amylase, as well as L. stoechas, with 51.05 ± 0.90% and 46.70 ± 0.73%, respectively. In comparison, acarbose, a reference inhibitor, achieved inhibition rates of 67.30 ± 0.45% for α-glucosidase and 80.56 ± 0.52% for α-amylase. Previous studies have reported that the plant T. capitatus has an IC50 of 0.8 ± 0.1 mg/mL for α-amylase inhibition, 73 while Iauk et al 74 noted IC50 values of 422.5 mg/mL (n-hexane fraction) and 565.9 mg/mL (methanol fraction) for the same enzyme. More recently, Barkat 75 demonstrated that EOs from T. capitatus have an α-amylase inhibitory effect approximately 50% of that of acarbose at a concentration of 400 μg/mL, which aligns with our observations. Currently, there are no published studies on T. fontanesii regarding the inhibition of α-glucosidase and α-amylase, highlighting the significance of our findings for this species. For L. stoechas, Mustafa et al 76 showed that the hydroalcoholic extract reduces hyperglycemia in diabetic mice, suggesting a broader antidiabetic potential. Additionally, Sebai et al 77 proposed that D-fenchone, a major component of lavender oil, might contribute to protective effects against diabetes, although our results do not confirm the specific contribution of this compound. El Omari et al 78 reported an IC50 of 76.92 ± 2.43 μg/mL for camphor against α -amylase, while fenchone and L. stoechas EO presented IC50 values of 104.19 ± 78 μg/mL and 106.73 ± 3.27 μg/mL, respectively. Concerning α-glucosidase, fenchone exhibited an IC50 of 69.03 ± 2.31 μg/mL, surpassing acarbose (IC50 = 199.53 ± 3.26 μg/mL). Finally, El Hachlafi et al 79 reported IC50 values of 3.00 ± 0.04 mg/mL for α-amylase and 2.74 ± 0.01 mg/mL for α-glucosidase.
Inhibiting the enzymes AChE and BChE is crucial because they degrade acetylcholine, a key neurotransmitter for nerve transmission. This approach is promising for treating Alzheimer's disease, where maintaining high levels of acetylcholine is essential. Plants, rich in bioactive compounds, offer a potential source of natural inhibitors of these enzymes, playing an important role in the development of new treatments. 80 In our study, T. capitatus EO and L. stoechas EO demonstrated significant inhibition of both AChE and BChE (P < .05), while T. fontanesii EO showed more modest inhibition. Notably, no prior studies have investigated the inhibitory properties of T. fontanesii for these enzymes, highlighting the novelty of our findings for this species. Previous research provides useful context. Aazza et al 73 and Albano et al 81 reported that the essential oil of T. capitatus, primarily composed of carvacrol, exhibits high inhibition of AChE, with an IC50 of 0.03 mg/mL. Jaouadi et al 82 also observed significant AChE inhibition for Tunisian T. capitatus oil, with IC50 values ranging from 0.11 to 0.61 mg/mL. Additionally, Mousafq et al 83 found that the methanolic extract of L. stoechas significantly reduces brain AChE activity, confirming our findings of high inhibition for this plant. However, these inhibitions are still lower than those achieved with galantamine, a reference inhibitor.
Urease, a nickel-containing enzyme, catalyzes the hydrolysis of urea into carbon dioxide and ammonia. This enzyme is associated with pathological conditions such as gastric and gastroduodenal ulcers, making its inhibition crucial for countering H. pylori by preventing its adherence to the gastric mucosa.41,84 Tyrosinase, a copper-containing enzyme, plays a key role in melanin pigment biosynthesis. Inhibiting tyrosinase can address hyperpigmentation disorders and skin melanoma, positioning these inhibitors as promising agents for skin whitening, aging, and other dermatological conditions. 85 Our results show that EOs of T. capitatus, T. fontanesii, and L. stoechas exhibit notable inhibition of urease and tyrosinase, although this inhibition is lower than that observed with standards such as thiourea for urease and kojic acid for tyrosinase. T. capitatus EO and T. fontanesii EO provide moderate inhibition of urease and tyrosinase, while L. stoechas demonstrates a higher inhibition of tyrosinase (33.80%) but remains also lower than kojic acid (82.84%). The results from El Omari et al 78 highlight that fenchone, a major component of L. stoechas EO, exhibits remarkable activity with an IC50 value of 53.14 ± 3.06 μg/mL, surpassing the EO and camphor, which have IC50 values of 66.21 ± 3.10 μg/mL and 97.45 ± 5.22 μg/mL, respectively. This suggests a higher potential for natural compounds in developing new treatments for pigmentation disorders and related conditions.
The antimicrobial activities of EOs from plants are often attributed to the presence of certain specific constituents. Numerous studies have demonstrated that compounds such as carvacrol,86,87 thymol, 88 p-cymene, 89 γ-terpinene,90,91 fenchone, 92 camphor, 93 and 1,8-cineole91,94 possess powerful antimicrobial properties. Our oils contain all of these compounds, some of which are even major components. Our EO of T. capitatus, with a chemotype rich in carvacrol (81.07%), demonstrates antibacterial activity that is interestingly comparable to the results of several previous studies. Jayari et al 95 reported that their Tunisian EO, also rich in carvacrol (88.98%), exhibited strong activity against E. coli, S. typhimurium, and S. aureus, but had lower sensitivity against P. aeruginosa, which aligns with our observation. Amarti et al 96 found that the Moroccan EO, containing 70.92% carvacrol, inhibited E. coli and S. aureus at a concentration of 1/2000 (v/v), which is consistent with our efficacy against these bacteria. However, notable differences emerge when comparing with EOs rich in thymol. Aouadhi et al 97 observed that the T. capitatus EO from Bizerte, containing 81.49% thymol, had significantly higher antibacterial activity than that from Sousse, with MICs ranging between 0.025% and 0.8%, suggesting superior efficacy compared to our carvacrol-rich EO. Similarly, Hajlaouia et al 98 found that the EO containing 39.66% thymol had higher antimicrobial activity, with MICs for bacteria ranging from 0.019 to 0.15 mg/mL, and for fungi, MICs ranged from 0.004 to 0.078 mg/mL. Studies by Goudjil et al 66 and Barakat et al 75 on T. capitatus EOs from Algeria and Palestine, respectively, show that EOs containing thymol and γ-terpinene present broader antibacterial activity. Goudjil et al reported MICs of 29.44, 14.72, 14.72, and 7.36 μg/mL against S. aureus, S. typhi, E. coli, and S. pneumoniae, while Barakat et al observed MICs ranging from 0.1953 to 1.5625 μg/mL for various chemotypes, including carvacrol and γ-terpinene. These results highlight that EOs rich in thymol or γ-terpinene may exhibit stronger activity against a wider range of bacteria. In conclusion, existing data indicate that the antibacterial activity of T. capitatus EO is largely influenced by its chemical composition. While our EO, rich in carvacrol, is effective against certain bacteria, its activity is less pronounced against Gram-negative bacteria like P. aeruginosa, compared to EOs rich in thymol or γ-terpinene. This variation in efficacy is consistent with the literature, emphasizing the need to select the appropriate chemotype to optimize antibacterial properties. Finally, the lower activity observed for our EO against P.s aeruginosa, partly due to the predominance of carvacrol, is consistent with the findings of Ibraliu et al (2011), who noted the specific resistance of this strain to carvacrol due to the MexAB-OprM efflux pump. The results obtained with the EO of T. fontanesii show significant antimicrobial activity against a range of microorganisms, with IZ ranging from 13 to 45 mm and MIC varying from 0.007% to 1% v/v. These findings are consistent with data reported in previous studies on T. fontanesii EOs. Bekhechi et al 57 observed significant antimicrobial activity of T. fontanesii oil against various bacteria, except P. aeruginosa, with notable activity against S. typhii, E. coli, and other Gram-negative bacteria. This study highlights comparable antimicrobial activity against S. aureus and E. coli, with MICs of 300 µg/mL for S. aureus and 380 µg/mL for E. coli, whereas our study reports lower MICs of 0.031% v/v for S. aureus and 0.062% v/v for E. coli. The work of Dob et al 99 shows that T. fontanesii oil has a low MIC for Fusarium oxysporum and B. subtilis, with similar MICs for S. cerevisiae and C. albicans. These observations align with our results, where the MIC for B. subtilis is 0.015% v/v and for C. albicans is 0.007% v/v, highlighting the oil's effectiveness against these strains. Nabet et al 58 reported that T. fontanesii essential oil exhibited superior antimicrobial activity, with inhibition zone diameters up to 48.56 mm for S. aureus and 44.99 mm for C. albicans. The MICs for these strains were 0.11 µL/mL, similar to our findings where the MIC for S. aureus is 0.031% v/v and for C. albicans is 0.007% v/v. Sidaali et al 100 found that inhibition zones for bacteria ranged from 11 to 36 mm, with maximum activity observed against Candida species and variable sensitivity for Gram-positive and Gram-negative bacteria. Our study also shows strong activity against C. albicans (IZ of 45 mm and MIC of 0.007% v/v), but less effectiveness against P. aeruginosa (IZ of 13 mm and MIC of 1% v/v), reflecting challenges noted in previous studies. Finally, Abdellah et al 101 reported MICs for various bacterial strains, ranging from 3.44 to 5.26 µL/mL for the tested strains. These results are consistent with our findings, where MICs for S. aureus and E. coli are below 0.062% v/v, while MICs for P. aeruginosa are higher, at 1% v/v. The results obtained with the EO of L. stoechas show notable antimicrobial activity against a range of microorganisms, with IZ ranging from 10 to 28 mm and MIC varying from 0.062% to 2% v/v. These results largely align with those reported in previous studies on L. stoechas EO from different geographical regions. For example, Goren et al 102 and Bouzouita et al (2005) 103 observed strong antimicrobial activity of L. stoechas EO from Turkey and Tunisia, with similar MIC values for S. aureus. Similarly, the work of Kırmızıbekmez et al 104 demonstrated significant activity against S. aureus (MRSA), with MICs up to 125 µg/mL for leaf and flower oils, comparable to the current results where the MIC for S. aureus is 0.125% v/v. Studies by Sarac and Ugur 105 on EOs of L. stoechas cultivated in Turkey showed activity against several Gram-positive and Gram-negative bacteria, aligning with our observations of inhibition for strains like E. coli and K. pneumoniae. The work of Benabdelkader et al 35 in Algeria confirms this efficacy, showing similar antimicrobial activities against S. aureus. Moroccan studies, notably those by Cherrat et al 106 and Bouyahya et al, 107 indicate high antimicrobial activity against E. coli and S. aureus, with inhibition diameters and MICs close to our results (IZ of 27 mm for E. coli and MIC of 0.125% v/v). Finally, Baali et al 72 also reported comparable activity against P. aeruginosa, although this strain shows higher resistance, consistent with our observation of a higher MIC for P. aeruginosa (2% v/v).
To the best of our knowledge, there are very few studies, if any, on the antibiofilm activity of EOs from T. capitatus, T. fontanesii, and L. stoechas. Furthermore, no research has been conducted regarding the anti-quorum sensing activity of the EOs of T. fontanesii and T. capitatus, as well as their effect on swimming and swarming. The results of our study provide an interesting perspective on the antibiofilm activity of the EOs from T. capitatus, T. fontanesii, and L. stoechas. T. capitatus showed significant inhibition of biofilms, reaching up to 81.26% for S. aureus at MIC, with reduced but still notable effectiveness at lower concentrations. These results partially align with the observations of Alabdullatif et al, 108 who reported a logarithmic reduction of 1.17 to 1.42 log10 CFU/mL for S. epidermidis at concentrations of 10% to 30% v/v. However, the reductions observed in our study were lower than those reported by Almeida et al, with an average decrease of 2.6 log10 CFU/mL for methicillin-resistant S. aureus, suggesting potential differences in experimental methods or concentrations used. Another study by Vrenna et al 56 found that the essential oil of Coridothymus capitatus (synonymous with T. capitatus) at 1% v/v inhibited biofilm formation and reduced preformed biofilms in approximately half of the eleven clinical isolates of P. aeruginosa from cystic fibrosis patients. Essid et al 109 demonstrated that the EOs of T. capitatus inhibited the growth of C. albicans with an MIC of 125 µg/mL. Additionally, at MIC/2 concentration, this EO significantly reduced the biofilm formation of C. albicans, achieving 80.6% inhibition. The work of Machado et al 110 showed a reduction in biofilm biomass of Gardnerella vaginalis ranging from 49% to 91% with T. capitatus. Moreover, Maniki et al 111 found that the EO of T. capitatus had a MBIC of 0.03% v/v for L. monocytogenes and Y. enterocolitica. This result reinforces the idea that T. capitatus possesses significant antibiofilm activity against various bacterial strains. For T. fontanesii, our results show superior antibiofilm activity compared to T. capitatus, with inhibitions reaching up to 86.78% for S. aureus at MIC. This result is particularly relevant as T. fontanesii has not been specifically studied in the literature, suggesting promising potential for this essential oil in biofilm control. Regarding L. stoechas, our study reveals notable inhibition of P. aeruginosa biofilms (96.48%) at MIC, although the effectiveness quickly decreases at lower concentrations. These results are consistent with the work of ÜNLÜ et al, 112 who found a minimum biofilm inhibitory concentration (MBIC) of 5 µL/mL against S. aureus ATCC 29213 and 2.5 to 10 µL/mL for methicillin-resistant strains, as well as with those of Hickl et al, 113 who reported inhibition of biofilm formation by Streptococcus mutans at concentrations as low as 0.04 mg/mL. Karpiński et al 114 also found that L. stoechas at 1.47 mg/mL inhibited 85% of the biofilms of C. albicans ATCC 14053.
The MIC of EOs were assessed for their ability to inhibit QS, violacein production and motility in P. aeruginosa. T. capitatus and T. fontanesii demonstrated complete inhibition of violacein production at the MIC, although this effectiveness diminished at lower concentrations. L. stoechas also showed nearly total inhibition at the MIC, with a rapid decrease at lower dilutions. Regarding bacterial QS inhibition, T. capitatus and T. fontanesii produced significant inhibition zones at the MIC, reduced to about 8 mm at lower concentrations, while L. stoechas showed a broader inhibition zone at the MIC but with a rapid decrease at lower concentrations. For motility, all EOs showed significant effects, with T. fontanesii being the most effective. These results align with previous studies by Ceylan et al, 115 who found that the MIC of L. stoechas EO against C. violaceum was 0.39 µL/mL, with no detectable anti-QS activity but a reduction in violacein pigment production. The MICs against P. aeruginosa were above 50 µl/ml, with QS-dependent migration inhibition reaching 38% for swarming and 30% for swimming at 50 µl/ml. Additionally, Vrenna et al 56 showed that T. capitatus EO at 1% v/v significantly inhibited motility and biofilm formation in P. aeruginosa. Although studies on QS inhibition, violacein production, and motility of P. aeruginosa are scarce, it has been shown that these activities are also limited and inhibited by other Lamiaceae such as T. vulgaris, 116 T. musilii, lavender and their major compounds like thymol and carvacrol.117,118 The studied essential oils can be used in aromatherapy to remedy a variety of clinical conditions. By applying the EOs, it can restore healthy condition. By modifying the functional properties of EOs, they directly affect physiological processes in the human body and can be used in cosmetics, for massage, medical preparations, olfactory and psycho-aromatherapy. The emergence of antimicrobial resistance and other ailments has urged researchers to explore therapeutic alternatives, such as essential oils (EO) notably from clove, oregano, thymus, cinnamon bark, rosemary, eucalyptus and lavender that have shown significant inhibitory effects against microorganisms. 119 Essential oils are natural aromatic liquids obtained from fruits, flowers, seeds, stems, bark, and other parts of the plant with numerous biological activities exploitable in conventional medication especially to treat infectious diseases, neurological disorders among orders. 120
Nonetheless, this study may involve some limitations. The poor solubility of essential oils in water and organic solvents may affect the results obtained in studies involving EO. Also, the components in the essential oils may act synergistically or some may have antagonistic effects on others, thereby influencing the results when testing the essential oils in bulk.
Conclusion
Lamiaceae also known as sage family of flowering plants with great aromatic qualities and rich in relevant essential oils. They are widespread, easily cultivated and propagated by stem cuttings. They are used for seasoning of foods, as herbal teas and also for medicinal purposes. In this study, three plants from this family namely, T. capitatus, T. fontanesii and L. stoechas were evaluated for their chemical composition through GC-MS as well as their inhibitory effects on certain enzymes and microbial virulence factors. The EOs inhibited cholinesterase enzymes, indicating that they can be used as remedy to relieve the symptoms of cognitive impairments involved in Alzheimer's disease. Additionally, enzymes (α-amylase and α-glucosidase) that hydrolyze starch into glucose causing rise in blood sugar levels were inhibited by the EOs, thereby indicating their potential in reducing the emergence of type 2 diabetes. Furthermore the EOs showed low inhibitory potentials of urease and tyrosinase, indicating that they can reduce the effects of ureolytic bacteria and also prevents hyperpigmentation of skin. At low concentrations, the EOs were able to inhibit growth of bacteria and fungi, biofilm formation, violacein production, swimming and swarming motilities are important virulence factors in bacteria and fungi which the EOs inhibited in this study. This shows that the EOs can reduce the pathogenicity of microbes and attenuate severity of infections. These EOs can find applications s preservatives and biocontrol substances. The future perspectives of this study will involve clinical aromatherapy, which is the incorporation of Lamiaceae essential oils as a complementary therapy within the medical field in accordance with medical safety, ethical, and regulatory requirements.
Footnotes
Acknowledgements
This work is supported by the General Directorate for Scientific Research and Technological Development (DGRSDT), Algeria. Material and technical support from Mugla Sitki Kocman University, Turkey is greatly acknowledged.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical Approval
Ethical approval is not applicable to the article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Statement of Human and Animal Rights
This article does not contain any studies with human or animal subjects.
Statement of Informed Consent
There are no human subjects in this article and informed consent is not applicable.