
Abstract
Naturally occurring fires have been shaping landscapes long before mankind existed. Presumably, it is an early observation that in some habitats, fire is crucial to maintaining species diversity and for the rejuvenation of the vegetation, and so early settled farmers might have started to take advantage of the controlled burning of a desired area. The heat of fire is essential to break the dormancy of many fire ephemerals and for the seed release of some serotinous or woody taxa. Besides the physical effects caused by the heat on seeds, smoke and burnt organic material contain chemical cues that regulate the germination of seeds and the early development of seedlings. The scientific community really started to reveal the secrets of these enigmatic components from the early 1990s, although there are still a number of questions to be answered. In this review, we briefly introduce the path which leads to our current knowledge on smoke-derived compounds and their enormous effects on plant life.
Periodic wildfires are a natural phenomenon with a frequency of 1 to 40 years, although fire occurrence increases due to anthropogenic activity. 1 Fire, which is an integral part of many terrestrial ecosystems, is essential to maintain healthy vegetation dynamics, in particular by promoting synchronous germination of fire-followers and sprouting of bulbs, rhizomes, and woody species (Figure 1(a), upper panel). The most spectacular change is definitely the germination burst of the so-called fire ephemerals in the postfire landscape. This positive effect of fire has been widely used in agriculture either to promote germination or seedling vigor (Figure 1(b), lower panel). In this review, we present a brief history of the efforts to identify germination-promoting, fire-borne chemical cues.

(a) Upper panel: Postfire landscape in the Drakensberg, South Africa. Fire promoted a rapid sprouting in the burnt area after a heavy spring rainfall. Lower panel: Tomato seedlings grown on charred wood and agricultural waste in Siby, Mali. Local farmers use charred wood to stimulate germination and improve seedling vigor. (b) The fynbos, the characteristic vegetation type of the Cape Floristic Region (Cape Peninsula, South Africa). A high proportion of fynbos species is responsive to smoke. All pictures by Vilmos Soós.
Fire provides physical and chemical cues by nature to promote germination in many disparate taxa both in fire-prone and fire-free habitats across the globe. It is clear that the heat produced by fire has the most enormous effect on seeds, mainly by drying and cracking the seed coat, and to a lesser extent, by stimulating embryo growth directly. 2 Heat can break dormancy either by weakening hard seed coats or by disrupting any structures blocking radicle emergence, such as chalazal plugs. 3 Besides heat, fire provides a set of chemical cues which potentially act on seeds in the soil seed bank or on seeds stored in the canopy. Together with physical cues, chemical signals may fine-tune germination time following fire so as to occur when germination or sprouting is optimal. 4 These fire-ephemeral plants pursue a very successful strategy wherein fire eliminates canopy, potential competitors and the ashes of former vegetation enrich the soil with nutrients providing a temporarily open habitat for thriving. Sensing the chemical signal released by the fire therefore is an ancient, adaptive trait, developed even by phylogenetically distant taxa.
Originally, gases released by fire were regarded as the primary chemical stimulants. Although ammonia and ethylene 5 -7 are known active germination cues, and both may be produced in fire, it was shown that ethylene is not necessarily the compound in smoke solutions that is responsible for germination activity, 8 since ethylene did not increase seed germination to the same extent as smoke. Thus, it was theorized that nutrients, especially nitrogenous forms released from the burned vegetation and chars, could mostly account for the rapid sprouting in the postfire landscape. It is well known that fire has a tremendous effect on the soil total nitrogen content, and ammonium formed in fire can trigger the germination of some fire followers, like Phacelia grandiflora and Salvia mellifera. 9 Organic nitrogen and ammonium could also be transformed into nitrate during the nitrification process by soil-borne bacteria. Such microorganisms require enough moisture and a neutral-basic pH, thus conditions for nitrification are more favorable when the wet season follows fire, which slightly increase the soil pH. 10,11 The fortunate constellation of these conditions temporarily elevates nitrate levels which eventually promotes germination of many taxa of fire-prone habitats. 9 However, nitrate accumulation sometimes is too low to promote germination, and in the chaparral region, it is not the nitrate which accounts for the rapid sprouting of the postfire flora. 12,13
In 1990, De Lange and Boucher 14 reported that smoke-water, a leachate from charred plant material or smoke bubbled through water, contained a chemical(s) stimulating agent indicating that smoke is the major source of activity. This finding captured the imagination of ecologists and plant biologists, and it became clear that these fire-borne chemical signals in smoke are transmissible and could reach plant communities’ distances away from the origin of the fire. 15 Furthermore, the signal could persist for considerable periods of time as the active compound(s) could be deposited in soil, absorbed by soil particles, and released slowly in water. Thus, fire not only influences seed banks and the immediate postfire environment but it can also have an enormous effect lasting for extended periods of time. 15 To date, as many as 1335 species from more than 120 families were shown to be smoke-responsive (Figure 1(b) 16 ).
Nitrous oxide (NO) was the first reported fire-derived chemical cue which promoted the germination of Emmenanthe penduliflora, a characteristic chaparral annual. 12 However, not all fire-responsive pyroendemics tested were stimulated by NO, indicating that multiple chemical cues may be responsible for the promotion of germination in postfire environments. Another line of evidence indicated that NO-free smoke, or smoke from low intensity fires where an insufficient amount of NO was generated, could also stimulate germination, 17,18 suggesting that it is not NO that solely accounted for the germination activity. Another nitrogen containing chemical cue formed by slow combustion of plant material is glyceronitrile, which is hydrolysed in water to release cyanide, a well-known germination promoting agent of multiple plant species, apart from its toxic effects on living organisms. 19 -21 Taken together, these findings supported the hypothesis that not only nutrient-containing gases and leachates were a source of germination-promoting activity but that other chemicals that were likely to act as major, universal stimuli were also involved. Such chemicals are present in smoke as a “message” which can drift large distances away from the source of fire. These air-borne, volatile compounds are water soluble since smoke bubbled through water also retains germination activity. 14 The production of smoke-water opened the way for commercialization of the germination stimulatory agent, and also made possible to standardize smoke solutions for research. 2 Furthermore, these seemingly stable cues can absorb to surfaces including biochars and seeds, although rinsing could also dilute or remove the chemicals. 2 Originally, it was surmised that the mysterious cue is formed exclusively by the combustion of pyroendemics found in fire-prone habitats. However, it was evident that the combustion (burning) of almost any kind of plant material is capable of producing these stimuli as well as the combustion of cellulose (paper), agar, simple carbohydrates, pyromeconic acid, and propionic acid to produce germination active chemicals. 22 Interestingly, the phylogenetic spread of smoke-responsive species is wide and includes many nonfire follower species, suggesting that this trait is an early developmental feature of angiosperms. 16,23
Early attempts to isolate the active compound(s) from smoke were unsuccessful. By means of bioassay guided fractionation, 71 different compounds were found in smoke, but unfortunately, none of them were proven to promote germination in the postfire annual Nicotiana attenuate, which was used as a model plant. 24 Similarly, 12 compounds were found in smoke-water from the combustion of Themeda triandra and Passerina vulgaris, and also, none of them were active in the lettuce bioassay system. 25 Nonetheless, these attempts highlighted an important feature of the fire-borne chemical cue(s), namely, that it is present in smoke in extremely low quantities, a trait which probably hindered its identification. Efforts to identify the principal fire-borne, germination promoting cue(s) were finally rewarded in 2004 when simple butenolides were shown to be responsible for the activity by the Dixon group, 26 and confirmed shortly thereafter by the van Staden research group. 27 The first identified germination active compound was 3-methyl-2H-furo[2,3 c]pyran-2-one, which is now referred to as karrikinolide (KAR1), having been named as such after the aboriginal word for smoke, which is “karrik.” So far, 5 analogs of karrikinolide have been isolated from smoke (KAR2-KAR6), though KAR1 to KAR4 are the most active ones. 28,29 These germination promoting constituents of smoke have a butenolide fused to a pyran ring. Supposedly, they are produced from polysaccharides and sugars (pyranose) by combustion in the presence of oxygen. Although one can assume that karrikins are heat stable, it seems that this is not the case. 30 An important chemical feature of the karrikins is the butenolide ring, which was immediately noted upon their discovery, since it structurally resembles strigolactones, novel plant hormones involved in various physiological and developmental processes in plants. 26,27 Karrikins can mimic a still elusive, enigmatic endogenous substance, called karrikin ligand which, by binding to the karrikin receptor KAI2, elicits a signal transduction machinery through MAX2 and SMAX1, SMXL2 proteins to regulate multiple developmental processes such as leaf development, mycorrhiza, and light responses. 31 Hence, the identification of karrikins paved the way to the discovery of a novel signaling pathway, which is a separate, but mechanistically similar to the strigolactone signaling orchestrated by the D14 protein. 31
In addition to karrikins, other fire-borne active compounds have been subsequently identified. These have been shown to either influence seedling growth, such as catechol, 32 or promote germination, like hydroquinone. 33 The finding that not all smoke-responsive species responded to KAR treatment has led to the presumption that other active compounds may be present and may interact with KAR. 34 -36 Very little is known about the orchestrated, interactive effects of various smoke-derived compounds, which might be synergistic or antagonistic. 36 The most active compounds (such as karrikins and glyceronitrile) possibly act in concert and are likely to be responsible for the overall effect of smoke on germination or seedling vigor.
It has been well noted that concentrated smoke-water inhibits germination, 37 whereas the more diluted solutions generally promoted germination. This “dual regulatory” effect consequently led to the speculation that inhibitory cues could also be present in smoke. The search for the enigmatic inhibitory compound revealed another small butenolide, 3,4,5‐trimethylfuran‐2(5H)‐one (trimethylbutenolide; TMB), which is structurally related to karrikins, 38 as well as 2 other furanones which also inhibit germination. 39 Interestingly, TMB is more water soluble than karrikins and is present in smoke in much larger quantities. 34 These findings support the concept of the “dual regulatory” effect wherein TMB might act as a “release” signal: once fire occurs, TMB accumulates in the soil until the next heavy rainfall, which dilutes the inhibitory compound below a threshold level. 35 In this model, TMB impedes germination, while the other compounds have a promotive role, although their effect is masked by the inhibitor. However, once the TMB is sufficiently reduced by washing away, the germination promotion cues can exert their activity. The synergistic-inhibitory relationship of these diverse compounds can, therefore, fine-tune germination time and adjust seedling establishment to sufficient rainfall to optimize seedling survival (Figure 2 35 ). There are a number of proposed technologies to utilize the knowledge of these chemical stimulants in farming practices, habitat restoration, and natural resource management. 40
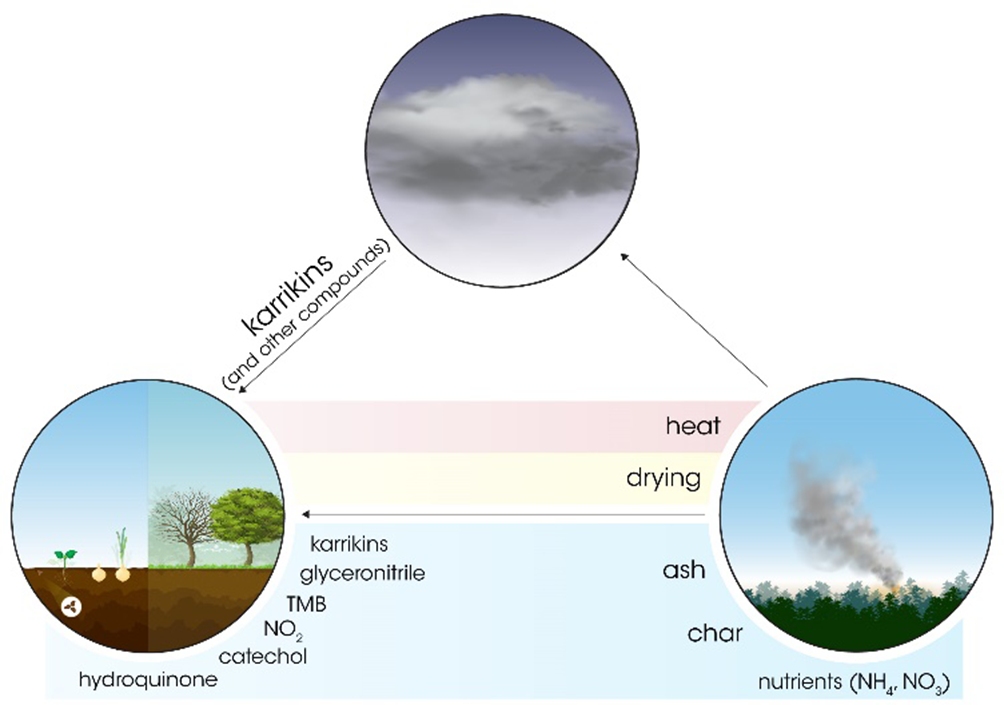
The positive effects of wildfires on the vegetation.
Since strigolactones and karrikins have been shown to promote the germination of the parasitic weed Striga spp. and Orobanche spp., it has been proposed to be used as a stimulant to induce suicidal germination of these harmful species. 41 -44 Similarly, karrikins or smoke-water could also be used to initiate an early, synchronized germination of (even dormant) weed seeds, following which a mechanical or chemical weed control could be done, thus enhancing depletion of the weed seed bank in the soil. 45 The germination promoting activity of plant-derived smoke has been observed on several horticulturally important plants such as tomato, celery, or lettuce. It has been demonstrated that soaking seeds either in diluted smoke-water or in very low concentrations of KAR1 stimulates germination, 26 even at nonoptimal temperatures. 46 Smoke-water has been commercially used in the form of seed pads and solutions (seed primers), and the use of biochar may also offer an ecologically beneficial solution for agriculture, reforestation, or habitat restoration. However, care should be taken to understand the potentially complex effects of these treatments on the overall environment and ecology where they may be utilized. 47
In conclusion, understanding how smoke affects germination not only has a huge impact on the ecological research of fire-prone habitats but has also revealed a novel, still enigmatic signaling pathway, which is apparently universal in the plant kingdom. Furthermore, the use of smoke-water might provide a new set of methods to improve crop productivity and yield, especially for resource-poor farmers in the developing countries. However, there is still a great deal of research to be done to fully understand how plants sense and respond to these fire-borne chemical signals.
Footnotes
Acknowledgment
Thanks are due to Marnie E. Light for the useful comments and language editing.
Author Note
On the occasion of the 80th birthday of Professor van Staden appreciating his pioneering work on smoke research.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Fundings were provided by Hungarian Scientific Research Fund (OTKA K128644 and NN114567).