Structural diversity of triterpene glycosides produced by sea cucumbers or holothurians (Holothuroidea, Echinodermata) is extremely high, although all of them are either lanostane derivatives or, rarely, products of their molecular rearrangements. The majority of them are holostane derivatives possessing an 18(20)-lanostane lactone as aglycone. They contain carbohydrate chains consisting of one to six monosaccharide units including sulfated ones. The glycosides demonstrate interesting biological activities, mainly caused by membranolytic action, namely cytotoxic, ichthyotoxic, antifungal, and hemolytic properties, as well as a series of additional effects at sub-toxic doses, including immunomodulatory, and cancer preventive. This review summarizes the literature data concerning structures and biological activities of all the new triterpene glycosides isolated from sea cucumbers during 2017 to 2021.
Sea cucumbers are animals belonging to the class Holothuroidea of the phylum Echinodermata. They have lost a skeleton, which was reduced to separate ossicles during evolution. Some sea cucumbers have a secondary skeleton formed by merged ossicles. The main defensive chemical tools protecting these animals against such predators as fish are toxic triterpene glycosides of either the lanostane series or, rarely, products of their intramolecular rearrangements. Most of them preferably have an 18(20)-lactone and carbohydrate chains, mainly consisting of two to six sugars, some of which can be sulfated. The sugars, including xylose, quinovose, glucose, 3-O-methylglucose, 3-O-methylxylose, 3-O-methylglucuronic acid, 3-O-methylquinovose and 6-O-acetylglucose, belong to the D-series and are in the β-pyranose form. Because of the independent presence or absence of double bonds, hydroxy groups, acetates, keto- and other functionalities in their aglycones and numerous variants of sulfated or non-sulfated carbohydrate chains, sea cucumber glycosides are a unique example of great biochemical diversity. As result, these natural products have attracted the attention of several generations of chemists and biologists.1‐7
It is of interest that these glycosides demonstrate interesting biological activities, caused by membranolytic action, namely cytotoxic, ichthyotoxic, antifungal, and hemolytic effects, preferably having a sterol-dependant character.8
The activity of sub-toxic doses of the glycosides have attracted special attention and includes immunomodulatory, cancer preventive and other properties. Generally, glycosides and preparations thereof are considered as perspective leads with cytotoxic, antitumor, cancer preventive and immunomodulatory properties.9‐11 The possible practical use of these compounds has stimulated serious efforts of scientists directed to studies of the molecular-genetic basis of sea cucumber glycosides biosynthesis, namely to the genes of 2,3-oxydosqualene cyclases, cytochrome C, glycosidases, and glycosyltransferases.12,13 The main goal of these investigations consists in the biotechnological application of these enzymes in the obtaining of glycosides and their corresponding derivatives. Interesting studies concerning the total chemical synthesis of sea cucumber glycosides are also known.14,15
Recently, significant progress in chromatographic methods of separation and physicochemical methods of structural analysis of natural products has been achieved, mainly through 2D-NMR spectroscopic procedures and high resolution mass-spectrometry. As result, many dozens of new structures of sea cucumber triterpene glycosides have been discovered, which remain outside of earlier reviews. In this article we intend to review all the works concerning the structural elucidation of new sea cucumber glycosides carried out from 2017 to 2021 and briefly discuss the data concerning their biological activities studied in the same period. Herein the review covers only compounds isolated as individual compounds and studied by NMR spectroscopy and does not include the numerous structures deduced by HPLC-MS because the use of mass spectrometry does not allow complete structures to be set.
Results and Discussion
Structures of new Triterpene Glycosides From sea Cucumbers Belonging to the Order Dendrochirotida and Their Biological Activities
Two unprecedented triterpene glycosides, fallaxosides B1 (1) and D3 (2) (Figure 1), with two new skeletal types of aglycone, have been found in the sea cucumber Cucumaria fallax (Cucumariidae, Dendrochirotida).16 It seems to be obvious that the first aglycone is formed as result of intramolecular aldol condensation of a precursor containing a 1,6-diketone fragment (Figure 2). The second aglycone could be formed by pinacol-pinacolone-like rearrangement of another precursor, probably containing a 7,8,9-trihydroxy fragment (or by relative Meinwald rearrangement of its 8,9-epoxy-7-hydroxy analog, an acid-catalyzed conversion of a tetrasubstituted epoxide to a carbonyl compound) (Figure 3). The pentasaccharide chains of 1 and 2 branched by the second monosaccharide residue are common in the glycosides of sea cucumbers belonging to the genus Cucumaria. Both of the carbohydrate chains are sulfated at C-4 of the first monosaccharide residue (xylose); the chain of 1 is additionally sulfated at C-6 of the terminal 3-O-methylglucose residue and is disulfated. The carbohydrate chain of 2 bears two additional sulfate groups at C-6 of the terminal 3-O-methylglucose and C-6 of the third monosaccaride residue (glucose). The structures of these compounds were established by analysis of 1D and 2D NMR spectral, ESI MS, and CD data using quantum-chemical calculations.
Chemical structure of fallaxosides B1 (1) and D3 (2).
Possible biosynthetic pathway to fallaxoside B1 (2) aglycone.
Possible biosynthetic pathway for the formation of fallaxoside D3 aglycone.
Thirteen new mono-, di- and trisulfated triterpene glycosides, quadrangularisosides A (3), A1 (4), B (5); B1 (6) B2 (7), C (8), C1 (9), D (10), D1-D4 (11-14) and E (15) have been isolated from the sea cucumber Colochirus quadrangularis (Cucumariidae, Dendrochirotida), collected in Vietnamese shallow waters (Figure 4).17 Their structures were elucidated by 2D NMR spectroscopy and HR-ESI-MS (high-resolution electrospray-ionization mass spectrometry). The novel carbohydrate moieties of quadrangularisosides, belonging to group D (10 − 14), and quadrangularisoside E (15) contain three sulfate groups, with one of the sulfates in an unusual position, at C-4 of the terminal 3-O-methylglucose. Quadrangularisosides A (3) and D3 (13), as well as quadrangularisosides A1 (4) and D4 (14), contain new aglycones having 25-hydroperoxy or 24-hydroperoxy groups in the side chains, respectively. Recently, coloquadranoside A (16), a tetraoside sulfated at C-4 of the first xylose residue and having two xylose residues in the carbohydrate chain, was isolated from the same sea cucumber species by Chinese investigators. The glycoside has a trivial holostane aglycone with 7(8)- and 25(26)-double bonds and a 16β-acetoxy group.18 Еarlier, the structure of philinopside B was published by Chinese authors as a disulfated glycoside having a structure almost identical to that of 16, but with an additional sulfate group attached to С-2 of the third monosaccharide residue (xylose)19. In the Chinese paper, thorough analysis of the NMR data showed the absence of the second sulfate group in the carbohydrate chain of philinopside B. In the 13С NMR spectra of philinoposide B and its desulfated derivative only the signals corresponding to the first xylose residue differed, indicating the presence of one sulfate group at С-4 of the first xylose. Moreover, in the Chinese paper, the HR mass-spectrometric data were absent and the peaks of fragment ions, observed as a result of the molecular ion fragmentation, did not correspond to the disulfated compound. It seems obvious that the structure of philinopside B needs to be reconsidered and actually it may coincide with the structure of coloquadranoside A (16).
The cytotoxic effects of glycosides 3 to 15 against normal epithelial JB-6 cells, erythrocytes, human colorectal adenocarcinoma HT-29 and mouse neuroblastoma Neuro 2a cells were investigated. The studied substances were shown to be strong hemolytics. The structural features most significantly affecting bioactivity were found to be the hydroperoxy-group in the side chains and the number of sulfate groups. The activities of monosulfated quadrangularisosides of group A (3, 4) against Neuro 2a and JB-6 cells, and erythrocytes were relatively low due to the presence of the hydroperoxy group, whereas trisulfated quadrangularisosides D3 (13) and D4 (14), with the same aglycones as in 3 and 4, were the least active substances among the tested compounds due to the combination of their structural peculiarities. Erythrocytes were more sensitive to the glycosides than either Neuro 2a or JB-6 cells, but structure-activity relationships for the glycosides 3 to 15 were similar in all the cell lines used. The glycosides 5–7, 10 and 11 effectively decreased the viability of HT-29 cells. Quadrangularisosides A1 (4), C (8), C1 (9) and E (15) possessed strong inhibitory action on colony formation of HT-29 cells at concentrations much lower than their IC50. The synergic action of these glycosides, applied at a dose of 0.02 μM, and radioactive irradiation (1 Gy) decreased the number of tumor cell colonies. Glycosides 3, 6 and 11 enhanced the effect of irradiation by about 30%. Coloquadranoside A (16) showed strong antifungal action, and increased the apoptosis and decreased the migration of HCT-116 tumor cells. Glycoside 16 also demonstrated anti-angiogenetic action, decreasing cell migration, cell proliferation and tube formation in the human intestinal microvascular endothelial cells (HIMEC) model. Homograft and xenograft tumor models were used for testing the activity of glycoside 16in vivo. It was active mainly through reducing the weight and volume of tumors. This glycoside also demonstrated immunomodulatory activity in immunodeficient mice used in antitumor experiments. The authors concluded that the presence of a 25(26)-double bond and 16β-OAc seemed to be useful for the membranolytic action of sea cucumber glycosides.
Six new monosulfated triterpene tetra-, penta- and hexaosides, namely, kurilosides A1 (17), A2 (18), C1 (19), D (20), E (21) and F (22), along with the known kuriloside A, having unusual non-holostane aglycones without a lactone, have been isolated from the sea cucumber Thyonidium (=Duasmodactyla) kurilensis (Levin) (Cucumariidae, Dendrochirotida) (Figure 5), collected in the Sea of Okhotsk near Onekotan Island. Their structures were elucidated by 2D NMR spectroscopy and HR-ESI-MS. Kurilosides, belonging to groups A and E, contain carbohydrate chains of rare architecture (a pentasaccharide branched by C-4 Xyl1), and differ from each other in the second sugar (quinovose or glucose, correspondingly). Kurilosides of group C are characterized by a unique tetrasaccharide chain branched at C-4 of Xyl1, while kurilosides belonging to groups D and F are hexaosides differing by the presence of an O-methyl group in the fourth (terminal) sugar unit. All these glycosides contain a sulfate group at C-6 of the glucose residue attached to C-4 of Xyl1, and non-holostane aglycones having a 9(11)-double bond but lacking the γ-lactone.20
Chemical structures of glycosides isolated from Thyonidium kurilensis:17 – kuriloside A1; 18 – kuriloside A2; 19 –kuriloside C1; 20 – kuriloside D; 21 – kuriloside E; 22 – kuriloside F.
The cytotoxic activities of glycosides 17 to 22 against mouse neuroblastoma Neuro 2a cells, normal epithelial JB-6 cells and erythrocytes were studied. The activities were weak due to the absence of 18(20)-lactones and shortened side chains in the aglycones. However, kuriloside A1 (17) was surprisingly the most active compound in this series, demonstrating strong cytotoxicity against erythrocytes and JB-6 cells and a moderate effect against Neuro 2a cells. All the other glycosides were inactive against Neuro 2a cells, but revealed weak action on JB-6 cells.
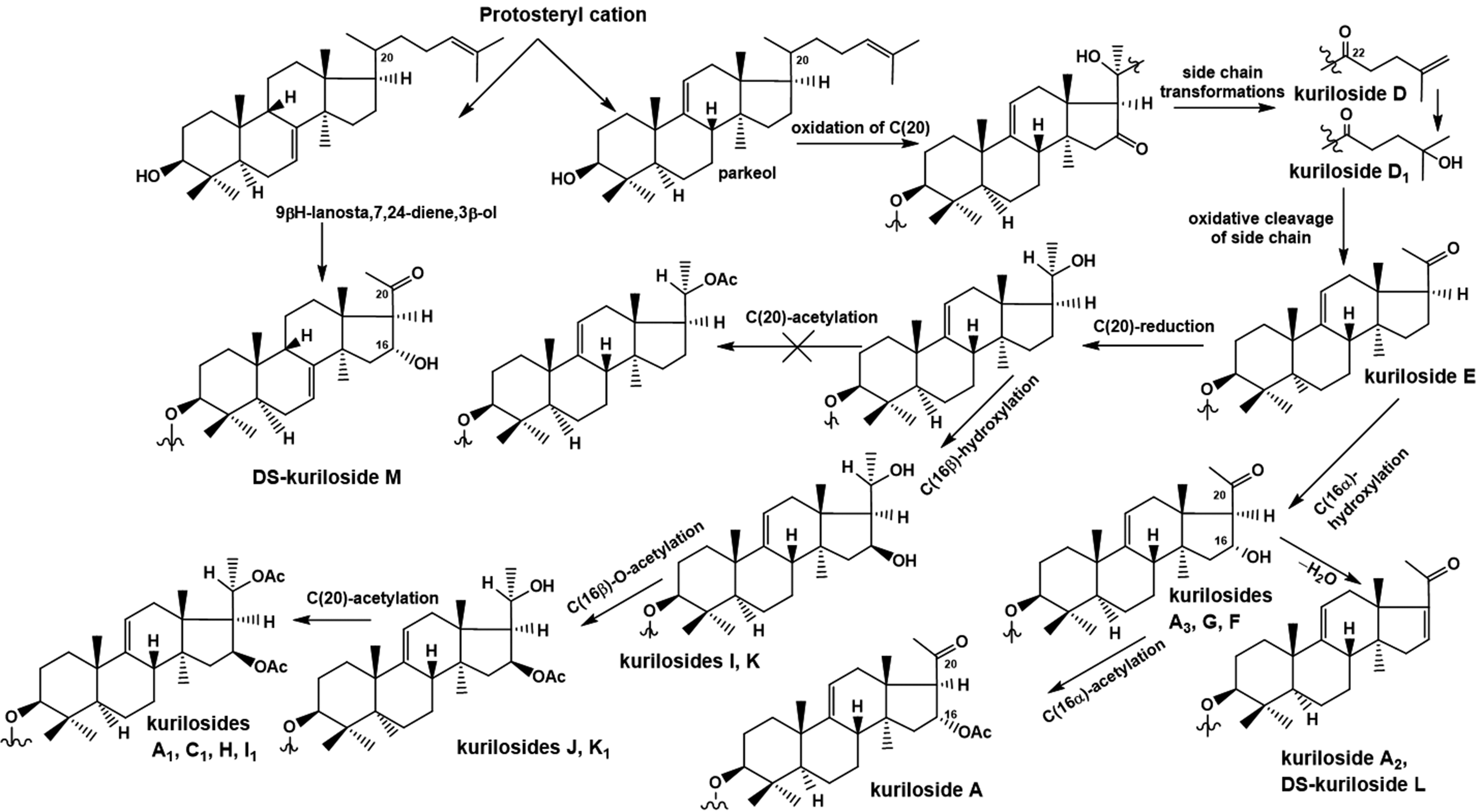
In the completion of studies on triterpene glycosides from Thyonidium kurilensis, nine new mono-, di-, and trisulfated triterpene penta- and hexaosides, kurilosides A3 (23), D1 (24), G (25), H (26), I (27), I1 (28), J (29), K (30), and K1 (31) (Figure 6) and two desulfated derivatives, DS-kuriloside L (33), having a trisaccharide branched chain, and DS-kuriloside M (32) (Figure 7), having a hexa-nor-lanostane aglycone with a 7(8)-double bond, were isolated.21 Structures of native glycosides and their desulfated derivatives were elucidated using 2D NMR spectroscopy and high resolution ESI mass-spectrometry. Five earlier unknown carbohydrate chains and two aglycones (having a 16β,20S-dihydroxy-fragment and a 16β-acetoxy,20S-hydroxy fragment) were found in these glycosides. All the glycosides 23 to 31 have a sulfate group at C-6 of Glc, attached to C-4 of Xyl1, while the positions of the other sulfate groups vary in different groups of kurilosides. Analysis of the structural features of the aglycones and the carbohydrate chains of all the glycosides of T. kurilensis showed their biogenetic relationships (Figure 8).
Chemical structures of desulfated glycosides obtained from Thyonidium kurilensis:32 – DS-kuriloside M; 33 – DS-kuriloside L.
Biosynthetic pathways to aglycones of glycosides from T. kurilensis.
Nine new triterpene glycosides were isolated from the sea cucumber Thyonidium (=Duasmodactyla) kurilensis in addition to the series of kurilosides, found recently.20,21 Five new types of carbohydrate chains (kurilosides of groups G–K) were discovered. There were trisulfated penta- (kurilosides of the group I [27, 28]) and hexaosides {kuriloside H (26)} among them. Kuriloside H (26) is the second example of the most polar triterpene glycoside, along with tetrasulfated pentaosides found earlier in the sea cucumber Psolus fabricii (see below). The structures of disulfated hexa- and pentasaccharide chains of kurilosides of groups G (25), J (29), and K (30, 31) reveal a combinatorial (mosaic) type of biosynthesis of the glycosides, namely, different positions of the sulfate.22 Nevertheless, the position of the sulfate group at C-6 Glc, attached to C-4 Xyl1 remained the same in all the glycosides from T. kurilensis. Three new non-holostane aglycones lacking a lactone ring, two of them being of the 22,23,24,25,26,27-hexa-nor-lanostane type and one having a normal side chain, were found in glycosides 23 to 31. The majority of the aglycones in T. kurilensis glycosides differed from each other in the substituents at C-16 (α- and β-oriented hydroxy- or acetoxy groups, or keto-group) and at C-20 (hydroxy-, acetoxy-, or keto-groups), representing biogenetically related rows of compounds. The glycosides with 16α-substituents were isolated from T. kurilensis only.20 The 16β-hydroxylated aglycones were also found for the first time. Such compounds are undoubtedly the “hot metabolites”, biosynthetic intermediates or precursors of the aglycones with a 16β-acetoxy-group.
Cytotoxic activities of compounds 23 to 31 on mouse neuroblastoma Neuro 2a, normal epithelial JB-6 cells, and erythrocytes were also studied. The highest cytotoxicity in the series was demonstrated by the trisulfated hexaoside kuriloside H (26), having acetoxy-groups at C-16 and C-20, that obviously compensated for the absence of a side chain, essential for the membranolytic action of the glycosides. Kuriloside I1 (28), differing from 26 in lacking a terminal glucose residue in the bottom semi-chain, was slightly less active. Compounds 23 to 25, 27, and 30 did not demonstrate cytotoxic activity due to the presence of hydroxy groups in their aglycones.
Six monosulfated glycosides, cladolosides I1 (34), I2 (35), J1 (36), K1 (37), K2 (38) and L1 (39) (Figure 9) have been isolated from the tropical Indo-West Pacific sea cucumber Cladolabes schmeltzii (Cladolabinae, Sclerodactylidae, Dendrochirotida).23 Their structures were elucidated by 2D NMR spectroscopy and high resolution mass-spectrometry. Cladolosides of groups I and J contain pentasaccharide carbohydrate chains sulfated at the terminal 3-O-methyl-glucose residue and branched by C-4 of the first xylose residue, but differing from each other in the length of carbohydrate half-chains. Cladolosides of groups K and L contain hexasaccharide sugar chains with different positions of the sulfated terminal 3-O-methyl-glucose residue (the fourth or the sixth monosaccharide residue). Sulfated hexasaccharide carbohydrate chains were first found in sea cucumber glycosides. The pentasaccharide carbohydrate chain of cladoloside J1 (35), having a disaccharide moiety, consisted of glucose and sulfated 3-O-methyl-glucose, attached to the first xylose in the trisaccharide linear fragment, which is also very unusual.
The cytotoxic activities of glycosides 34 to 39 were studied against mouse spleenocytes and ascite forms of mouse Ehrlich carcinoma cells, as well as hemolytic activity against mouse erythrocytes. All these compounds demonstrated rather strong cytotoxic and hemolytic effects, except cladoloside K2 (38) that revealed only moderate cytotoxic and hemolytic activities due to the presence of a hydroxy group in the aglycone side chain. The absence of a linear tetrasaccharide fragment, important for membranolytic activities in cladoloside J1 (36), decreased the cytotoxicity against spleenocytes and to a lesser degree against mouse Ehrlich carcinoma cells, as compared to the other glycosides.
In continuation of these studies, eight new triterpene oligoglycosides, cladolosides C4 (40), D1 (41), D2 (42), M (43), M1 (44), M2 (45), N (46) and Q (47) (Figure 10), were isolated from the same species.24 Structures of these glycosides were elucidated by 2D NMR spectroscopy and HR ESI mass spectrometry. A novel hexasaccharide carbohydrate chain having xylose residues as the first, second and third sugars was found in glycoside 46. Cladoloside C4 (40) contains a very uncommon 20,21,22,23,24,25,26,27-octa-norlanostane aglycone. Cladolosides D1 (41), D2 (42) and Q (47) are new representatives of the hexaosides with a non-methylated terminal sugar unit in the “upper” half-chain.
Analysis of the aglycone structures of the glycosides of C. schmeltzii revealed their biogenetic interconnections. The majority of cladolosides have holostane-type aglycones (with 18[20]-lactone), differing in the substituents at C-16 and C-22, and with a terminal double bond in the side chain. It is known that holostane aglycones are biosynthesized via hydroxylation at C-20, followed by C-18 oxidation that results in the formation of an 18(20)-lactone. With the simultaneous presence of hydroxy groups at C-16 and C-20 in the 18-carboxylated derivative, 18(16)-lactone formation occurs. Either acetylation at С-16 or oxidation of the corresponding hydroxy group to a keto-group apparently precedes the carboxylation at C-18, which prevents the formation of an 18(16)-lactone. The incorporation of a functionality at C-16 could also be realized after 18(20)-lactonization, but the absence of unsubstitution at C-16 in the triterpene glycosides of C. schmeltzii corroborated the precedence of C-16 oxidation to that of C-18. Hence, the holostane aglycones of known cladolosides (more than 30 compounds) could be biosynthesized from one generic precursor by two alternative ways due to the time shifting of the oxidation at C-18 and C-22 (Figure 11). Moreover, modifications of the side chain could occur before the C-18 oxidation.
Possible biosynthetic pathways of glycoside aglycones in Cladolabes schmeltzii.
There is another biosynthetic pathway leading to the formation of a unique aglycone of cladoloside С4 (40). It is realized through the oxidation of C-22 (before C-18 oxidation), followed by side chain cleavage and oxidative cleavage of the 17(20)-bond with consequent dehydration. Similar oxidative transformations take place in the biosynthesis of steroidal hormones of vertebrates. The presence of hydroxy- or acetoxy-groups at C-22 in the majority of the aglycones of cladolosides confirms the possibility of the realization of such a biosynthetic pathway. All these aglycone modifications corroborate the mosaic type of biosynthesis of sea cucumber triterpene glycosides, ie it is organized as a metabolic network.
Cytotoxic activities were evaluated of glycosides 40 to 47 against the ascite form of mouse Ehrlich carcinoma cells, mouse erythrocytes and human colorectal adenocarcinoma HT-29 cells. The majority of the glycosides revealed potent cytotoxic and hemolytic activities, except cladoloside D2 (42), which demonstrating a moderate effect. The decreased activity of 42 can be caused by the presence of an hydroxy group in the aglycone side chain. Cladoloside C4 (40) was surprisingly highly cytotoxic, despite the absence of a lactone and the side chain in the aglycone. The erythrocytes were more sensitive to the action of compounds 41 to 47 in comparison with the Ehrlich carcinoma cells, but glycoside 40 demonstrated the opposite tendency. The influence of compounds 40 to 47 on cell viability, formation and growth of colonies of human colorectal adenocarcinoma HT-29 cells has been tested. Cladoloside D2 (42) was not cytotoxic against HT-29 cells at a dose of 20 μM; this correlated with the data on Ehrlich carcinoma cells. In contrast, with experiments on Ehrlich carcinoma cells, cladoloside C4 (40), having an unusual octanorlanostane aglycone, was not cytotoxic up to a concentration of 20 μM. The action of compounds 40 to 47 on colony formation of HT-29 cells was studied at non-toxic concentrations by soft agar assay. Cladoloside D2 (42) did not inhibit colony formation and growth of HT-29 cells by 50% up to a concentration of 10 μM. Cladoloside C4 (40) was less active in this test in comparison with 41 and 43 to 47. The highest inhibition of colony formation and growth of HT-29 cells was demonstrated by cladoloside M1 (44) (ICCF50 0.05 μM), whereas it was active against the cancer cells at 16.9 μM (IC50).
Moreover, synergism was observed in the decrease in the number of colonies of HT-29 cells between radioactive irradiation and non-toxic doses of compounds 40 to 47. Cladoloside N (46) was the most active and increased the inhibitory effect from radiation by 75%.
In the completion of investigations of glycosides from the sea cucumber Cladolabes schmeltzii (Sclerodactylidae, Dendrochirotida) six new triterpene oligoglycosides, cladolosides O (48), P (49), P1 (50), P2 (51), P3 (52) and R (53) (Figure 12), have been isolated.25 Their structures were elucidated using 2D NMR spectroscopy and HR ESI mass spectrometry. Cladoloside O (48) is featured by a pentasaccharide carbohydrate chain. Cladolosides of group P and cladoloside R (53) include novel hexasaccharide carbohydrate chains with different positions of non-methylated terminal monosaccharide residues.
All the isolated glycosides revealed strong cytotoxic activities against mouse Ehrlich carcinoma cells (ascite form) and mouse erythrocytes. The cytotoxicity against human colorectal adenocarcinoma HT-29 cells was lower. Glycosides 48 to 53 inhibited the colony formation and growth of HT-29 cells at non-cytotoxic concentrations. Cladoloside P1 (50) had the highest inhibitory effect. The synergism between radioactive irradiation and non-toxic doses of glycosides 48 to 53 on colony formation of HT-29 tumor cells was also discovered. Cladolosides P2 (51) and P3 (52) were the most active. They increased the inhibitory effect of radiation by more than 70%.
In total, 40 novel triterpene glycosides were isolated from C. schmeltzii. These compounds contained 19 types of carbohydrate chains and 9 different aglycones. Their structure metabolic network showed that each type of carbohydrate chain could be biosynthesized as a result of attachment of different monosaccharides to certain positions of the forming sugar chains. The introduction of sulfate groups seems to be a final stage of the carbohydrate chain biosynthesis in C. schmeltzii because the sulfate groups are attached to the terminal 3-O-methylated monosaccharide residues. The structural diversity of the glycosides of C. schmeltzii could be explained by the process of active evolutionary searching followed by selection of the compounds that more effectively performed their biological functions. On the other hand, chemical diversity contributes to the quick adaptation of organism-producer to the changing environmental conditions thereby providing evolutionary plasticity. The ability of different variable structural elements of the glycosides providing their corresponding contributions to the general biological activity of a molecule and its biological functions (structural degeneracy of biological function) may also explain such impressive biodiversity.26
Recently, the metabolic profile of triterpene glycosides of Eupentacta fraudatrix (Sclerodactylidae, Dendrochirotida) has been studied using LC-ESI MS. In total, 54 compounds were found, including 26 sulfated, 18 non-sulfated and 10 disulfated glycosides. One of the most significant findings was the presence of two pairs of pentaosides (sulfated and non-sulfated) having 3-O-methylglucose as a terminal monosaccharide residue. Such a structural peculiarity has never been found before in the glycosides of E. fraudatrix, having a 3-O-methylxylose residue as a terminal unit.27 It would be very interesting to isolate any of these “candidates” having 3-O-methylglucose in amounts sufficient for obtaining their NMR spectra, including 2D NMR procedures in order to confirm the presence of 3-O-methylglucose. As result of these special efforts, a new minor triterpene glycoside, namely cucumarioside D (54) (Figure 13), has been isolated from E. fraudatrix as an individual substance.28 The structure of the glycoside, elucidated by 2D NMR spectroscopy and high resolution mass-spectrometry, is characterized by the non-sulfated pentasaccharide carbohydrate chain branched by the second monosaccharide residue (quinovose) with 3-O-methylglucose as a terminal sugar.
Structure of glycoside: 54 – cucumarioside D.
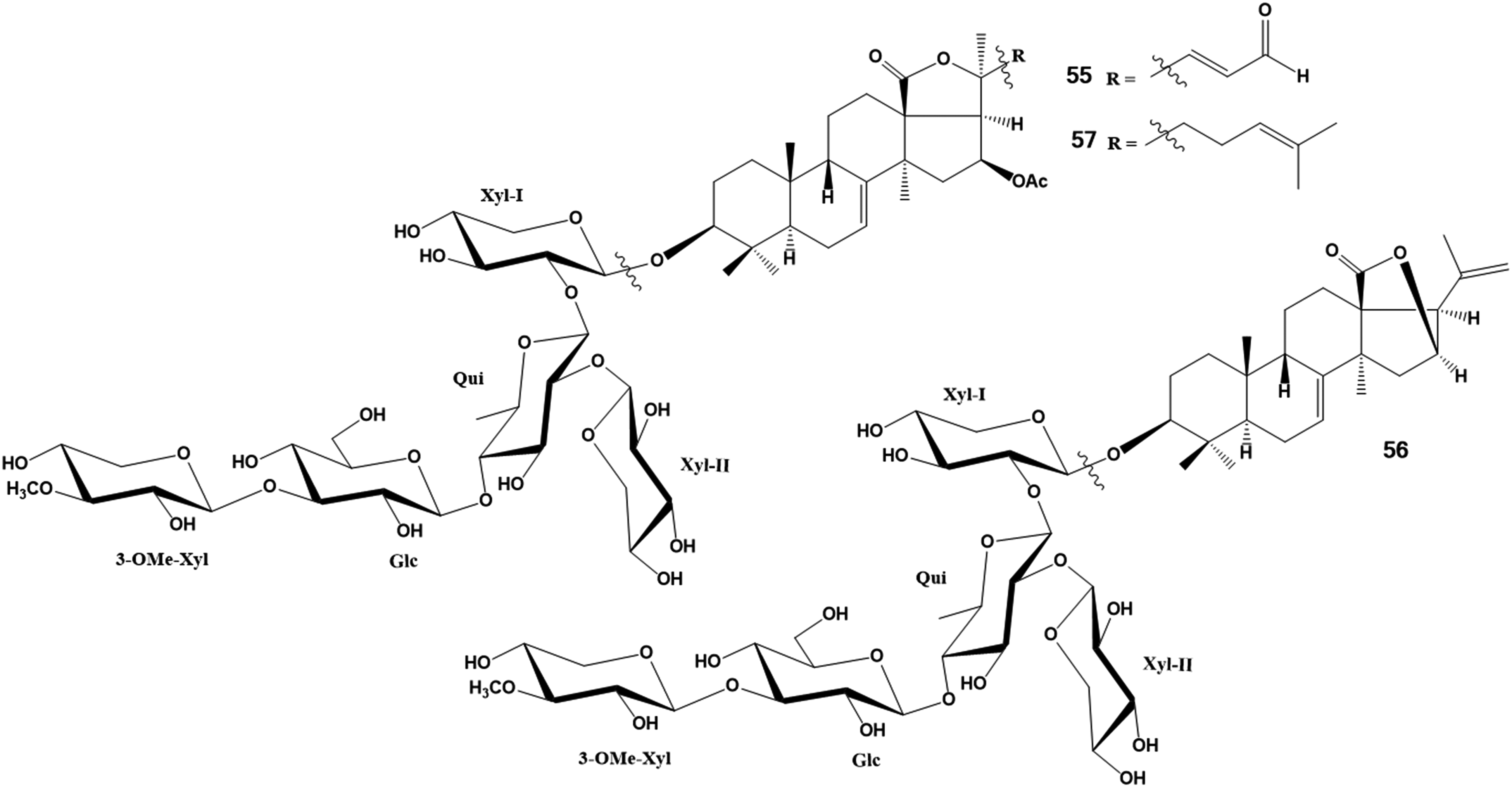
Three new triterpene glycosides, pacificusosides A − C (55 − 57), structurally very closely related to the glycosides from E. fraufatrix, and three previously known triterpene glycosides from the same species of sea cucumber, cucumariosides C1, C2, and A10, were isolated from the alcoholic extract of the Far Eastern starfish Solaster pacificus. The structures of 55 − 57 were elucidated by extensive NMR and ESIMS experiments, and chemical transformations used for determination of the absolute configurations of the sugars (Figure 14). Pacificusoside A (55) has a unique shortened side chain in the aglycone with a terminal aldehyde group at C-24. All the known glycosides and the new ones were undoubtedly obtained by the starfish during feeding on the sea cucumber, possibly E. fraudatrix, as described in the literature.29 The cytotoxicity of compounds 55 to 57 was studied in relation to human embryonic kidney HEK 293 cells, melanoma RPMI-7951 cells, colorectal carcinoma HT-29 cells, and breast cancer MDA-MB-231 cells using MTS assay. The cytotoxic effects of 55 to 57 were decreased by cholesterol. Compound 57 almost completely suppressed colony formation of HT-29, RPMI-7951, and MDA-MB-231 cells on agar media at a non-toxic concentration of 0.5 µM.
Structures of glycosides: 55 – pacificusoside A; 56 – pacificusoside B; 57 – pacificusoside C.
Three new triterpene glycosides, psolusosides C1 (58), C2 (59) and D1 (60) (Figure 15), have been isolated from the sea cucumber Psolus fabricii collected from the Sea of Okhotsk in the shallow waters of Onekotan Island (Kurile Islands) (Figure 15). Five known glycosides have also been found: cladolosides B, M1, P2, isolated earlier from sea cucumbers of the genus Cladolabes, and holotoxin A1, as well as 27-nor-25-oxo-holotoxin A1, found earlier in Apostichopus japonicus. The structure elucidation of the new substances and identification of known compounds were made by 2D NMR spectroscopy and HR mass-spectrometry. Compounds 58 to 60 possess hexasaccharide carbohydrate chains, identical for 58 and 59, but differing in the third monosaccharide residue from 60. All these glycosides have holostane type aglycones with a 9(11)-double bond and 16-keto-group and differ from each other in details of side chain structures. The aglycone of psolusoside D1 (60) and carbohydrate chains of all the new compounds 58 to 60 were found earlier in the glycosides of C. schmeltzii. All these findings are an example of evolutionary parallelisms in the taxonomical distribution of triterpene glycosides.30
Structures of glycosides from Psolus fabricii: 58 – psolusoside C1; 59 – psolusoside C2; 60 – psolusoside D1.
Continuation of investigations of Psolus fabricii led to the isolation of five new triterpene glycosides, psolusosides C3 (61), D2 (62), D3 (63) D4 (64) and D5 (65) (Figure 16). The structures of these glycosides were elucidated by 2D NMR spectroscopy and HR mass-spectrometry. All the compounds contain hexasaccharide carbohydrate chains, differing from each other in the third monosaccharide residue: xylose was found in psolusosides of group C and glucose – in group D. Aglycones of the isolated compounds belong to the holostane type, containing a 9(11)-double bond and a 16-keto-group, but with different side chains. The aglycone of psolusoside D3 (63) possesses a very rare hydroperoxy group at C-25.31
Structures of glycosides from Psolus fabricii: Structures of glycosides: 61 – psolusoside C3; 62 – psolusoside D2; 63 – psolusoside D3; 64 – psolusoside D4; 65 – psolusoside D5.
The series of side chains characteristic of psolusosides C1 (58) and D4 (64) (24-hydroxy-group and 25[26]-double bond), C2 (59) and D2 (61) (24-keto-group and 25[26]-double bond), and C3 (61) and D5 (65) (25-hydroxy-group and 23E-double bond) are simultaneously presented in the glycosides from different sea cucumber taxa. The same set of side chains has been found in aglycones of glycosides from Achlionice violaescupidata (Elpidiidae, Elasipodida), Eupentacta fraudatrix (Sclerodactylidae, Dendrochirotida) and Colochirus robustus (Cucumariidae, Dendrochirotida).1 These data revealed common biosynthetic patterns realized in sea cucumbers belonging to different families and orders.
The cytotoxic action of glycosides 61 to 65 along with psolusosides C1 (58), C2 (59), D1 (60), and 27-nor-25-oxo-holotoxin A1 on mouse erythrocytes (hemolytic activity), the ascite form of mouse Ehrlich carcinoma cells, and neuroblastoma Neuro 2a cells have been studied. These substances have hexasaccharide carbohydrate chains and holostane-type aglycones that should provide high membranolytic activity. Actually, five of the compounds were highly cytotoxic. 27-nor-25-oxo-holotoxin A1 having a shortened side chain demonstrated moderate cytotoxicity against erythrocytes and Neuro 2a cells, but was not active against Ehrlich carcinoma cells. Psolusosides C1 (58) and C3 (61) were the least active. Nevertheless, they revealed moderate hemolytic and cytotoxic action. Such a decrease in activities could be explained by the presence of an hydroxy group in the side chains of the aglycones. However, the sugar chain having a linear tetrasaccharide fragment partially compensates for the activity-decreasing influence of the hydroxy-group in the side chain. Psolusosides C2 (59), D1 (60), and D2 (62), having aglycones without such a functionality, showed the highest hemolytic action. Psolusoside D3 (63) containing a peroxide group in the side chain of the aglycone was surprisingly highly cytotoxic. The aglycones of psolusosides C1 (58) and C3 (61) were identical to those of psolusosides D4 (64) and D5 (65), correspondingly. Compounds 64 and 65 differ from the previous glycosides by the third monosaccharide unit in the carbohydrate chain (glucose instead of xylose). However, psolusosides of group D (64, 65) were more active than their structural analogs belonging to group C.
In continuation of the studies of the glycosides from Psolus fabricii, seven sulfated triterpene glycosides have been isolated, psolusosides B (66), E (67), F (68), G (69), H (70), H1 (71) and I (72) (Figure 17), along with the earlier known psolusoside A and colochiroside D. Additionally, the structure of psolusoside B (66), elucidated in 1989 as a monosulfated tetraoside,32 was revised by application of modern NMR spectroscopic and particularly MS techniques and proved to be a disulfated tetraoside.33 The structures of the other glycosides were elucidated by 2D NMR spectroscopy and HR-ESI-MS. Psolusosides E (67), F (68) and G (69) contain holostane aglycones identical to each other, but differ in their sugar compositions, namely by the number and positions of sulfate groups in the linear tetrasaccharide carbohydrate chains. Psolusosides H (70) and H1 (71) are characterized by an unusual sulfated trisaccharide carbohydrate moiety with glucose as the second monosaccharide unit. Psolusoside I (72) has an unprecedented branched tetrasaccharide disulfated carbohydrate chain with the xylose unit in the second position.
Chemical structures of glycosides isolated from Psolus fabricii:66 – psolusoside B; 67 – psolusoside E; 68 – psolusoside F; 69 – psolusoside G; 70 – psolusoside H; 71 – psolusoside H1; 72 – psolusoside I.
The cytotoxic activities of glycosides 67 to 72 against several mouse cell lines (ascite form of Ehrlich carcinoma, neuroblastoma Neuro 2a, normal epithelial JB-6 cells, and erythrocytes) were quite different, because the hemolytic effects of the tested substances were higher than their cytotoxicity, especially against the ascite form of Ehrlich carcinoma. It is interesting that psolusoside G (69) was not cytotoxic against normal JB-6 cells, but was highly active against Neuro 2a cells. The cytotoxic activity against human colorectal adenocarcinoma HT-29 cells and the influence on colony formation and growth of HT-29 cells were also studied for glycosides 66 to 69, 70 to 72 and psolusoside A. The highest inhibitory activities were revealed for psolusosides E (67) and F (68).
The synergistic effects of these compounds (0.05 µM) and radioactive irradiation (2 Gy) in relation to HT-29 cells have also been studied. The number of colonies of HT-29 cells was found to be decreased after radiation exposure at a dose of 2 Gy, but a synergic effect of these glycosides and irradiation was not observed.
In a continuation of the studies on the glycosides from P. fabricii, ten new di-, tri- and tetrasulfated triterpene glycosides, namely, psolusosides B1 (73), B2 (74), J (75), K (76), L (77), M (78), N (79), O (80), P (81) and Q (82), were isolated (Figure 18). Structures of these glycosides were established by 2D NMR spectroscopy and HR-ESI-MS.34 It is particularly interesting that highly polar compounds 81 and 82 contain four sulfate groups in their carbohydrate moieties, including two sulfates in the same terminal glucose residue. Glycoside 74 has an unusual non-holostane aglycone with an 18(16)-lactone and a unique 7,8-epoxy fragment.
Psolusosides B1 (73) and B2 (74) have disulfated tetrasaccharide carbohydrate chains branched at C-4 of Xyl1 identical to that of psolusoside B and non-holostane aglycones having an 18(16)-lactone moiety. They differ from other glycosides by a unique structural feature such as a 7,8-epoxy-fragment in 74 or in the combination of unusual features such as a 7-keto-8,9-ene fragment and 18(16)-lactone in 73. Compounds 75 to 82 contain common for sea cucumber aglycones, but unique carbohydrate chains. Psolusosides J (75) and K (76) are characterized by new trisulfated tetrasaccharide branched chains with the terminal glucose unit sulfated at two positions: at C-2 and C-4 in compound 75, and at C-2 and C-6 in compound 76. Psolusosides L (77), M (78) and P (81) have branched at C-4 of Xyl1 pentasaccharide chains with quinovose as the second sugar unit. These compounds differ from each other by the quantity and positions of sulfate groups in the fifth (Glc5) residue. Psolusoside P (81) is tetrasulfated glycoside, containing two sulfate groups at C-4 and C-6 of the same terminal (Glc5) residue. Psolusosides N (79), O (80) and Q (82) have carbohydrate chains with the same architecture as glycosides 77, 78 and 81, but differ from those in the second monosaccharide residue, which is glucose instead of quinovose. Psolusosides N (79) and O (80) are the structural analogs of psolusosides L (77) and M (78), correspondingly, having identical positions of the sulfate groups. Tetrasulfated psolusoside Q (82) differs from psolusoside P (81) by the positions of sulfation – at C-2 and C-6 of the terminal (Glc5) residue. Up to this time, tetrasulfated glycosides had not been found as natural products.
Generally, 27 new and 5 known earlier triterpene glycosides have been isolated from Psolus fabricii. These compounds contain 6 previously unknown aglycones and 13 novel carbohydrate chains.
Sulfated oligosaccharide moieties predominate in the glycosides of P. fabricii. Monosulfated trisaccharide (psolusosides H [70] and H1 [71]) and linear tetrasaccharide (psolusosides E and F) moieties, disulfated branched tetrasaccharide (psolusosides B [66], B1 [73], B2 [74] and I [72]), linear tetrasaccharide (psolusosides A and G [69]) carbohydrate chains, trisulfated branched tetrasaccharide (psolusosides J [75] and K [76]) chains, and finally trisulfated (psolusosides L [77], M [78], N [79], O [80]) and tetrasulfated pentasaccharide (psolusosides P [81] and Q [82]) sugar chains were found in glycosides of P. fabricii. These moieties also differ from each other in the second monosaccharide unit (quinovose, glucose or xylose). The most variable structural feature of the carbohydrate chains of glycosides 73 to 82 is the number (one or two) and positions of sulfate groups in the terminal glucose unit, attached to C-4 of Xyl1. There are three combinations of such positions of sulfate groups in this residue: C-2 and C-4, C-2 and C-6 and C-4 and C-6 of Glc, whereas a single sulfate group is attached to only either C-2 or C-6 of a terminal glucose unit.
Psolusosides (A–Q) are a diverse group of glycosides with the same carbohydrate chain, but different aglycones. All the psolusosides belonging to groups C and D (both containing hexasaccharide non-sulfated sugar chains) contain holostane-type aglycones with a 9(11)-double bond, a 16-keto-group, and different side chains (5 variants). Psolusosides of group B contain exclusively non-holostane aglycones with an 18(16)-lactone and 7(8)-double bond, completely different from the aglycones of the other groups of psolusosides. This could be explained by their special biological functions in the organism-producer. Four holostane aglycones with either a 7(8)- or 9(11)-double bond were found in five glycosides having either trisaccharide (psolusosides H [70] and H1 [71]) or tetrasaccharide branched carbohydrate chains (psolusosides I [72], J [75], K [76]).
Pentaosides (psolusosides L–Q [77-82]) and tetraosides with linear sugar chain fragments (psolusosides A, E [67], F [68], G [69]) contain the same holostane-type aglycone with a 9(11)-double bond. This suggests that linear tetraosides are biosynthetic precursors of pentaosides – psolusosides L (77), M (78) and P (81), which are biosynthesized via glycosylation and sulfation of psolusosides A, E (67) and F (68), correspondingly. Psolusosides N (79), O (80), and Q (82) are formed from psolusoside G (69) through the same processes. Hence, biogenetic analysis of the structures of glycosides found in P. fabricii show that the biosynthesis of the carbohydrate chains and aglycones possess a mosaic (combinatoric) character, but some sequences of the transformations are regular.
The cytotoxic activities of glycosides 73 to 82 against mouse cell lines, including Ehrlich ascites carcinoma cells, neuroblastoma Neuro 2a, normal epithelial JB-6 cells, and erythrocytes, depend both on the structural peculiarities of the glycosides and the type of cells. Psolusoside L (77), a pentaoside having three sulfate groups at C-6 of two glucose and one 3-O-methylglucose residue, along with a holostane aglycone, was the most active tested substance. The presence of a sulfate group at C-2 of the terminal glucose residue attached to C-4 of the first (xylose) residue significantly decreases activities of the corresponding glycosides. Psolusosides belonging to group B (73, 74 and 66) were inactive due to the presence of non-holostane aglycones and tetrasaccharide branched chains sulfated at C-2 of the fourth monosaccharide residue (glucose).
Nine new sulfated triterpene glycosides, magnumosides A1 (83), A2 (84), A3 (85), A4 (86), B1 (87), B2 (88), C1 (89), C2 (90) and C4 (91) (Figure 19), as well as the known colochiroside B2, have been isolated from the tropical Indo-West Pacific sea cucumber Massinium (=Neothyonidium) magnum (Phyllophoridae, Dendrochirotida) collected in Vietnamese shallow waters.35 The structures of the new glycosides were elucidated by 2D NMR spectroscopy and mass spectrometry. The configuration of the C-24 hydroxy-group in glycosides 84, 88 and 90 was found to be 24S, using Mosher's method. All the isolated new glycosides were characterized by non-holostane type lanostane aglycones having an 18(16)-lactone and 7(8)-double bond and differing from each other in the side chains and carbohydrate moieties. Magnumoside A1 (83) has an unprecedented 20(24)-epoxy-group in the aglycone side chain. Magnumosides of group A (83-91) contain disaccharide monosulfated carbohydrate moieties, those of group B (87, 88) tetrasaccharide monosulfated moieties, and, those of group C (89-91) tetrasaccharide disulfated moieties.
The cytotoxic activities of glycosides 83 to 91 in relation to mouse spleen lymphocytes, the ascites form of mouse Ehrlich carcinoma cells, and human colorectal carcinoma DLD-1 cells, as well as their hemolytic effects were studied. The erythrocytes were more sensitive than spleenocytes and cancer cells. Disulfated glycosides 89 and 90 revealed surprisingly high hemolytic and cytotoxic actions in spite of the presence of hydroxy groups in the aglycone side chains. The data concerning cytotoxic activities against DLD-1 human colorectal adenocarcinoma cells correlated well with those for mouse ascites Ehrlich carcinoma cells. This confirmed the usefulness of the latter model for screening of substances for cytotoxicity to human tumor cells. Compounds 85 and 89 significantly inhibited the colony formation and decreased the size of colonies of DLD-1 cancer cells in agar media at non-cytotoxic concentrations. The data concerning synergy of glycosides 85 and 89 to 91 in sub-cytotoxic doses and sub-toxic doses of radiation showed that magnumoside C4 (91) revealed the highest increase (of 45%) in the inhibitory effect of radiation on cell proliferation.
In further studies on the triterpene glycosides from Massinium (=Neothynidium) magnum, two new monosulfated triterpene glycosides, magnumosides B3 (92) and B4 (93), and one new disulfated triterpene glycoside, magnumoside C3 (94), were isolated (Figure 20). Structures of these glycosides were elucidated using 2D NMR spectroscopy and HR ESI mass-spectrometry.36
Structures of glycosides from Massinium (=Neothyonidium) magnum: 92 – magnumoside B3; 93 – magnumoside B4; 94 – magnumoside C3.
All the isolated glycosides possess non-holostane type aglycones having an 18(16)-lactone and a 7(8)-double bond, and differ from each other by the side chain structures and the number of sulfate groups in the carbohydrate chains. Obviously, in the series of magnumosides B3 (92), B4 (93) and C3 (94), the attachment of the second sulfate group in the carbohydrate chain of 94 is a final stage of biosynthetic transformation.
The cytotoxic activities of glycosides 92 to 94 were tested against mouse spleen lymphocytes, the ascite form of mouse Ehrlich carcinoma cells, and human colorectal carcinoma DLD-1, as well as their hemolytic effects. The sensitivity of the cells to the action of the glycosides decreased in the order: erythrocytes → spleenocytes → Ehrlich carcinoma cells. Magnumoside B4 (93), with a 24(25)-double bond in the aglycone side chain, was more active in all the tests in comparison with magnumosides B3 (92) and C3 (94) with a terminal double bond in the side chain. Disulfated glycoside 94 demonstrated lesser hemolytic and cytotoxic activity than the corresponding monosulfated glycosides 92 and 93. This is in good agreement with the known observation that sulfate groups attached to C-6 of terminal glucose and 3-O-methylglucose residues usually decrease cytotoxic activity.8
Magnumoside B4 (93) significantly suppressed the colony formation and decreased the size of colonies of DLD-1 cancer cells at non-cytotoxic doses. The study on synergy of radiation and compounds 92 to 94 in relation to proliferation of DLD-1 cells demonstrated their almost equal enhancing antiproliferative effect on radioactive irradiation.
Hence, in total, 11 new glycosides with non-holostane aglycones with an 18(16)-lactone and one known holostane glycoside were isolated from Massinium (=Neothyonidium) magnum.
Structures of new Triterpene Glycosides From sea Cucumbers Belonging to the Order Synallactida and Their Biological Activities
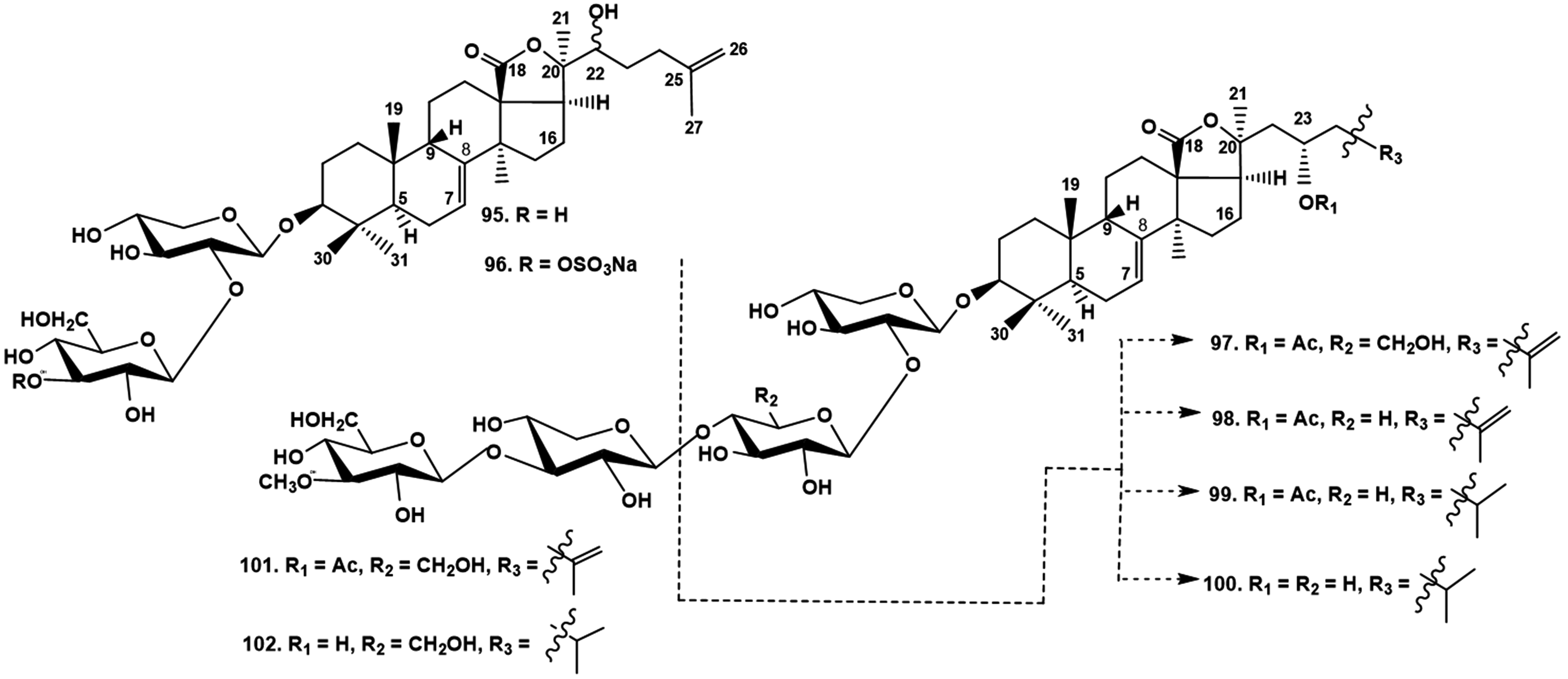
A series of minor triterpene biosides was isolated from the Indo-West Pacific sea cucumber Stichopus horrens (Stichopodidae, Synallactida), including stichorrenosides A–D (95-98), along with known stichoposides A and B, and two progenines, (23S)-23-acetoxyholost-7-en-3β-yl-O-β-D-xylopyranosyl-(1→2)-β-D-xylopyranoside (99) and (23S)-23-hydroxyholost-7-en-3β-yl-O-β-D-xylopyranosyl-(1→2)-β-D-xylopyranoside (100) (Figure 21), obtained earlier by enzymatic hydrolysis of stichoposide E from Stichopus chloronotus.37 These progenins were firstly found as natural products. The authors found a very rare sulfate group at C-3 of the second monosaccharide residue (glucose) in stihorrenoside B (96). All the other isolated biosides were non-sulfated. The same authors isolated a series of non-sulfated tetraosides, including stihorenoside E (101), which is the 25(26)-dehydro-derivative of the known thelenotoside B, and deacetylated thelenotoside B (102), known earlier as a progenin obtained by enzymatic hydrolysis from stichoposide C from S. chloronotus and firstly found as a natural product (Figure 21). All the structures were elucidated using 1D and 2D NMR spectroscopy and HR ESI MS.38
The in vitro cytotoxic activities were studied of stichorrenosides A − D (95 − 98), (23S)-23-acetoxyholost-7-en-3β-yl-O-β-D-xylopyranosyl-(1→2)-β-D-xylopyranoside (99) and (23S)-23-hydroxyholost-7-en-3β-yl-O-β-D-xylopyranosyl-(1→2)-β-D-xylopyranoside (100) against a series of human cancer cell lines, namely Hep-G2 (hepatoma cancer), KB (epidermoid carcinoma), LNCaP (prostate cancer), MCF7 (breast cancer), and SK-Mel2 (melanoma). Compounds 98 and 99 showed strong cytotoxicity against all five tested cancer cell lines, while significant effect was also observed for stichorrenosides C (97) and B (96). The tetraosides, stichorrenoside E (101) and deacetyl thelenotoside B (102) exhibited significant cytotoxic action against the tested cell lines.
Three glycosides were reported to be isolated from the viscera of Apostichopus japonicus (Stichopodidae, Synallactida), namely apostichoposides A–C (103-105) (Figure 22).39 The glycosides have only glucose residues in the carbohydrate chains, except for apostichoposide B (104), which has a xylose residue as the first sugar. The authors could not register the peaks of the molecular ions for glycosides 103 and 105 and explained this by a unique fragmentation with elimination of C-18 and two neighboring oxygen atoms as a CO2 molecule. However, such fragmentation has never been found in numerous mass spectra obtained for several hundred sea cucumber triterpene glycosides. Moreover, the NMR spectral data were not sufficient. The HSQC and HMBC data were presented for the anomeric protons only and were absent for the other carbons of the carbohydrate chains of glycosides 103 to 105. A 1D TOCSY experiment, enabling the deduction of the isolated spin system characteristic for each sugar, was apparently not made. Consequently, these results cannot be taken with sufficient seriousness without additional check.
Proposed structures of the glycosides from the viscera of Apostichopus japonicus. 103 – apostichoposide A; 104 – apostichoposide B; 105 – apostichoposide C.
Two other tetraosides, “compound 1” (106) and “compound 2” (107) with non-holostane aglycones containing an 18(16)-lactone and carbohydrate chains having glucose as the first monosaccharide residue were also isolated from Apostichopus japonicus viscera (Figure 23). The structures of the carbohydrate chains seem to us to be doubtful because of the presence of glucose as the first monosaccharide residue in the isolated glycosides.40 The authors postulated another unique mass spectrometry fragmentation caused by the uncommon aglycone fragmentation resulting in the ion peaks [M – O – H]− and [M + Cl – O]− for glycoside 106, and [M – H]− and [M + Cl]− for 107. Moreover, they reported D-glucose, D-xylose and D-quinovose after acid hydrolysis in both glycosides, although glycoside 106 did not contain a xylose residue. It is not clear why the authors used only used the negative mode of ions registration for obtaining the mass-spectra and did not explain the origin of Cl.
Proposed structures of the glycosides isolated from Apostichopus japonicus viscera. 106 – “compound 1”; 107 – “compound 2”.
Structures of new Triterpene Glycosides From sea Cucumbers Belonging to the Order Holothuriida and Their Biological Activities
Eight triterpene glycosides, including one new one, namely holothurin A5 (108) (Figure 24), were isolated from the Vietnamese sea cucumber Holothuria edulis. Their structures were identified and elucidated using different spectroscopic methods including 1D and 2D NMR spectroscopy and HR-ESI-MS.41 Holothurin A5 (108) has a 23(24)-double bond in the aglycone side chain and a hydroperoxy-group at C-25; this is a rare structural feature in sea cucumber triterpene glycosides. The known glycosides were identified as holothurin A2, marmoroside C, moebioside A, leucospilotaside A, holothurin A, holothurin B and leucospilotaside C.1 Identification of leucospilotaside C, which is a sulfated monoxyloside, is of special interest because it is only the second case of finding a monoside among sea cucumber triterpene glycosides. The new glycoside 108 possessed very slight cytotoxicity against five human cancer cell lines (HepG2, KB, LNCaP, MCF7 and SK-Mel2) due to the presence of peroxide in the aglycone side chain.
Structure of the glycoside isolated from Holothuria edulis. 108 – holothurin A5.
The saponin composition of Holothuria atra collected in the Persian Gulf was studied by a combined approach including LC-MS/MS and molecular networking, followed by the further isolation of pure compounds and their investigation by NMR spectroscopy. The authors showed that the saponin composition of Holothuria atra was more complex than that reported earlier. Four individual glycosides were isolated and characterized by 1D and 2D NMR experiments, including one new glycoside 109, independently also named as holothurin A5 (108) (Figure 25).42 The side chain of 109, unprecedented among triterpene glycosides, is characterized by a 23(24)-ene-22-one fragment and 25-hydroxy group. The other three glycosides were holothurin A, holothurin A2 (echinoside A) and 24-dehydrochinoside A.1 Hence LC-MS/MS, molecular networking and other metabolomic approaches require the application of such HPLC procedures that are necessary for the separation of complicated glycosidic mixtures from sea cucumber extracts and isolation of individual compounds. Glycoside 109 was remarkably cytotoxic against the human cervix carcinoma HeLa cell line.
Structure of new glycoside isolated from Holothuria atra. 109 – holothurin A5.
Several known glycosides belonging to the holothurin B group were isolated from H. atra collected in the Red Sea. The authors postulated that the isolated 12-O-methyl derivative of holothurin B is a natural product, but they used methanol as a solvent during chromatographic separation that suggests the artificial origin of the isolated new compound.43
Conclusions
During the last five years 104 new triterpene glycosides have been isolated from sea cucumbers belonging to the order Dendrochirotida (91 glycosides), a starfish obviously obtained the glycosides by eating the sea cucumber Eupentacta fraudatrix (3 new glycosides), and from Stichopus horrens, a representative of the order Synallactida (8 new glycosides) (Table 1). The structures of four glycosides, isolated from the viscera of Apostichopus japonicus and having a glucose residue as the first sugar, in our opinion, requires checking. Two different glycosides with the same name holothurin A5 were independently isolated from two different representatives of the order Holothuriida. These 104 new glycosides were isolated as individual substances and their structures were elucidated using 1D and 2D NMR procedures. The majority of them were found in sea cucumbers belonging to different families of the order Dendrochirotida. Although many glycosides are members of the holostane series, 57 are nonholostane derivatives. Two glycosides have rearranged lanostane carbocyclic skeletons as a result of intramolecular aldol condensation and Meinwald rearrangement. Nineteen glycosides are either lanostane or nor-lanostane derivatives without any lactone and 15 are either lanostane or nor-lanostane derivatives with an 18(16)-lactone. Thus, the diversity of uncommon aglycones of glycosides, especially from sea cucumbers belonging to the order Dendrochirotida, is very impressive and provides good perspectives for further structural investigation of carefully isolated and purified individual substances.
Taxonomic Distribution of new Triterpene Glycosides Found in sea Cucumbers From 2017 to 2021.
Stichorrenosides A (95), B (96), C (97), D (98), 3β-O-[β-D-xylopyranosyl-(1→2)-β-D-xylopyranosyl]-23S-acetoxyholost-7-ene (99), 3β-O-[β-D-xylopyranosyl-(1→2)-β-D-xylopyranosyl]-23S-hydroxyholost-7-ene (100)
42, Figure 25. The coincidence of the names of two different glycosides 108 and 109 is random.
The diversity of carbohydrate chains is very impressive and several dozen different kinds of them have been found. The most interesting findings were the differences in numbers and positions of sulfate groups. Even highly polar carbohydrate chains with four sulfates were discovered, and a lot of positions of the sulfates were revealed in addition to the common C-6 of glucose and 3-O-methylglucose, and C-4 of the first xylose residues.
The majority of the sea cucumber triterpene glycosides isolated between 2017 to 2021 possess common activities characteristic for this class of natural products, namely cytotoxicity towards different kinds of cells such as erythrocytes and tumor cells. The glycosides may induce apoptosis in tumor cells, and inhibit the formation and growth of tumor cell colonies in agar media in sub-toxic doses. The glycosides also possess antitumor activities in vivo on homograft and xenograft tumor models. A very interesting new kind of activity was found recently: the ability of some glycosides to decrease the effective doses of radiation that may be used for radiation therapy against human tumors.
The structure-activity relationships for cytotoxic activities of the new glycosides are very complicated and depend on both aglycone and carbohydrate chain structures. Generally, the presence of an 18(20)-lactone, 16β-OAc, linear tetrasaccharide chains, sulfates at C-4 of the first xylose residue, and the presence of quinovose as the second monosaccharide residue in the carbohydrate chain are very significant for the biological activity. It was found that the presence of a hydroxy group on the aglycone side chain dramatically decreases the activities, and the presence of two sulfates at the same monosaccharide residue has the same effect. The most interesting finding was the moderate cytotoxic activities of several non-holostane glycosides without a lactone and even with shortened side chain or having an 18(16)-lactone. All these findings reveal the necessity of further detailed studies on modelling of possible different mechanisms of the membranotropic action of glycosides using an in silico approach.
Thus, the results of studies during the last five years show that sea cucumber triterpene glycosides continue to be a subject for this type of investigation from both the structural and functional points of view.
Footnotes
Acknowledgments
The reported study was supported by the Russian Foundation for Basic Research No. 19-04-000-14.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Russian Foundation for Basic Research (grant number No. 19-04-000-14.).
Ethical Approval
Not applicable, because this article does not contain any studies with human or animal subjects.
Informed Consent
Not applicable, because this article does not contain any studies with human or animal subjects.
Trial Registration
Not applicable, because this article does not contain any clinical trials.
ORCID iD
Vladimir I. Kalinin
References
1.
MondolMAMShinHJRahmanMA. Sea cucumber glycosides: chemical structures, producing species and important biological properties. Mar Drugs. 2017;15(10):e317. doi:10.3390/md15100317
2.
Honey-EscandonMArreguin-EspinosaRSolis-MartinFASamynY. Biological and taxonomic perspective of triterpenoid glycosides of sea cucumbers of the family Holothuriidae (Echinodermata, Holothuroidea). Comp Biochem Physiol. 2015;180B:16‐39. doi:10.1016/j.cbpb.2014.09.007
3.
BahramiYFrankoCMM. Acetylated triterpene glycosides and their biological activity from Holothurioidea reported in the past six decades. Mar Drugs. 2016(8);14:e147. doi:10.3390/md14080147
4.
StonikVAKalininVIAvilovSA. Toxins from the sea cucumbers (Holothuroids): chemical structures, properties, taxonomic distribution, biosynthesis and evolution. J Nat Toxins. 1999;8(2):235‐248.
5.
KalininVISilchenkoASAvilovSAStonikVASmirnovAV. Sea cucumbers triterpene glycosides, the recent progress in structural elucidation and chemotaxonomy. Phytochem Rev. 2005;4(2,3):221‐236. doi:10.1007/s11101-005-1354-y
ZhaoY-CXueC-HZhangTTWangY-M. Saponins from sea cucumber and their biological activities. Agr.Food Chem. 2018;66(28):7222‐7237. doi:10.1021/acs.jafc.8b01770
8.
KalininVIAmininDLAvilovSASilchenkoASStonikVA. Triterpene glycosides from sea cucumbers (holothurioidae, echinodermata), biological activities and functions. In: Atta-ur-Rahman (ed.). Studies in natural product chemistry (bioactive natural products). 35:135‐196. Elsevier Science Publisher; 2008.
9.
AmininDLPislyaginEAMenchinskayaESSilchenkoASAvilovSAKalininVI. Immunomodulatory and anticancer activity of sea cucumber triterpene glycosides. In: Atta-ur-Rahman (ed.). Studies in natural products chemistry. 41:75‐94. Elsevier Science B.V.; 2014.
10.
AmininDLMenchinskayaESPisliaginEASilchenkoASAvilovSAKalininVI. Anticancer activity of sea cucumber triterpene glycosides. Mar Drugs. 2015;13(10):1202‐1223. doi:10.3390/md13031202
11.
AmininDLMenchinskayaESPisliaginEASilchenkoASAvilovSAKalininVI. Sea cucumber triterpene glycosides as anticancer agents. In: Atta-ur-Rahman (ed.). Studies in natural product chemistry. 49:55‐105. Elsevier B.V.; 2016.
12.
LiYWangRXunX, et al. Sea cucumber genome provides insights into saponin biosynthesis and aestivation regulation. Cell Discov. 2018;4:e29. doi:10.1038/s41421-018-0030-5
13.
LiuHKongXChenJZhangH. De novo sequencing and transcriptome analysis of Stichopus horrens to reveal genes related to biosynthesis of triterpenoids. Aquaculture. 2018;491:358‐367. doi:10.1016/j.aquaculture.2018.01.012
14.
ChenXShaoXLiWZhangXYuB. Total synthesis of echinoside A, a representative triterpene glycoside of sea cucumbers. Angewandte Chemie. International Edition. 2017;56(26):7648‐7653. doi:10.1002/anie.201703610
15.
ShaoXWangXZhuKDangYYuB. Synthesis of sea cucumbers saponins with antitumor activities. J Org Chem. 2020;85(19):12080‐12096. doi:10.1021/acs.joc.0c01191
16.
SilchenkoASKalinovskyAIAvilovSA, et al. Fallaxosides B1 and D3, triterpene glycosides with novel skeleton types of aglycones from the sea cucumber Cucumaria fallax. Tetrahedron. 2017;73(17):2335‐2341. doi:10.1016/j.tet.2017.02.041
17.
SilchenkoASKalinovskyAIAvilovSA, et al. Structures and bioactivities of quadrangularisosides A, A1, B, B1, B2, C, C1, D, D1–D4, and E from the sea cucumber Colochirus quadrangularis: the first discovery of the glycosides, sulfated by C-4 of the terminal 3-O-methylglucose residue. Synergetic effect on colony formation of tumor HT-29 cells of these glycosides with radioactive irradiation. Mar Drugs. 2020;18(8):e394. doi:10.3390/md18080394
18.
YangW-SQiX-RXuQ-Z, et al. A new sulfated triterpene glycoside from the sea cucumber Colochirus quadrangularis, and evaluation of its antifungal, antitumor and immunomodulatory activities. Bioorg Med Chem. 2021;41:e116188. doi:10.1016/j.bmc.2021.116188
19.
YiY-HXuQ-ZLiL, et al. Philinopsides A and B, two new sulfated triterpene glycosides from the sea cucumber.Helv Chim Acta. 2006;89(1):54‐63. doi:10.1002/hlca.200690012
20.
SilchenkoASKalinovskyAIAvilovSA, et al. Kurilosides A1, A2, C1, D, E and F – triterpene glycosides from the Far Eastern sea cucumber Thyonidium (=Duasmodactyla) kurilensis (Levin): structures with unusual non-holostane aglycones and cytotoxicities. Mar Drugs. 2020;18(11):e551. doi:10.3390/md18110551
21.
SilchenkoASKalinovskyAIAvilovSA, et al. Triterpene glycosides from the Far Eastern sea cucumber Thyonidium (=Duasmodactyla) kurilensis (Levin): the structures, cytotoxicities, and biogenesis of kurilosides A3, D1, G, H, I, I1, J, K, and K1. Mar Drugs. 2021;19(4):e187. doi:10.3390/md19040187
22.
KalininVIStonikVA. Application of morphological trends of evolution to phylogenetic interpretation of chemotaxonomic data. J Theor Biol. 1996;180(1):1‐10. doi:10.1006/jtbi.1996.0073
23.
SilchenkoASKalinovskyAIAvilovSA, et al. Cladolosides I1, I2, J1, K1 and L1, monosulfated triterpene glycosides with new carbohydrate chains from the sea cucumber Cladolabes schmeltzii. Carbohydr Res. 2017;445:80‐87. doi:10.1016/j.carres.2017.04.016
24.
SilchenkoASKalinovskyAIAvilovSA, et al. Cladolosides C4, D1, D2, M, M1, M2, N and Q, new triterpene glycosides with diverse carbohydrate chains from sea cucumber Cladolabes schmeltzii. An uncommon 20,21,22,23,24,25,26,27-okta-nor-lanostane aglycone. The synergism of inhibitory action of non-toxic dose of the glycosides and radioactive irradiation on colony formation of HT-29 cancer cells. Carbohydr Res. 2018;468:36‐44. doi:10.1016/j.carres.2018.08.003
25.
SilchenkoASKalinovskyAIAvilovSA, et al. Cladolosides O, P, P1–P3 and R, triterpene glycosides with two novel types of carbohydrate chains from the sea cucumber Cladolabes schmeltzii. Inhibition of cancer cells colony formation and its synergy with radioactive irradiation. Carbohydr Res. 2018;468:73‐79. doi:10.1016/j.carres.2018.08.004
26.
KalininVI. System-theoretical (holistic) approach to the modelling of structural-functional relationships of biomolecules and their evolution: an example of triterpene glycosides from sea cucumbers (Echinodermata, Holothurioidea). J Theor Biol. 2000;206(1):151‐168. doi:10.1006/jtbi.2000.2110
27.
PopovRSIvanchinaNVSilchenkoAS, et al. Metabolite profiling of triterpene glycosides of the Far Eastern sea cucumber Eupentacta fraudatrix and their distribution in various body components using LC-ESI QTOF-MS. Mar Drugs. 2017;14(10):e302. doi:10.3390/md15100302
28.
SilchenkoASKalinovskyAIAvilovSA, et al. Triterpene glycosides from the sea cucumber Eupentacta fraudatrix. Structure and cytotoxic action of cucumarioside D with a terminal 3–O-Me-glucose residue unique for this species. Nat Prod Commun. 2018;13(2):137‐140.
29.
MalyarenkoTVKichaAAKalinovskyAI, et al. New triterpene glycosides from the Far eastern starfish Solaster pacificus and their biological activity. Biomolecules.2021;11(3):e427. doi:10.3390/biom11030427
30.
SilchenkoASAvilovSAKalinovskyAIKalininVIAndrijaschenkoPVDmitrenokPS. Psolusosides C1, C2 and D1, novel triterpene hexaosides from the sea cucumber Psolus fabricii (Psolidae, Dendrochirotida). Nat Prod Commun. 2018;13(12):1623‐1628.
31.
SilchenkoASAvilovSAKalinovskyAI, et al. Psolusosides C3 and D2–D5, five novel triterpene hexaosides from the sea cucumber Psolus fabricii (Psolidae, Dendrochirotida): chemical structures and bioactivities. Nat Prod Commun. 2019;14(7):1-12. doi:10.1177/1934578X19861253.
32.
KalininVIKalinovskiiAIStonikVADmitrenokPSEl’kinYN. Structure of psolusoside B – A nonholostane triterpene glycoside from the holothurian genus Psolus. Chem Nat Compd. 1989;25(3):311‐317. doi:10.1007/BF00597709
33.
SilchenkoASKalinovskyAIAvilovSA, et al. Structures and bioactivities of six new triterpene glycosides, psolusosides E, F, G, H, H1 and I and the corrected structure of psolusoside B from the sea cucumber Psolus fabricii. Mar Drugs. 2019:17(6):e358. doi:10.3390/md17060358
34.
SilchenkoASKalinovskyAIAvilovSA, et al. Structures and bioactivities of psolusosides B1, B2, J, K, L, M, N, O, P, and Q from the sea cucumber Psolus fabricii. The first finding of tetrasulfated marine low molecular weight metabolites. Mar Drugs. 2019;17(11):e631. doi:10.3390/md17110631
35.
SilchenkoASKalinovskyAIAvilovSA, et al. Nine new triterpene glycosides, magnumosides A1–A4, B2, C1, C2 and C4, from the Vietnamese sea cucumber Neothyonidium (=Massinum) magnum: structures and activities against tumor cells independently and in synergy with radioactive irradiation. Marine Drugs. 2017;15(8):e256. doi:10.3390/md15080256
36.
SilchenkoASKalinovskyAIAvilovSA, et al. Magnumosides B3, B4 and C3, mono- and disulfated triterpene tetraosides from the Vietnamese sea cucumber Neothyonidium (=Massinum) magnum. Nat Prod Commun. 2017;12(10):1577‐1582.
37.
CuongNXVienLTHoangL, et al. Cytotoxic triterpene glycosides from the sea cucumber Stichopus horrens. Bioorg Med Chem Let.2017;27(13):2939‐2942. doi:10.1016/j.bmcl.2017.05.003
38.
VienLTHoangLHanhTTH, et al. Triterpene tetraglycosides from the sea cucumber Stichopus horrens. Nat Prod Res. 2018;32(9):1039‐1043. doi:10.1080/14786419.2017.1378206
39.
ZhangX-MLiX-BZhangS-S, et al. LC-MS/MS identification of novel saponins from the viscera of sea cucumber Apostichopus japonicus.Chem Nat Comp. 2018;54:721‐725. doi:10.1007/s10600-018-2454-4
40.
ZhangX-MHanL-WShengW-L, et al. Two novel non-holostane type glycosides from the viscera of sea cucumber Apostichopus japonicus. J Asian Nat Prod Res. 2020;22(4):329‐337. doi:10.1080/10286020.2019.1576643
41.
HoangaLVienLTHanhTTH, et al. Triterpene glycosides from the Vietnamese sea cucumber Holothuria edulis.Nat Prod Res. 2020;34(8):1061‐1067. doi:10.1080/14786419.2018.1548451
42.
GrauzoLYegdanehASharifiMMangoniAZolfaghariBLanzottiV. Molecular networking-based analysis of cytotoxic saponins from sea cucumber Holothuria atra. Mar Drugs. 2019;17(2):e86. doi:10.3390/md17020086
43.
HawasUWAbou El-KassemLTShaherFMGhandourahMAl-FarawatiR. Sulfated triterpene glycosides from the Saudi Red Sea cucumber Holothuria atra with antioxidant and cytotoxic activities. Thalassas. 2021;37:817-824. doi:10.1007/s41208-021-00305-4.