
Abstract
Objective
Phyto-oxylipins are lipid molecules produced in plants by the oxidative transformation of unsaturated fatty acids via diverse metabolic pathways. Recently, the chemical diversity and functional significance of oxylipins is gaining significant attention. However, the functional significance of these compounds as anti-cancer agents remains largely uncharacterized. The objective of this review is to provide a comprehensive synthesis and analysis of the functional significance of plant oxylipins as anti-cancer agents to facilitate their exploitation in drug discovery and development.
Methods
This review was based on a thorough compilation and analysis of research work carried out on biological significance and implications of plant-derived anti-cancer oxylipins. Curation of data was based on several databases and resources such as Scopus, PubMed, DrugBank and PubChem. Within the context of the scope and subject matter as guided by the objective, no exclusion and inclusion criteria were necessarily employed in the screening of articles.
Results
The present review explores the origins, anti-cancer properties and functional mechanisms of phyto-oxylipins. The potential functional significance of new and poorly characterized plant oxylipins have also been highlighted. The prospects of plant oxylipins in research, medicine and biotechnology that could optimize their potential are also explored. Insights into the promising avenues that may originate from innovative therapeutic approaches are also discussed.
Conclusion
Despite the rich source of oxylipins in plants, much of their potential as therapeutic agents for cancer treatment remains to be fully established. Clinical investigations are also needed to determine safe doses and effective delivery methods. Research into phyto-oxylipins require significant attention due to the promise it may hold in addressing key challenges in biotechnology, health, and environmental sustainability.
Introduction
The transformation of normal cells into cancer cells is a multifaceted and intricate process central to the development of cancer. 1 Tumorigenesis involves genetic alterations in the genome coupled with evolution into multiple mechanisms over an extended period. 2 It is essential to comprehend the complex molecular mechanisms of tumorigenesis such as those mediated by various forms of stress factors (Figure 1). This is particularly important in order to facilitate the development of effective approaches for cancer prevention, early detection, and treatment. Understanding the intricate processes of tumorigenesis and cancer development is paramount for devising effective strategies against the biological and clinical consequences of uncontrolled cell divisions.
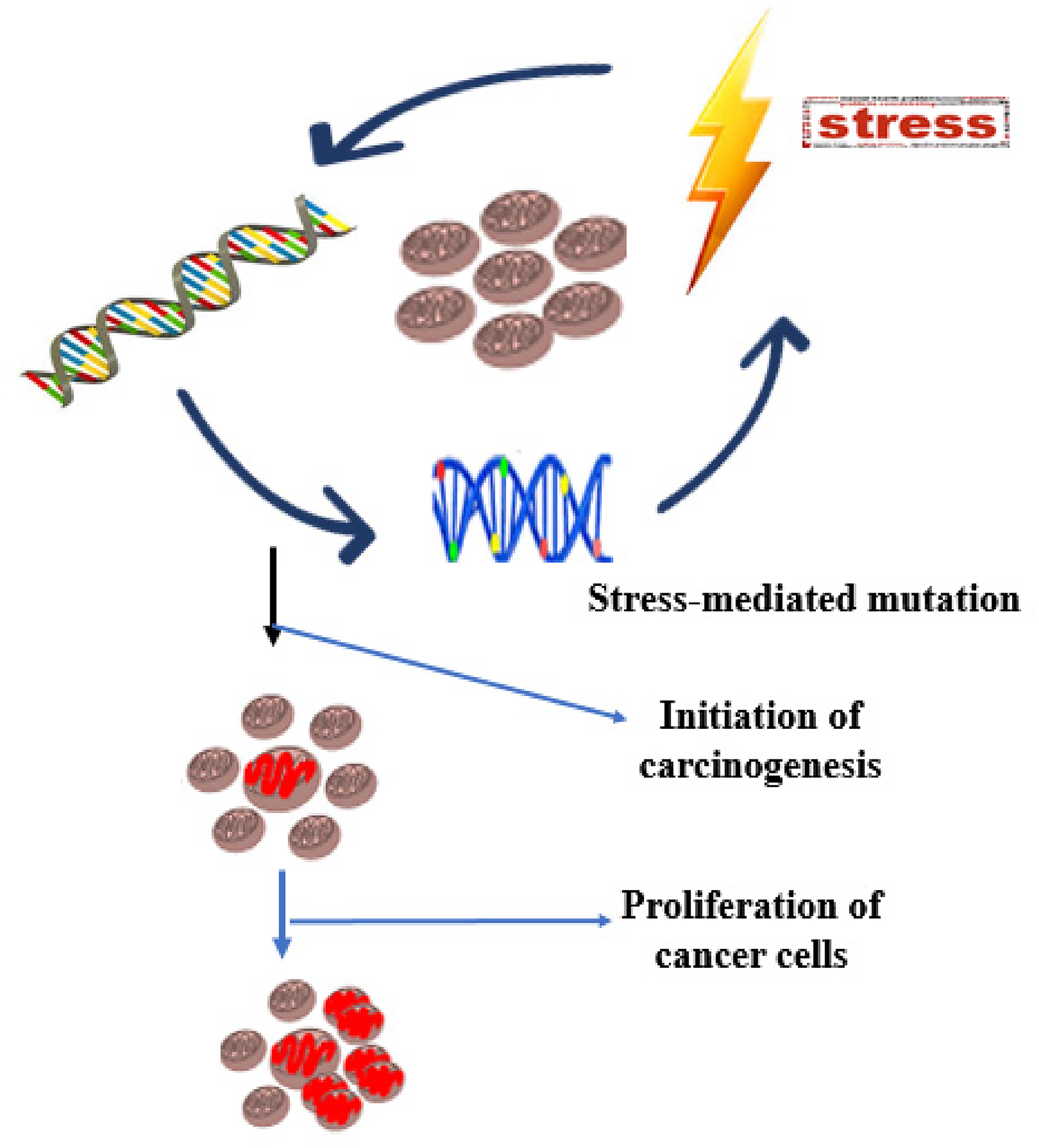
Stress-mediated proliferation of cancer cells. Stress-mediated mutations are often fundamental to the initiation of carcinogenesis. Stress-related factors may include free radicals, UV radiation and other forms of carcinogens/mutagens.
Oxylipins are lipid-soluble signaling molecules formed through enzymatic reactions or the oxidation of polyunsaturated fatty acids (PUFAs) in almost all organisms.3,4,5 Oxylipins include the phytohormone and a number of hydroxy-, oxo- or keto-fatty acids or volatile aldehydes that may perform various biological functions such as second messengers, signaling, or as antimicrobial agents.3,6 Moreover, their structural diversity may be further increased by esterification to different forms of primary metabolites such as carbohydrates and amino acids. Even several techniques have been employed in the structural elucidation of oxylipins, their mechanisms of catabolism and anabolism require further and extensive investigation. 3
Phyto-oxylipins are metabolites produced in plants by the oxidative transformation of unsaturated fatty acids via diverse metabolic pathways. Biochemical dissection and genetic approaches suggest that these oxygenated derivatives actively participate in plant defense mechanisms. Several roles have been attributed to oxylipins, namely anti-microbial activities, 7 regulation of cell death,8,9,10 control of oxidative stress, 11 and induction of defense-mediate genes.12, 13,14,15
The functional significance of plant oxylipins in carcinogenesis may involve a myriad of mechanisms. Phyto-oxylipins may exhibit anticancer effects through the regulation of apoptosis. 16 Plant oxylipins may also possess the capability to influence angiogenesis. 17 Another key mechanism may involve the regulation of inflammation. 18 Moreover, mechanisms may involve effects on tumor microenvironment and the immune response. For example, lipoxins and resolvins possess immunomodulatory properties that can enhance anti-tumor immune responses. 19 Thus, the potential biological significance of phyto-oxylipins in cancer therapy cannot be overemphasized.
In tandem with this, the chemical diversity and functional significance of oxylipins has gained significant attention. 20 Their involvement in inflammation, cell signaling, and regulation of physiological processes positions them as potential targets for cancer therapy. However, the intricate interplay between genetic and epigenetic factors in cancer development that underscores the complexity of tumorigenesis presents the need for critical insights into effective preventive and therapeutic strategies. This review seeks to explore the origins and importance of plant oxylipins, biochemical features and properties and potential mechanisms of anti-cancer properties in the context of tumorigenesis. It provides insights into carcinogenesis and the promising avenues that research into plant oxylipin can open for innovative therapeutic approaches.
Methodology
This review was based on a thorough compilation and analysis of research work carried out on biological significance and implications of plant-derived anti-cancer oxylipins. Curation of data was based on several databases and resources such as Google Scholar, Web of Science, Scopus, PubMed, DrugBank and PubChem. Key words and phrases used in the search process included ‘’carcinogenesis’’, ‘’tumorigenesis’’, ‘’anti-cancer properties of phyto-oxylipins’’, ‘’functions of oxylipins’’, ‘’anti-cancer properties of plants’’, and ‘’clinical investigations of plant oxylipins’’. In the context and guidelines of the subject matter, no exclusion and inclusion criteria were necessarily employed in the screening of articles.
Biochemical Features of Oxylipins
The defining feature of oxylipins is the presence of oxygenated functional groups within the fatty acid chains. These oxygenated groups can include hydroxyl, keto, or epoxy (cyclic ether) groups. 21 The specific type and position of these oxygenated groups vary among different oxylipin molecules. Some oxylipins contain hydroxyl or keto groups at specific carbon positions on the fatty acid chain, which can be critical for their biological activity. 22
Oxylipins may feature a cyclopentane ring structure in addition to oxygenated functional groups. 23 This ring imparts unique properties and biological activities to these molecules. Many oxylipins retain some of the double bonds present in the parent PUFAs. In some cases, the oxidation process may result in the formation of conjugated double bond systems, which can affect the molecule's reactivity and biological functions. Moreover, the length of the fatty acid chain and its degree of saturation can vary among different oxylipins. For example, prostaglandins derived from arachidonic acid typically have 20 carbon atoms in their fatty acid chains. 24 Oxylipins can exist as different isomers or structural variants, depending on the positions of oxygenated groups and substituents on the fatty acid chain.
Oxylipins may contain chiral centres thereby existing as enantiomers. These enantiomers may have different biological effects due to their stereochemistry. 25 For instance, prostaglandins contain a five-membered ring structure, with various substituents such as hydroxyl and keto groups at different positions around the ring that contribute to their diverse biological activities. Leukotrienes are characterized by a three-conjugated double bond system and contain various oxygen-containing functional groups. Jasmonic acid (JA), derived from linolenic acid, features a cyclopentanone ring and a carboxylic acid group. 12 Resolvins contain multiple hydroxyl groups and epoxy rings in their structures. 26
The biosynthesis of oxylipins is a multifaceted process that begins with the release of PUFAs from cellular membranes. 27 Lipoxygenases (LOXs) then catalyze the oxygenation of PUFAs at specific carbon atoms. The process introduces molecular oxygen into the fatty acid chain, forming hydroperoxy groups (-OOH) at specific positions. For example, 5-lipoxygenase in animals and plants oxygenates arachidonic acid to produce 5-hydroperoxyeicosatetraenoic acid. 28 Enzymes such as cyclooxygenases (COX) and prostaglandin synthase also play critical roles in prostaglandin biosynthesis. 25 Alternatively, oxylipins may be formed through non-enzymatic autoxidative processes. Phytoprostanes and isoprostanes, for instance, may result from the autoxidation of PUFAs. 29
Esterified oxylipin biosynthesis starts with the release of fatty acids from membrane lipids by the action of a lipase. 30 Initially, the free fatty acid undergoes oxidation with molecular oxygen, resulting in the formation of a hydroperoxy fatty acid. 30 This process can be facilitated by either LOX or an α-dioxygenase (α-DOX). The products generated by α-DOX spontaneously decompose into carbon monoxide and shortened aldehyde derivatives. Alternatively, they can be reduced by chemical or enzymatic means, via glutathione or peroxygenase, respectively. Conversely, the hydroperoxy products produced by LOX enzymes may exhibit relative stability and undergo further metabolism by other enzymes. These include specific members of the P450 enzyme group, known as the Cyp74-family such as allene oxide synthase, hydroperoxide lyase, divinylether synthase, peroxygenase or epoxyalcohol synthase. 30
Plants as Sources of Oxylipins
Plants serve as rich sources of oxylipins of immense chemical diversity. Most of these compounds are actively produced after wounding or pathogen attack. They have been implicated in diverse roles such as anti-microbial activities, 7 regulation of cell death,8,9,10 control of oxidative stress, 11 and induction of defense-mediate genes.12,13,14,15
Arabidopsis thaliana is a small plant species from the mustard family that is native to Eurasia and Africa. JA, linolenic acid, 13-hydroperoxylinolenic acid and 12-oxo-phytodienoic acid are all important intermediates in oxylipin pathways in Arabidopsis. 31 The signaling pathways involving JA in plants is regulated by a core of COI1/JAZs/MYC2 proteins that may also be critical in Arabidopsis.32,33,34,35 Particularly, JA may play important roles in wound-mediated expression of defense factors in Arabidopsis. 33 A chloroplast lipoxygenase was required for wound-induced JA accumulation in Arabidopsis. 32
Potatoes are tubers of the plant Solanum tuberosum, a perennial in the nightshade family Solanaceae. The major elicitor-inducible oxylipins in potato cells are 9-LOX-derived metabolites of PUFAs. 36 Typically, potato plants react to pathogen infection with increases in the amounts of the 9-LOX-derived 9,10,11- and 9,12,13-trihydroxy derivatives of linolenic acid, the divinyl ethers colnelenic acid and colneleic acid as well as 9-hydroxy linolenic acid. 36 Accumulation of these compounds was shown to be more pronounced during the interaction of potato with Pseudomonas syringae pv. maculicola, which is not pathogenic, compared to infection with Phytophthora infestans, the causal agent of late blight disease. 36 Furthermore, JA was shown to accumulate in potato leaves after infiltration with P. syringae pv. maculicola, but not after infection with P. infestans. 36
Solanum lycopersicum L is a fruit from the nightshade family native to South America. Despite botanically being a fruit, it's generally eaten and prepared like a vegetable. The oxylipin pathway in tomatoes uses free unsaturated fatty acids with the sequential action of lipoxygenase, hydroperoxide lyase and alcohol dehydrogenase to produce volatile aldehyde and alcohol compounds which contribute to fresh tomato aroma. 37 Interestingly, oxylipins are also implicated as communication signals during interactions between tomato and the root-knot nematode (Meloidogyne javanica), with (9S,10E,12Z,15Z)-9-hydroxy-10,12,15-octadecatrienoic acid (9-HOT) playing potentially crucial role. 38 Metabolism of oxylipins in tomatoes may also depend on genetically distinct signaling pathways for the regulation of different classes of wound responsive genes which involve plant-specific cytochrome P450 s allene oxide synthase and fatty acid hydroperoxide lyase. 39 Oxylipin gene expression profiles in tomato may also be altered by eicosapolyenoic fatty acids. 40
Nicotiana tabacum L is a perennial herbaceous plant which is the most commonly grown species in the Nicotiana genus. The induction of LOX-mediated pathway of oxylipins in tobacco cells was demonstrated via concentrated culture filtrate (CCF) from Phytophthora infestans and lipopolysaccharide from Pectobacterium atrosepticum. 41 Generally, oxylipin pathway was strongly induced in tobacco with CCF treatment, with colneleic acid and 9(S)-hydroxyoctadecatrienoic acid increasing up to 24 h and 16 h, respectively. 41 Moreover, alpha-dioxygenase-generated 2-hydroxylinolenic acid, and LOX-generated 9-hydroxyoctadecatrienoic acid were shown to exert a tissue-protective effect in bacterially infected tobacco leaves. 9
Physcomitrella patens is the first moss species with an available sequenced genome. 42 The species has eight genes that encode lipoxygenase of which seven are functionally active in vitro. 43 Two of these enzymes were shown to be selective towards arachidonic acid as the substrate, while the other five preferred alpha-linolenic acid. 43 Also, all of the active lipoxygenases exhibited an optimum pH at 7.0, except for one which recorded highest activity at pH 5.0. HPLC analyses also indicated that the two arachidonic acid lipoxygenases gave rise to (12S)-hydroperoxy eicosatetraenoic acid as the main product, while the other five produced (13S)-hydroperoxy octadecatrienoic acid from alpha-linolenic acid. 43
As with most land plants, macroalgae also form oxylipins via enzymatic and non-enzymatic routes. 44 Marine algae are rich and heterogeneous sources of great chemical diversity, among which oxylipins are a well-recognized class of natural products. 44 Algal oxylipins comprise an assortment of oxygenated, halogenated, and unsaturated functional groups, as well as several carbocycles that vary in ring size and position in lipid chain. 44 Kanamoto et al (2011) 45 identified cyclooxygenase gene in the red alga Gracilaria vermiculophylla and cloned it in E. coli for the production of prostaglandin F2α from arachidonic acid. The green alga Acrosiphonia coalita is found on rocks in the mid to low intertidal in protected to semi-exposed habitats in spring and early summer. 46 Acrosiphonia coalita was reported to produce an assortment of oxidized PUFAs, the smallest of which was a 10-carbon conjugated trienal with antimicrobial properties. 47 The study also reported three novel branched-chain conjugated trienals in which the aldehyde was present as a branch on a 17-carbon fatty acid chain. 47 Additionally, two novel conjugated unbranched trienone octadecanoids were also isolated and characterized in addition to a family of related epoxy-alcohols. 47
Anti-Cancer Properties of Plant Oxylipins
One of the key mechanisms by which oxylipins exhibit anticancer effects is through their influence on inflammation. Chronic inflammation is a hallmark of cancer development and progression. Through the modulation of inflammatory pathways, oxylipins can either contribute to or counteract the pro-inflammatory microenvironment conducive to cancer. 18 For instance prostaglanoids and leukotrienes induce inflammation during acute inflammatory responses, while eicosanoids sustain inflammation and regenerate hepatocytes. Pro-resolving eicosanoids resolve inflammation and return the liver to homeostasis. 48 In chronic inflammation, eicosanoids sustain inflammation and promote tumor growth. The absence of pro-resolving lipids can promote chronic inflammation while their presence may directly attenuate tumor cell growth.
Oxylipins have been implicated in the regulation of apoptosis. 16 Pro-apoptotic oxylipins, such as 15-hydroxyeicosapentaenoic acid and 13-hydroxyoctadecadienoic acid, induce apoptosis in cancer cells, inhibiting uncontrolled proliferation and survival. A growing body of evidence indicates that in contrast to the downstream ω-6 fatty acid (arachidonic acid) which has been linked to cancer development, the upstream ω-6 fatty acids [linoleic acid, γ-linolenic acid, and dihomo-γ-linolenic acid] might actually possess anti-cancer properties. As a result, these upstream ω-6 fatty acids hold promise as potential dietary elements for cancer prevention and treatment. 49 It's also worth-noting that the upstream ω-6 fatty acids may be converted into arachidonic acid through a series of enzymatic processes in fatty acid metabolism. In fact, it has been suggested that 15-LOX metabolites of docosahexaenoic acid, which is a member of the omega-3 PUFA family, reduce prostate cancer growth by converting arachidonic, eicosatrienoic, and/or oreicosapentaenoic acids to omega-6 hydroxy metabolites. 50 15-LOX-1 is overexpressed in cancerous prostate tissue and correlates positively with disease severity. It prefers linoleic acid to arachidonic acid and creates the metabolite 13-hydroxy-octadecaenoic acid. However, 15-LOX-2 prefers arachidonic acid to linoleic acid, producing the metabolite 15-hydroxy-eicosatetraenoic acid. 51 Similar investigations into the relative ability and specificity of the isoforms have been conducted in different types of cancers, although elucidation of the exact functional significance may require further studies.52,53
Furthermore, oxylipins possess the ability to influence angiogenesis that is important for tumor growth and metastasis. 17 Specific oxylipins, including epoxyeicosatrienoic acids, exert anti-angiogenic effects by inhibiting the proliferation and migration of endothelial cells, thereby obstructing the blood supply to tumors. Angiogenesis constitutes a vital element of cancer progression. 17 These findings open the door to the potential development of therapies based on oxylipins to target tumor vasculature.
Moreover, oxylipins can exert an influence on tumor microenvironment and the immune response. Oxylipins, such as lipoxins and resolvins, possess immunomodulatory properties that can enhance anti-tumor immune responses. Acetylenic oxylipins of the falcarinol-type have demonstrated covalent binding to the efflux protein ABCG2, also recognized as the breast cancer resistance protein (BCRP), which is associated with resistance to breast cancer chemotherapy. 19 The electrophilic nature of C17 and C18 acetylenic oxylipins, coupled with their capacity to react with -thiol groups in cysteine, implies their potential to activate the Kelch-like ECH-associated protein 1 (Keap1)/nuclear factor erythroid 2–related factor 2 (Nrf2)/antioxidant response element (ARE) pathway. 54 The Keap1-Nrf2 pathway plays a pivotal role in regulating the expression and formation of a spectrum of antioxidant, anti-inflammatory, and cytoprotective phase 2 enzymes. As a result, this pathway assumes significant importance in providing chemopreventive protection against carcinogens. These compounds promote the resolution of inflammation and facilitate a more effective immune response against cancer cells.
In a study aimed to determine if baseline oxylipins or PUFAs could predict who might develop symptoms related to Aromatase Inhibitor-Induced Arthralgia (AIA) in breast cancer patients, the preliminary results showed that baseline levels of 8-HETE (hydroxyeicosatetraenoic acids) were significantly related to worsening AIA symptoms from baseline to six months of adjuvant therapy with anastrozole. 55 8-HETE is primarily produced from arachidonic acid via 15-LOX and is a strong activator of peroxisome proliferator-activated receptor (PPAR) alpha and a weak activator of PPAR gamma, regulators of lipid homeostasis, and induces differentiation of preadipocytes. 56 Additionally, two linoleic acid metabolites produced via the LOX pathway (13(S)-HOTrE and 9-OxoODE) were significantly related to the development of hot flashes. 56 However, no association was found between these LOX metabolites and pain scores on treatment with anastrozole. The study suggests that additional suppression of estrogen through 8-HETE and an inflammatory profile related to overexpression of 15-LOX may predispose women to AIA.
Methyl jasmonates (MJ) are particularly selective towards malignant cells. 57 They are involved in the inhibition of multiple intracellular pathways in different cancer cell types. 58 MJ may induce reactive oxygen species which can stimulate MAPK-stress signaling pathways, cause cell cycle arrest at different phases in different types of cancer cells, promote the detachment of hexokinase from the voltage dependent-anion-channel isoform 1 in the outer mitochondrial membrane, induce cell death by triggering intrinsic/extrinsic proapoptotic and nonapoptotic pathways, inhibit metabolic enzymes such as the aldo-keto reductase 1 and 5-LOX that favor cancer cells survival and proliferation and downregulate NF-αB transcription factor activity and inhibition of the 5-LOX pathway.51,58 Integrating all these mechanisms may lead to the death of mammalian cancer cells. Table 1 highlights the biological functions of some common groups of plant oxylipins, while Figure 2 summarizes important mechanisms by which phytochemicals may exhibit anti-cancer effects.

Phytochemical-mediated control of tumorigenesis. Mechanisms may involve induction of cell death (apoptosis/autophagy), regulation of cell cycle or the inhibition of pathways such as angiogenesis, pro-inflammation and epithelial-mesenchymal transition (EMT).
Biological Functions of Common Classes of Plant Oxylipins.

Functional Implications and Future Directions of Oxylipins
Cancer may be treated by various innovative approaches. Treatment strategies such as the use of radiomics/pathomics, thermal ablation and magnetic hypothermia, gene therapy, immunotherapy, utilization of extracellular vesicles and nanomedicine may find useful applications in cancer treatment. However, the antioxidant properties of phytochemicals may also play important roles in the fight against cancer (Figure 3). In this light, it is important to pay attention to phyto-oxylipins to harness their full therapeutic potential.
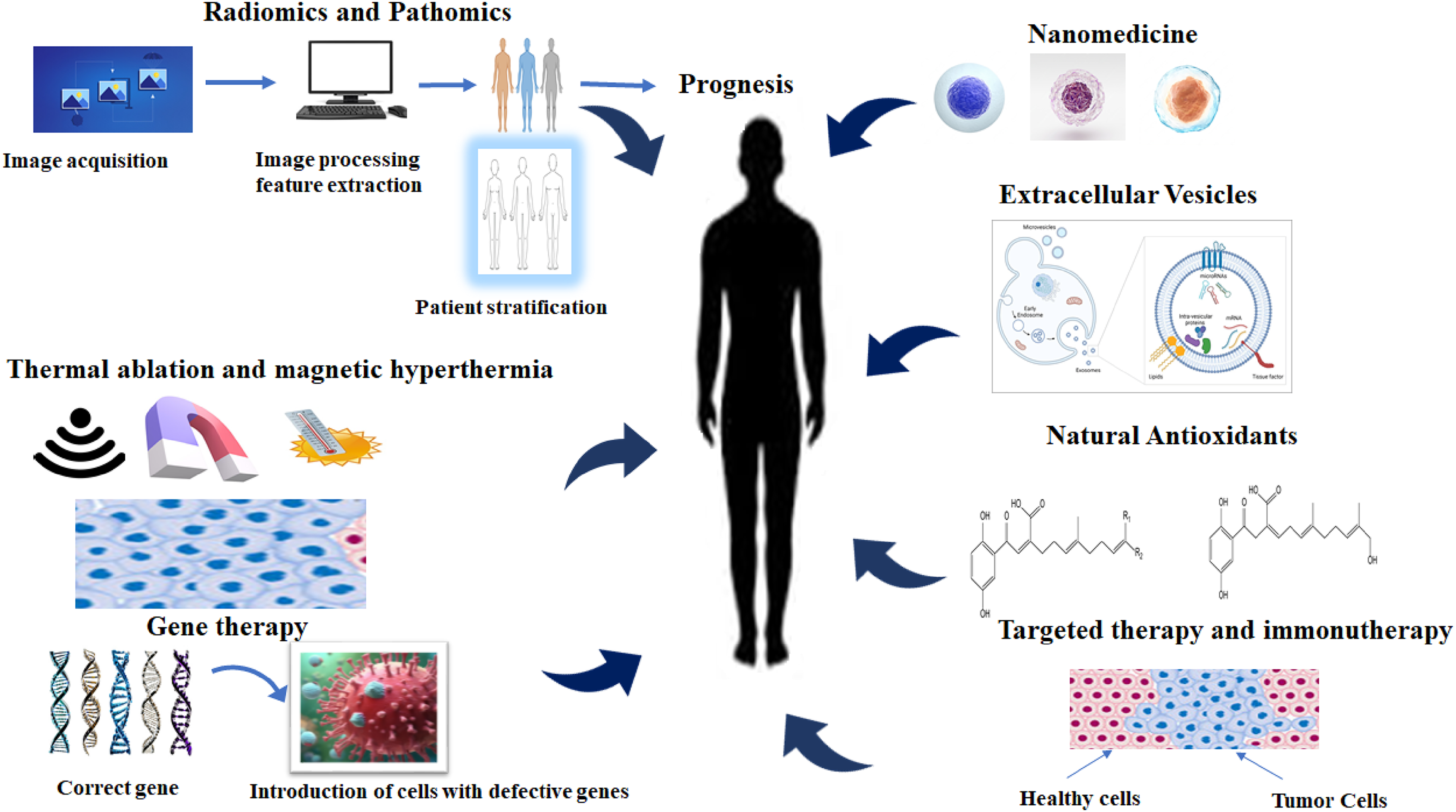
Treatment strategies in cancer. Treatment strategies may involve radiomics/pathomics, thermal ablation and magnetic hypothermia, gene therapy, immunotherapy, utilization of extracellular vesicles and nanomedicine.
Studies highlight the roles of oxylipins in the regulation of growth and development in different organisms. Current investigations employ various tools to unravel molecular mechanisms of action of oxylipins. These studies provide profound insights into oxylipin-mediated stress signal transduction, the regulation of stress-inducible gene expression, and interplay with other cellular signaling pathways. 69 Furthermore, beyond the well-known JA biosynthesis pathway mediated by 13-allene oxide synthase, plants utilize alternative pathways to transform 13-hydroperoxides into green leaf volatiles and divinyl ethers through cytochrome P450 family proteins. 70 These findings underscore the diverse roles and regulatory mechanisms of oxylipins, enriching our understanding of their biological significance.
The utilization of plant metabolites and their synthetic equivalents has facilitated the development of various antibacterial, antifungal, antiviral, anticancer, and anti-inflammatory treatments. Oxylipins, are pivotal in plant adaptation to dynamic environments and possess therapeutic promise due to their capacity to regulate physiological processes in animal cells. They exhibit immunomodulatory effects, initiate cancer cell apoptosis, and regulate the expression of pro-inflammatory cytokines. 51 Biologically active eicosanoids, such as prostaglandins, prostacyclins, thromboxanes, leukotrienes, and lipoxins play crucial roles in regulating various animal tissues and systems, participating in immunopathological processes like inflammation, autoimmune responses, and cancer. 51 Moreover, the molecular composition of oxylipins suggests their role as adaptogens. 71
Oxylipins play pivotal roles in executing pathogenic strategies for host colonization, reproduction, and toxin production. For example, jasmonates act as signaling molecules in pathways activated by pathogens and insects, subsequently regulating the expression of genes implicated in defense mechanisms. 72 Oxylipins such as 13-keto-9(Z),11(E),15(Z)-octadecatrienoic acid and 12-oxo-10,15(Z)-phytodienoic acid were shown to strongly inhibit mycelial growth and spore germination of eukaryotic microbes. 7 In the same study, regulators of plant defense responses were also shown to be the most active oxylipins against eukaryotic microorganisms. 7 Additionally, a distinct array of oxylipins generated through the chloroplast-localized hydroperoxide lyase-mediated pathway, such as the green leaf volatiles (GLVs) may exhibit antimicrobial properties and act as signaling molecules in defense responses. 73
The utilization of phyto-oxylipins presents opportunities for developing innovative products with wide-ranging applications across industries. Recent focus on oxylipins has been linked with the progress of advanced technological platforms, particularly mass spectrometry techniques such as LC-MS/MS and GC-MS/MS. 74 There are notable improvements in the methodologies for extracting oxylipins from biological samples. 74 Moreover, the distinctive chemical structures and biological functionalities of these compounds render them as promising candidates for biotechnological advancements. Traditional analytical methods have been supplanted by state-of-the-art approaches employing mass spectrometry coupled with liquid chromatography and gas chromatography. 75 These advanced methods facilitate the quantitative detection of oxylipins. 75
Phyto-oxylipins could be utilized for generating bioactive components in functional foods or nutraceuticals aimed at enhancing human health. Additionally, the prospective utilization of phyto-oxylipins in agriculture holds potential for augmenting crop productivity and resilience to environmental pressures. Leveraging insights derived from studying plant oxylipin pathways, biotechnologists can explore methodologies to engineer crops with enhanced stress tolerance or improved nutritional compositions. Moreover, oxylipins may potentially represent a novel frontier in the identification of risk indicators and therapeutic targets across various diseases, particularly cardiovascular diseases (CVDs). A deeper understanding of their functional roles in medical and biotechnological interventions is thus required.
Conclusion
The functional implications of plant oxylipins in plant biology, human health, and agricultural innovation are elucidated in the present study. By investigating the mechanisms of these functions and harnessing their bioactive properties, new opportunities for scientific discovery, medical treatment, and biotechnological advancement can be unlocked. Moreover, much of the potential of phyto-oxylipins as therapeutic agents for cancer treatment remains to be fully established. Furthermore, clinical investigations are needed to determine optimal concentrations and effective delivery methods. The potential cytotoxic effects of oxylipins on normal cells and tissues should also be carefully evaluated. Overall, research into phyto-oxylipins require significant attention due to the promise it may hold in addressing key challenges in biotechnology, health, and environmental sustainability.
Footnotes
Abbreviations
Acknowledgments
I am grateful to colleagues and faculty members at the Department of Biological Sciences, University of Environment and Sustainable Development, PMB, Somanya, Ghana, for their motivation during the preparation of this manuscript.
Data Availability
All data used or generated from this study can be found in this manuscript.
Declaration of Conflicting Interests
The author declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author received no financial support for the research, authorship, and/or publication of this article.