Natural products and their derivatives have served as powerful therapeutics against pathogenic microorganisms and are the mainstay of our currently available treatment options to combat infections. As part of our ongoing search for antimicrobial natural products from marine organisms, one fraction prepared from the Arctic marine bryozoan Securiflustra securifrons was found to be active against the human pathogenic bacterium Streptococcus agalactiae (gr. B). Chemical investigation of the fraction revealed that it contained several variants of the highly modified secondary metabolites known as securamines. The securamines are alkaloids sharing a common isoprene-histamine-tryptamine backbone. In this study, we describe the antimicrobial activities of securamine C, E, and H – J (4, 5, and 1-3) and the attempt to deconvolute the mode of action of 1.
The increasing prevalence of antibiotic resistant pathogens is recognized as one of the most serious global threats to human health in the 21st century. Extensive use of antimicrobials together with declining investments into discovery and development of new treatment options to combat pathogens have aggravated the problem.1 Novel classes of antibiotics are therefore urgently needed for the future. In the search for new antimicrobial agents, nature remains the richest and most versatile source.2,3 In fact, close to 80% of all marketed anti-infective agents originate from a natural source.4
Bryozoans are a phylum of suspension-feeding mainly colonial invertebrates found in aquatic benthic ecosystems throughout the world.5 They generally form sessile colonies of genetically identical, polymorphic units termed zooids.6,7 These colonies are extremely vulnerable to biofouling, predation by grazers and pathogenic attacks.8,9 In order to thrive in this hostile environment, bryozoans have developed a chemical defense strategy, in which potent secondary metabolites are produced to combat these external threats. As a result, bryozoan biomass has yielded several structurally diverse bioactive secondary metabolites. The best known examples are the bryostatins, isolated from Bugula neritina (Linnaeus, 1758), some of which are under clinical development as an anticancer drug candidate for combination therapy10 and neurological disorders.11-13
As part of our ongoing search for bioactive secondary metabolites from marine organisms, the organic extract of Securiflustra securifrons (Pallas, 1766) was prepared into eight fractions and tested for antibacterial activity. Fraction three was found to be active against the pathogenic bacterium Streptococcus agalactiae (Gr. B). Previously, fraction five of the same extract was found to be active against a human melanoma cancer cell line.14,15 The components found to be responsible for the cytotoxicity in this fraction were the hexacyclic alkaloids securamine C (4), E (5), and H – J (1-3) (Figure 1). The structures and cytotoxic properties of 1-5 have been reported by our group.14 HRMS analysis of the fraction showing antibacterial activity revealed the presence of securamines. The antimicrobial properties of the securamines have not been previously examined. This, coupled with the knowledge that nature has provided a wealth of promising lead structures for antimicrobial development,16-18 motivated an investigation into the antimicrobial potential of the securamines.
Structures of securamine C, E, and H – J (4, 5, and 1-3) isolated from the organic extract of the Arctic marine bryozoan Securiflustra securifrons.
Results and Discussion
Biomass, Extraction, Fractionation, Compound Isolation, and Structure Elucidation
The biomass of S. securifrons was collected off the coast of Hjelmsøya, freeze dried and subjected to liquid-liquid extraction, providing an aqueous and an organic extract. The organic extract was fractionated into eight fractions using flash chromatography. Compounds 1-5 were isolated using mass guided semi-preparative HPLC and their structures elucidated using spectroscopic methods (HRMS, 1D- and 2D-MNR), as previously described.14
Antibacterial Screening and Chemical Investigation of the Flash Fractions of S. securifrons
The flash fractions of the organic S. securifrons extract were assayed for activity against the pathogenic bacterial strains S. aureus, E. faecalis, E. coli, P. aeruginosa, and S. agalactiae (Gr. B) at 250 µg/mL. Flash fraction three (eluting at 50% MeOH) was found to be active against the G+ bacterium s. agalactiae (Gr. B). Fraction three was inactive against the remaining bacteria and the remaining fractions were inactive against all bacteria. Chemical analysis of the fraction using UHPLC-HRMS revealed that it contained compounds belonging to the securamine family, including compounds 1, 2, 4, and 5.
Antimicrobial Activity of 1-5
The MIC values of compounds 1-5 were determined against four G+ and two G- bacterial strains, three yeast strains and toward the biofilm formation capability of S. epidermidis (Table 1, online supplementary file). Compounds 1, 2, and 5 showed activity against all or most of the G+ strains. No activity was found toward the G- bacteria, the yeast strains or against biofilm formation at the highest assay concentration (50 µM).
MIC (µM) of Securamine H-J, C, and E (1-5) Against four G+ and two G- Bacterial Strains, Against Staphylococcus epidermidis Biofilm Formation, and Against three Yeast Strains.
Microorganisms
Minimum inhibitory concentration (µM)
1
2
3
4
5
G+ bacteria:
Bacillus subtilis
6.25
>50
>50
>50
>50
Staphylococcus aureus
3.13
12.5
>50
>50
25
Enterococcus faecalis
6.25
25
>50
>50
50
Streptococcus agalactiae (Gr. B)
6.25
25
>50
>50
25
G- bacteria:
Escherichia coli
>50
>50
>50
>50
>50
Pseudomonas aeruginosa
>50
>50
>50
>50
>50
Biofilm formation:
Staphylococcus epidermidis
>50
>50
>50
>50
>50
Yeast:
Candida albicans
>50
>50
>50
>50
>50
Rhodotorula sp.
>50
>50
>50
>50
>50
Aureobasidium pullulans
>50
>50
>50
>50
>50
Based on these results it appears that a double bond in the D-ring between C-2 and C-3 and more than one bromine on the A-ring at C-16, C-17, and/or C-18 are important for the antibacterial activity of the securamines. This observed structure activity relationship (SAR) correlates well with the previously reported SAR of the securamines against melanoma, lung and breast cancer cell lines,14 where 1, 2, 4, and 5 inhibited cell viability. The lowest MIC value, 3.13 µM, was measured for 1 against S. aureus. Compound 1 is brominated at C-16, C-17, and C-18, and has a double bond between C-2 and C-3. This could indicate that the activities of the securamines against both bacteria and cancer cell lines are caused by an unspecific interaction with and disruption of biological membrane integrity. This is, however, unlikely, as the herein assayed eukaryotic yeast strains were unaffected by the securamines at the highest assayed concentration (50 µM), indicating an unrelated intracellular target in the cancer- and bacteria cells. Compound 1 showed the broadest and most potent inhibition, with MIC values ranging from 3.13 to 6.25 µM against the G+ bacteria, and was, therefore, chosen for further investigation.
Real-Time Measurement of Membrane Integrity of Bacteria When Exposed to 1
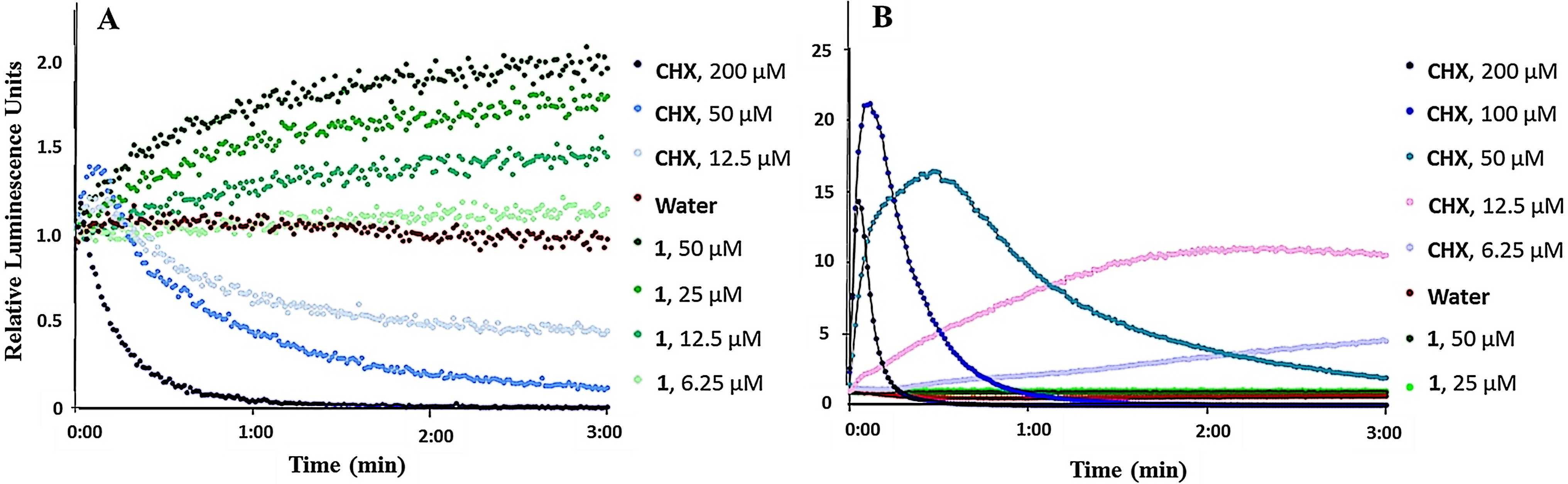
B. subtilis and E. coli, carrying the pCSS962 plasmid with the LucGR gene, were used to assess the membrane disruptive properties of 1. The strains express eukaryotic luciferase and will emit luminescence if their membrane is disrupted and D-luciferin from the growth medium is allowed to diffuse into the cell.19 If the bacterial cells die following membrane disruption, an initial rise in relative luminescence units (RLU) caused by D-luciferin influx, will be followed by declining RLU values as bacterial ATP reserves are exhausted and the enzymatic reaction consequently stopped. The luminescence measurements of B. subtilis and E. coli after exposure to ranging concentrations of 1 or chlorhexidine (CHX, positive control), an antibiotic known for its cell wall and membrane-disruptive activities,20 can be seen in Figure 2. CHX treated B. subtilis gave an initial increase followed by a decrease in RLU values. In contrast to this, 1 caused a persistent increase in light emission from the cells within the 3 minutes assay time at the 3 hours highest concentrations tested (12.5, 25, and 50 µM; 2, 4, and 8 × the MIC value of 6.25 µM, respectively). This increase was most likely caused by an increased D-luciferin influx into the cells caused by effects on the membrane. This effect does however not appear to affect the viability of B. subtilis, as the ATP reserves in the bacteria are not exhausted and resultantly no delayed drop in RLU was observed, indicating that the effect of 1 was different from that of CHX. The lack of a drop in ATP reserves following B. subtilis exposure to 1 furthermore indicates that 1 is bacteriostatic rather than bactericidal at the highest assayed concentrations. At 6.25 µM (and at lower concentrations, data not shown) 1 did not cause an influx of D-luciferin, as relative light emission remained equal to the water control. In contrast to CHX, no substantial increase in D-luciferin uptake and thus no effect on plasma membrane integrity of E. coli could be detected for 1, even at the highest assayed concentrations (Figure 2(B)).
Kinetics of the relative luminescence emission by A) B. subtilis (pCSS962) and B) E. coli (pCSS962) treated with ranging concentrations of either securamine H (1) or chlorhexidine (CHX). Each point is the mean of three independent measurements. CHX and water were used as positive and negative controls, respectively.
Assessment of the Membrane Potential of Bacteria When Exposed to 1
To elucidate further if 1 affected the membrane integrity of B. subtilis directly, the membrane potential of B. subtilis was measured after 3 minutes exposure to ranging concentrations of 1. Bacterial cells were stained with a membrane potential sensitive dye and subsequently analyzed by flow cytometry. The assay is based on the use of 3,3’-diethylcarbocyanine iodide (DiOC2(3)), the fluorescence of which shifts from green to red in response to higher cytosolic concentrations in cells with active membrane potential where the dye aggregates. Ratiometric analysis of green to red fluorescence allows for estimating changes in membrane potential of bacteria.21 CHX and carbonyl cyanide m-chlorophenylhydrazone (CCCP) were used as positive and negative controls, respectively. CCCP blocks the generation of the electrochemical proton gradient, and thus lowers the membrane potential in bacteria.22 Increasing concentrations of CHX decreased the ratio of red/green, showing that higher CHX concentrations result in a higher fraction of bacteria with disrupted membranes. For 1, increased concentrations resulted in a decreased red/green ratio, but this decrease was significantly less marked compared to the CCCP and CHX controls. At the observed MIC (6.25 µM), only a slight shift was visible, further indicating that the activity of 1 was not due to direct membrane integrity disruption (Figure 3).
The effect of securamine H (1) in comparison to chlorhexidine (CHX) on the membrane potential of B. subtilis. The bacteria were treated for 3 minutes with ranging concentrations of either 1 or CHX and subsequently incubated for at least 30 minutes with 30 µM DiSO2(3). The overlaid histograms show the positive and the negative controls treated with 5 µM carbonyl cyanide m-chlorophenylhydrazone (CCCP) (shaded dark grey) and water (shaded light grey), respectively. Measurements depicting analysis of 1 are highlighted. The ratiometric values (red/green) are depicted on the x axis, and the relative number of events on the y axis.
Effect of 1 on Bacterial Metabolism
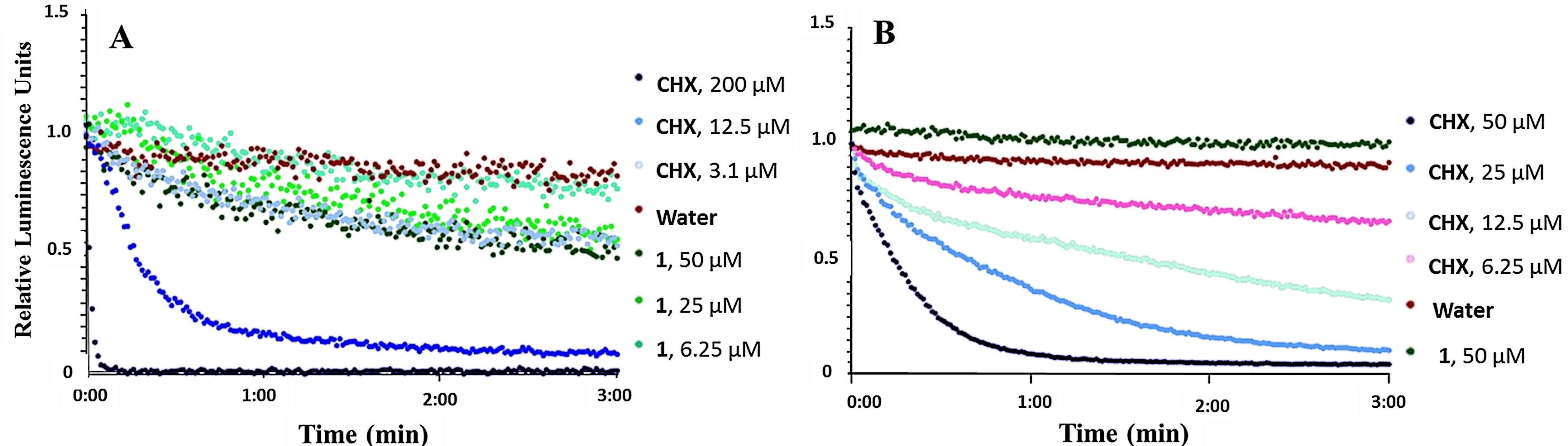
We proceeded to evaluate whether 1 affected bacterial metabolism. B. subtilis carrying a chromosomal integration of a luxABCDE23 operon and E. coli carrying the pCGLS-1124 plasmid with a Photorhabdus luminescens lux operon (luxCDABE) were used to assess the effect of 1 on bacterial cell viability measured in real-time. From these operons, the strains express a bacterial luciferase and fatty acid reductases for regeneration of long-chain fatty aldehydes, which serve as substrates for light production. Light production is therefore linked to several metabolic processes, which in turn depend on the regeneration of reduction equivalents and ATP.25,26 While light production indicates active metabolism, loss of light production indicates a decrease in metabolic activity, and hence, reduced viability of the cells. The measured luminescence of B. subtilis and E. coli after addition of ranging concentrations of either 1 or CHX can be seen in Figure 4. At concentrations above the MIC (6.25 µM), 1 affected the viability of B. subtilis within the 3 minutes assay time (Figure 4(A)). Indeed, 1 elevated light emission by the strain at a similar level to CHX at 3.1 µM, resulting in a decrease of around 40% of relative luminescence units after 3 minutes. The decrease in light emission within 3 minutes at concentrations above the MIC confirmed that cell viability was affected relatively fast. However, even at concentrations above the MIC, viability does not drop below 50%, which was the case for CHX. In addition, the ATP dependent membrane assay showed elevated light emission at these concentrations indicating that ATP levels are not the limiting factor. No effect was observed toward E. coli (Figure 4(B)).
Kinetics of the relative luminescence emission by A) B. subtilis (pCGLS-11) and B) E. coli (pCGLS-11) treated with ranging concentrations of either securamine H (1) or chlorhexidine (CHX).
In summary, the results from the membrane integrity and metabolic activity assays show an influx of D-luciferin at concentrations higher than the MIC, no effect on membrane potential, and reduction of metabolic activity in B. subtilis exposed to 1. The effect of 1 on B. subtilis viability is thus most likely not caused by direct effects on the cell membrane, but rather by interference with one or more metabolic processes in B. subtilis.
Investigation of Possible Intracellular Targets of 1
In an attempt to gather information regarding the mode of action of 1, the compound was tested against a panel of six biosensors responding to interference with some major metabolic pathways in B. subtilis. Compound 1 was tested for interference with DNA replication, transcription, translation and interference with fatty acid, cell wall and folic acid synthesis. Antibiotics of the respective modes of action, as shown in Table 2, served as positive controls. None of the strains reacted to co-incubation with 1 at the assayed concentrations during the 8 hours the assays were run (twofold dilution series between 50 and 0.39 µM). The purpose of the sensors is to detect similar activity, but negative results do not exclude a given mode of action. While, for example, erythromycin efficiently induces the sensor for translation interference, kanamycin does not.27 Similarly, the cell envelope stress sensor, which is based on the liaI promoter and the liaRS two-component system, is most sensitive when challenged with antibiotics affecting the lipid II cycle, as is the case for bacitracin and vancomycin, while they remain uninduced by penicillin.28 This hampers the interpretation of the results since all the assays for the activity of 1 were negative. However, 1 seems not to belong to the subgroups of antibiotic mode of actions that the sensors recognize. Interestingly, the liaI based cell envelope stress sensor is known to respond to membrane active compounds such as nisin.28 Therefore, the negative response to 1 was in accordance with earlier results indicating a different mode of action than interference with membrane integrity.
List of Sensor Strains With the Promoter Region Fused to the luxABCDE Operon Located in the sacA Region of the B. subtilis 168 Chromosome. Respective Positive Control Antibiotics Used Are Shown in the Last Column.
Strain number
Target
Promoter
Control antibiotic
EM10
DNA replication
yorB
Ciprofloxacin
EM11
Transcription
helD
Rifampicin
EM12
Translation
yheI
Erythromycin
HMB67
Wall and membrane
liaI
Vancomycin/Bacitracin
HMB69
Fatty acid synthesis
fabHB
Triclosan/Irgasan
HMB70
Folic acid synthesis
panB
Trimethoprim
HMB62
Viability control
laiG
all
Conclusions
Securamine H (1) was found to inhibit the viability of G+ bacteria and to reduce metabolic activity in B. subtilis. The effect was shown not to be caused by interference with DNA replication, transcription, or translation, nor by interference with fatty acid, cell wall and folic acid synthesis, and could not be explained by disruption of the cell membrane. The mode of action of 1 thus remains to be deconvoluted. However, as the result indicates that 1 has an intracellular target, the compound serves as an interesting starting point for further investigations. The herein presented results demonstrate that marine bryozoans can be used as a source of compounds with antibacterial activity.
Experimental
Animal Material, Extraction, Fractionation, Compound Isolation, and Structure Elucidation
Specimens of S. securifrons were collected off the coast of Hjelmsøya, Norway in 2014 using an Agassi’s dredge trawl at 72 m depth. The specimens were prepared as an organic extract, which was further fractionated into eight fractions using RP-flash chromatography and tested for bioactivity. The compounds were isolated using mass-guided semi-preparative HPLC and their structure elucidated using spectroscopic methods (HRMS, 1D- and 2D-NMR), all as previously described in detail.14
Microorganism Strains, Growth Media, and Assay Temperature
Enterococcus faecalis (ATCC: 29212) and Streptococcus agalactiae (Gr. B) (ATCC: 12386) were grown and assayed in brain-heart infusion broth (BHI; Oxoid, Hampshire, England). Staphylococcus aureus (ATCC: 25923), Escherichia coli (ATCC: 25922), Pseudomonas aeruginosa (ATCC: 27853), and Bacillus subtilis (ATCC: 23857) and its derivatives were grown and assayed in Mueller Hinton Broth (MH; Merck, Darmstad, Germany). Staphylococcus epidermidis (ATCC: 35984) was grown and assayed in tryptic soy broth (TS; Merck, Darmstadt, Germany). All bacteria were grown and assayed at 37 °C. The yeast strains Candida albicans (ATCC: 10231) and Rhodotorula sp. and the Aureobasidium pollulans mold (Rhodotorula sp and A. pollulans were obtained from Professor Arne Tronsmo, the Norwegian College of Life Sciences, Ås, Norway), were cultivated and assayed on potato dextrose agar (PD, Difco, Detroit, MI, USA) with 2% D(+)-glucose (Merck, Darmstad, Germany) at room temperature.
Antimicrobial and Anti Biofilm Formation Assays
Antibacterial assay
MIC values of 1-5 were determined using the broth microdilution method, as previously described.15 Briefly, suspended bacteria in log phase were added to 96-well microtiter plates at a concentration of 1,500‐15,000 colony forming units/mL. Serial dilutions of 1-5 (assay cons: 50‐0.78 µM) were subsequently added and left to inoculate for 24 hours before growth inhibition was measured using a Victor multilabel counter (Perkin Elmer, Singapore) at 600 nm. Growth medium diluted with water (1:1) was used as negative control and bacteria suspension diluted with water (1:1) as positive control. For B. subtilis, oxytetracycline was used as the positive assay control, and for the remaining strains, gentamycin was used. The assays were repeated three times.
Inhibition of biofilm formation
S. epidermidis was used to assess the effect of 1-5 on biofilm formation. An overnight culture of S. epidermidis was diluted with fresh TS broth with 1% glucose (1:100), transferred to the wells of 96-well microtiter plates, and ranging concentrations (assay cons: 50‐0.39 µM) of 1-5 were added. After overnight incubation, the bacterial suspension was carefully discarded, the biofilm fixed by incubation at 55 °C for 1 hour and stained with 0.1% crystal violet for 5 minutes before being washed away with water. The plates were once more left to dry at 55 °C for 1 hour before 70 µL 70% ethanol was added to each well and the plates left to incubate for 10 minutes before biofilm formation was observed by visual inspection of the plates. The MIC was defined as the lowest concentration where no biofilm formation was visible. S. epidermidis suspension, diluted with 50 µL of water, was used as a positive control, and 50 µL Staphylococcus haemolyticus suspension with 50 µL of water as negative control. A mixture of 50 µL water and 50 µL TS broth was used as assay control. The assay was repeated three times.
Antifungal assay
Fungal spores of yeast strains Candida albicans and Rhodotorula sp. and the A. pollulans mold were added to PD broth and the cell concentration determined and adjusted after counting in a Bürker chamber. A final fungal spore concentration of 2 × 105 spores/mL was inoculated in 96-well Nunc microtiter plates (100 µL total well volume) along with ranging concentrations of 1-5 (assay cons: 50‐0.78 µM). The assay plates were incubated at room temperature for either 24 hours (C. albicans) or 48 hours (A. pollulans and Rhodotorula sp.). Ranging concentrations of amphotericin-B was used as positive control (32-0.25 µg/mL), and water as a negative (growth) control. After incubation, the OD value (600 nm) was measured in a Synergy H1 Hybrid Reader (BioTek, Winooski, VT, USA). MIC values of 1-5 were defined as the lowest concentration of the compounds that showed >90% inhibition compared to the negative growth control (as measured by OD). The assays were repeated three times.
The real-time membrane integrity assay was performed using B. subtilis 168 and E. coli K12 (MC1061), both carrying the plasmid pCSS962 with the eukaryotic luciferase gene lucGR. Luciferase is dependent on D-luciferin as substrate to emit light, a substrate that does not penetrate intact cell membranes. The assay is a modification of a previously described protocol.19B. subtilis and E. coli were cultured overnight in MH broth with 5 µg/mL chloramphenicol (Merck KGaA, Darmstad, Germany) and a mixture of 20 µg/mL chloramphenicol/100 µg/mL ampicillin, respectively, before being centrifuged at 4500 rpm for 10 minutes. The supernatant was removed and the pellet resuspended in MH broth to give an OD600 of 0.1. D-luciferin potassium salt (assay concentration: 1 mM) was added and background luminescence measured. Ranging concentrations of 1 (assay conc.: 50‐0.78 µM) dissolved in water were added to black round-bottom 96-well microtiter plates (Nunc, Roskilde, Denmark), as well as a ranging concentrations of the control, CHX acetate (assay conc.: 50 µM-1.6 µM). The plates were placed in a Synergy H1 Hybrid Reader (BioTek, Winooski, VT, USA). Aliquots (90 µL, to give a total assay volume of 100 µL) of the prepared bacterial suspension were added to the test wells by an automatic injector with tracking of the luminescence emission every second for 180 s. The assays were repeated 3 times.
Membrane potential assay
To analyze the effects of 1 on the membrane potential of Bacillus subtilis 168, the BacLight bacterial membrane potential kit (Invitrogen, Carlsbad, CA, USA) was used. The assays were performed in 96 well 1.8 ml deepwell-plates (Corning, Corning, NY, USA) and analyzed by a Cube8 flow cytometer with an auto sampler (Sysmex, Kobe, Japan). The assay is based on the dye DiOC2(3), which causes green fluorescence in all bacterial cells. The fluorescence shifts to red when the dye molecules self-associate due to their accumulation in the cytoplasm of viable bacteria with intact membrane potential.21 Differences in fluorescence emission were detected by flow cytometry. The ratiometric values of red/green fluorescence were used to analyze if the proton gradient of the tested cells was affected or not. The experiment was performed according to the manufacturer’s suggestions, but adapted to the 96 well format and therefore conducted with reduced volumes. Briefly, 5 µl 3 mM DiOC2(3) solution was added to 500 µl cell suspension, which contained approximately 106 bacteria and had been pretreated for 3 minutes with different concentrations of the respective analytes. Before starting the measurement, the samples were incubated in the dark for 30 minutes at room temperature. The samples were then measured in the Cube8 and analyzed with excitation by the blue laser (488 nm) and forwards scatter (FSC), side scatter (SSC), FL1 (emission 536/40 nm) and FL3 (emission 675/20 nm). Data analysis was performed by the freely available flowing software using the first 2000 events of the bacterial population in each measurement for ratiometric analysis (flowingsoftware.btk.fi).
Real-time cell viability assay
The real-time cell viability assay was performed using B. subtilis 168 (ATCC: 23857) and E. coli K12 (ATCC: MC1061) carrying either a chromosomal integration of the luxABCDE operon or the plasmid pCGLS-11 with the lux operon luxCDABE, respectively. The assay is a modification of a previously described protocol.29B subtilis and E. coli were cultured overnight in MH broth with 5 µg/mL chloramphenicol and 100 µg/mL ampicillin (Merck KGaA, Darmstad, Germany), respectively, before being centrifuged at 4500 rpm for 10 minutes. The supernatant was removed and the pellet resuspended in MH broth to give an OD600 of 0.1. Ranging concentrations of 1 (assay conc.: 50‐0.78 µM) dissolved in water were added to black round-bottom 96-well microtiter plates (Nunc, Roskilde, Denmark), as well as ranging concentrations of the control, CHX acetate (assay conc.: 50 µM – 1.6 µM). The plates were placed in a Synergy H1 Hybrid Reader (BioTek, Winooski, VT, USA). Aliquots (90 µL, to give a total assay volume of 100 µL) of the prepared bacterial suspension were added to the test wells by an automatic injector with tracking of the luminescence emission every second for 180 s. The assays were repeated three times.
Whole cell mode of action, specific biosensors were used to determine if the activity of 1 correlates with some previously known modes of actions. The biosensors were generated by cloning promoter fusions to luxABCDE in the plasmid pBS3Clux and subsequent recombination into the sacA site on the B. subtilis chromosome, as described.23 Interference with DNA replication, transcription, translation and fatty acid synthesis was tested by B. subtilis strains containing luxABCDE fusions to the promotors of the genes yorB, helD, yheI and fabHB, respectively, as described for firefly luciferase.27 Inhibition of cell wall synthesis was tested by a bacitracin inducible promoter construct described23 based on the promotor of liaI fused to luxABCDE. The veg promoter fusion described in the same paper was used as a luminescence control. In addition, a panB-luxABCDE promoter fusion described as a lacZ-fusion in patent US20020164602A130 was used to test for inhibition of folic acid synthesis (details in Table 2). Compound 1 was tested in a two-fold dilution series starting with two x B. subtilis MIC (MIC = 6.25 µM). The respective control antibiotics were set up similar to the tested compound. The experiments were run at room temperature. Otherwise, the identical setup to the antimicrobial assay protocol was used with additional measurement of luminescence every 15 minutes for 8 hours. Peak luminescence of the controls was compared to luminescence of cells treated with 1. The assays were repeated three times.
Supplemental Material
online supplementary file 1 - Supplemental material for Antimicrobial Activity of Securamines From the Bryozoan Securiflustra securifrons
Supplemental material, online supplementary file 1, for Antimicrobial Activity of Securamines From the Bryozoan Securiflustra securifrons by Kine Ø. Hansen, Ida K. Ø. Hansen, Céline S. Richard, Marte Jenssen, Jeanette H. Andersen and Espen H. Hansen in Natural Product Communications
Footnotes
Acknowledgments
The authors are grateful for the help received by Dr. Hans-Matti Blencke in the mode of action studies and the preparation of this manuscript. We are grateful to Marbank for collecting the specimens of S. securifrons and to R. Johansen for taxonomic identification of the bryozoan.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The publication fee was covered by the open access publishing fund, UiT.
ORCID iDs
Kine Ø. Hansen
Céline S. Richard
Espen H. Hansen
Supplemental Material
Supplemental material for this article is available online.
References
1.
MedernachRL.LoganLK. The growing threat of antibiotic resistance in children. Infect Dis Clin North Am. 2018;32(1):1-17.doi:10.1016/j.idc.2017.11.00129406971
2.
NewmanDJ.CraggGM. Natural products as sources of new drugs from 1981 to 2014. J Nat Prod. 2016;79(3):629-661.doi:10.1021/acs.jnatprod.5b0105526852623
3.
GenilloudO. The re-emerging role of microbial natural products in antibiotic discovery. Antonie Van Leeuwenhoek. 2014;106(1):173-188.doi:10.1007/s10482-014-0204-624923558
4.
NewmanDJ.CraggGM.SnaderKM. Natural products as sources of new drugs over the period 1981-2002. J Nat Prod. 2003;66(7):1022-1037.doi:10.1021/np030096l12880330
5.
McKinneyFK.JacksonJBC. Bryozoan evolution. University of Chicago Press. 1991;1(1):1-252.
6.
SimpsonC.JacksonJBC.Herrera-CubillaA. Evolutionary determinants of morphological polymorphism in colonial animals. Am Nat. 2017;190(1):17-28.doi:10.1086/69178928617632
7.
KutyumovVA.MaltsevaAL.KotenkoN.OstrovskyAN. Functional differentiation in bryozoan colony: a proteomic analysis. Tsitologiia. 2016;58(1):152-159.doi:10.1134/S1990519X1602007327220253
8.
FiguerolaB.Angulo-PrecklerC.Núñez-PonsLet al. Experimental evidence of chemical defence mechanisms in Antarctic bryozoans. Mar Environ Res. 2017;129:68-75.doi:10.1016/j.marenvres.2017.04.01428487162
9.
GrayCA.McQuaidCD.Davies-ColemanMT. A symbiotic shell-encrusting bryozoan provides subtidal whelks with chemical defence against rock lobsters. Afr J Mar Sci. 2005;27(3):549-556.doi:10.2989/18142320509504115
10.
NewmanDJ.CraggGM. Marine natural products and related compounds in clinical and advanced preclinical trials. J Nat Prod. 2004;67(8):1216-1238.doi:10.1021/np040031y15332835
NelsonTJ.SunM-K.LimCet al. Bryostatin effects on cognitive function and PKCɛ in Alzheimer’s disease phase IIA and expanded access trials. J Alzheimers Dis. 2017;58(2):521-535.doi:10.3233/JAD-17016128482641
13.
KollárP.RajchardJ.BalounováZ.PazourekJ. Marine natural products: bryostatins in preclinical and clinical studies. Pharm Biol. 2014;52(2):237-242.doi:10.3109/13880209.2013.80410024033119
14.
HansenKine Ø.IsakssonJ.BayerA.JohansenJA.AndersenJH.HansenE. Securamine derivatives from the Arctic bryozoan Securiflustra securifrons. J Nat Prod. 2017;80(12):3276-3283.doi:10.1021/acs.jnatprod.7b0070329220180
15.
MichaelP.HansenKine Ø.IsakssonJ.AndersenJH.HansenE. A novel brominated alkaloid securidine A, isolated from the marine bryozoan Securiflustra securifrons. Molecules. 2017;22(7):1236.doi:10.3390/molecules2207123628737700
16.
ClardyJ.FischbachMA.CurrieCR. The natural history of antibiotics. Curr Biol. 2009;19(11):437-441.doi:10.1016/j.cub.2009.04.00119515346
17.
FairRJ.TorY. Antibiotics and bacterial resistance in the 21st century. Perspect Medicin Chem. 2014;6(6):25-64.doi:10.4137/PMC.S14459
18.
WencewiczTA. New antibiotics from Nature’s chemical inventory. Bioorg Med Chem. 2016;24(24):6227-6252.doi:10.1016/j.bmc.2016.09.014
19.
VirtaM.AkermanKE.SavirantaP.Oker-BlomC.KarpMT. Real-time measurement of cell permeabilization with low-molecular-weight membranolytic agents. J Antimicrob Chemother. 1995;36(2):303-315.doi:10.1093/jac/36.2.3038522460
20.
KuyyakanondT.QuesnelLB. The mechanism of action of chlorhexidine. FEMS Microbiol Lett. 1992;100(1-3):211-215.doi:10.1111/j.1574-6968.1992.tb05705.x1335944
21.
NovoD.PerlmutterNG.HuntRH.ShapiroHM. Accurate flow cytometric membrane potential measurement in bacteria using diethyloxacarbocyanine and a ratiometric technique. Cytometry. 1999;35(1):55-63.doi:10.1002/(SICI)1097-0320(19990101)35:1<55::AID-CYTO8>3.0.CO;2-210554181
22.
KasianowiczJ.BenzR.McLaughlinS. The kinetic mechanism by which CCCP (carbonyl cyanide m-chlorophenylhydrazone) transports protons across membranes. J Membr Biol. 1984;82(2):179-190.doi:10.1007/BF018689426096547
23.
RadeckJ.KraftK.BartelsJet al. The Bacillus BioBrick Box: generation and evaluation of essential genetic building blocks for standardized work with Bacillus subtilis. J Biol Eng. 2013;7(1):29.doi:10.1186/1754-1611-7-2924295448
24.
FrackmanS.AnhaltM.NealsonKH. Cloning, organization, and expression of the bioluminescence genes of Xenorhabdus luminescens. J Bacteriol. 1990;172(10):5767-5773.doi:10.1128/JB.172.10.5767-5773.19902211511
25.
GalluzziL.KarpM. Whole cell strategies based on lux genes for high throughput applications toward new antimicrobials. Comb Chem High Throughput Screen. 2006;9(7):501-514.doi:10.2174/13862070677793535116925511
26.
GalluzziL.KarpM. Intracellular redox equilibrium and growth phase affect the performance of luciferase-based biosensors. J Biotechnol. 2007;127(2):188-198.doi:10.1016/j.jbiotec.2006.06.01916891024
27.
UrbanA.EckermannS.FastBet al. Novel whole-cell antibiotic biosensors for compound discovery. Appl Environ Microbiol. 2007;73(20):6436-6443.doi:10.1128/AEM.00586-0717720843
28.
MascherT.ZimmerSL.SmithT-A.HelmannJD. Antibiotic-inducible promoter regulated by the cell envelope stress-sensing two-component system LiaRS of Bacillus subtilis. Antimicrob Agents Chemother. 2004;48(8):2888-2896.doi:10.1128/AAC.48.8.2888-2896.200415273097
29.
VesterlundS.PalttaJ.LaukováA.KarpM.OuwehandAC. Rapid screening method for the detection of antimicrobial substances. J Microbiol Methods. 2004;57(1):23-31.doi:10.1016/j.mimet.2003.11.01415003685
30.
MurphyCKinventor; High throughput screen for inhibitors of the folate biosynthetic pathway in bacteria patent application US20020164602A1. 2002.
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.