
Abstract
Extremophiles are a rich source of novel secondary metabolites and valuable antioxidative compounds. Here, we examined the antioxidative capacities of aqueous extracts derived from the halotolerant cyanobacterium Halothece sp. PCC7418. The antioxidative activities of extracts derived from Halothece sp. PCC7418 cells exposed to temperature shock were significantly enhanced. Quantification of antioxidant molecules (phenolic compounds and phycobiliproteins) in the aqueous extracts revealed that the amounts of these molecules were modulated by temperature shock. In addition, the intracellular accumulation of mycosporine-2-glycine, a potential antioxidative molecule, was strongly enhanced by cold shock. Our results show that the treatment of Halothece sp. PCC7418 cells with temperature shock may allow for the robust production of antioxidants.
Reactive oxygen species (ROS) are generated in all living organisms as a result of both normal cellular metabolism and a variety of environmental factors. Reactive oxygen species can induce oxidative modification of various cell structures (eg, carbohydrates, nucleic acids, lipids, and proteins), which alters their functions. 1 In order to protect against ROS-induced cell damage, organisms possess several antioxidant systems, which include both enzymatic and nonenzymatic components. Various nonenzymatic molecules, which function as antioxidants, have been previously characterized. Antioxidants can neutralize the harmful effects of ROS and thus help in preventing oxidative stress. With regard to human usage, the demand for natural product-derived antioxidants is growing because natural products are proven to be relatively safe for human consumption compared to synthetic compounds. Historically, plants served as the primary source for natural bioactive compounds including antioxidants. In addition to plants, cyanobacteria have also gained much attention as a promising group of microorganisms capable of producing industrially important compounds due to their photoautotrophic property, higher growth rate, and genetic manipulability. 2 Cyanobacteria can produce and accumulate various antioxidative molecules. 3 Recently, several reports have described the antioxidative activities and properties of extracts derived from terrestrial and freshwater cyanobacteria. 4 -7 However, the relationship between the antioxidative activities and the antioxidant-generating capacities of these cyanobacterial extracts is not fully understood. Moreover, a much needed resource for antioxidants, such as extremophilic cyanobacteria, has not yet been reported. In this study, we evaluated the antioxidative activities of aqueous extracts from the halotolerant cyanobacterium Halothece sp. PCC7418, firstly identified as Aphanothece halophytica. 8 Substantial evidence has documented several potential biocompounds produced by this cyanobacterium. 9 -13 When Halothece sp. PCC7418 cells were cultured under temperature shock conditions, the antioxidative capacities of their aqueous extracts were significantly enhanced. Herein, we describe the relationship between the antioxidative properties of these extracts and the dynamics of potential antioxidative molecules (including polyphenols, phycobiliproteins [PBPs], and mycosporine-like amino acids [MAAs]) under temperature stress conditions.
Cold Stress Induces the Antioxidative Capacity of Aqueous Extracts Derived From Halothece sp. PCC7418 Cells
We investigated the effects of culture temperature on the antioxidative activity of aqueous extracts derived from Halothece sp. PCC7418 cells. Halothece sp. PCC7418 cells were cultured with or without exposure to temperature shock for 7 days under higher (2.5 M) or lower (0.5 M) concentrations of NaCl. The temperature for culturing cells was shifted from 30°C to 23°C or 37°C for cold or heat shock, respectively. It should be noted that the maximum growth of Halothece sp. PCC7418 was obtained at 30°C. At temperatures below 23°C or above 37°C, the growth dropped gradually, which likely resulted from low- or high-thermal stress, respectively. Interestingly, a 2,2-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt (ABTS) assay using aqueous extracts prepared from Halothece sp. PCC7418 cells revealed that both cold and heat shocks enhanced the antioxidative activity, which was expressed as Trolox equivalents (TEs, Table 1). In cells grown under lower or higher NaCl conditions, cold shock significantly increased the antioxidative activity by 1.52- and 1.28-fold under conditions of 0.5 or 2.5 M NaCl, respectively. On the other hand, heat shock elevated antioxidative activity by 1.45-fold only in the case of cells grown under 0.5 M NaCl conditions. These observations prompted us to identify the antioxidative molecules in each extract subjected to temperature shock.
Total Phenolic Content, Total Flavonoid Content, and Antioxidative Power of Halothece sp. PCC7418 Extracts From Cells Grown Under Cold or Heat Stress Conditions.

ABTS, 2,2-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt; GAEs, gallic acid equivalents; QEs, quercetin equivalents; TEs, total equivalents; TFC, total flavonoid content; TPC, total phenolic content.
Significant differences between extracts derived from cells exposed to temperature shock and control (Cntl) extracts are indicated as *P < 0.05 and **P < 0.01, respectively; “-” indicates no significant difference.
Thermal Stress-Induced Modulation of Potential Antioxidant Molecule Accumulation in Halothece sp. PCC7418 Extracts
A variety of polyphenols, consisting of mainly flavonoids and phenolic acids, are known as powerful antioxidants. 14 As shown in Table 1, the phenolic contents of Halothece sp. PCC7418 extracts were examined by determining the total phenolic content (TPC) and the total flavonoid content (TFC). The TPC values were expressed in gallic acid equivalents (GAEs µg/mgFW). The TFC values were expressed in quercetin equivalents (QEs µg/mgFW). We found that both the TPC and the TFC were modulated by both cold and heat stress. Under both NaCl concentration conditions, heat shock enhanced the TPC by 1.49- and 1.44-fold in cells grown under 0.5 and 2.5 M NaCl conditions, respectively. In contrast, although the TPC content was increased by 1.25-fold in cold-shocked cells grown under 2.5 M NaCl conditions, in cold-shocked cells grown under 0.5 M NaCl conditions, the TPC was decreased by 1.13-fold. These results were not in accord with the modulation of antioxidative properties by these temperature shocks (Table 1). Thus, the TPC is not a direct indicator of the antioxidative power of each extract. In addition, the TFC showed a fundamentally similar pattern as the TPC except that the TFC remained unchanged in cold-shocked cells grown under 0.5 M NaCl conditions.
Next, the amounts of PBPs, which are known as water-soluble strong antioxidants, 4 were also quantified. In this study, the phycoerythrin (PE), phycocyanin (PC), and allophycocyanin (APC) contents were determined (Table 2). Under both NaCl concentration conditions, heat shock significantly enhanced the total PBP (PE +PC + APC) content by 2.48- and 2.28-fold in cells grown under 0.5 and 2.5 M NaCl conditions, respectively. On the other hand, similar to the TPC results, cold shock increased the total PBP content by 1.54-fold in cells grown under 2.5 M NaCl conditions, but the total PBP content was decreased by 1.88-fold in cold-shocked cells grown under 0.5 M NaCl conditions. This reduction in total PBP content was not consistent with the increase in antioxidative activity under the same conditions (Table 1). Thus, the PBPs may not be main responsible compounds for the antioxidative power of each Halothece sp. PCC7418 aqueous extract. Among the extracts tested in this study, the total PBP content was highest (58.50 µg/mgFW) in the extract from the heat-treated cells cultured under 0.5 M NaCl conditions. In addition, under these same conditions, the PC content was remarkably enhanced from 14.32 µg/mgFW in cells grown at 30°C to 39.78 µg/mgFW in cells subjected to heat shock at 37°C.
Phycobiliprotein Content in Halothece sp. PCC7418 Extracts From Cells Grown Under Cold or Heat Stress Conditions.
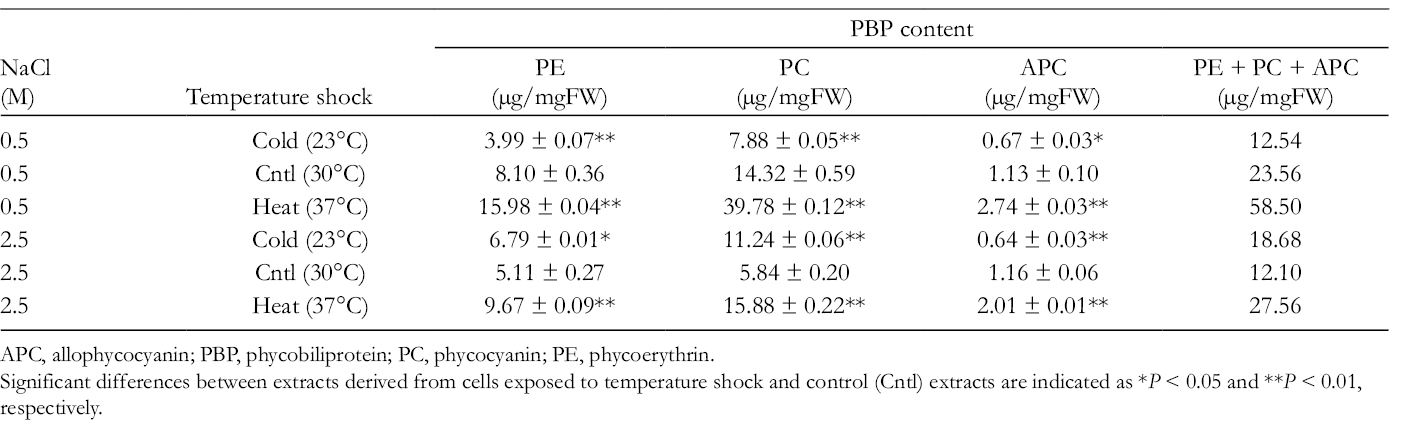
APC, allophycocyanin; PBP, phycobiliprotein; PC, phycocyanin; PE, phycoerythrin.
Significant differences between extracts derived from cells exposed to temperature shock and control (Cntl) extracts are indicated as *P < 0.05 and **P < 0.01, respectively.
Halothece sp. PCC7418 is able to bioproduce mycosporine-2-glycine (M2G), which is one member of MAA family. 9 MAAs including M2G are recognized as UV sunscreen compounds. The chemical structure of M2G is shown in Figure 1(a). In addition, accumulating evidence has shown that MAAs possess other functions, such as antioxidative activity 15 and osmoprotective property. 16,17 Previously, we demonstrated that M2G possessed stronger antioxidative activity than other MAAs, such as porphyra-334 and shinorine. 13,18,19 Since M2G is a superior contributor to antioxidative activity, we investigated the accumulation patterns of M2G in Halothece sp. PCC7418 cells grown under temperature stress conditions (Figure 2a,b). As shown in Figure 2, cold stress induced the accumulation of M2G in cells grown under both 0.5 and 2.5 M NaCl conditions. The M2G accumulation level was sharply increased by 3.5-fold (from 0.58 to 2.03 nmol/mgFW) in cells grown under conditions of 2.5 M NaCl at day 7. In addition, the M2G accumulation level increased by 2.6-fold (from 0.16 to 0.41 nmol/mgFW) in cells grown under conditions of 0.5 M NaCl at day 7. On the other hand, heat stress increased M2G accumulation by 2-fold (from 0.16 to 0.32 nmol/mgFW) in cells grown under conditions of 0.5 M NaCl at day 7. In contrast, M2G levels remained unchanged in heat-shocked cells grown under 2.5 M NaCl conditions at day 7 (Figure 2). To the best of our knowledge, this is the first demonstration that cold stress strongly enhances intracellular M2G content. In analogy with the case of salt stress treatment, 9 it is possible that the expression of M2G biosynthetic genes was upregulated by cold stress. Investigation of the expression profile of M2G biosynthetic genes under various abiotic stresses is an important future research subject. The induction of M2G by cold stress may be correlated with the higher antioxidative activities of the extracts prepared from cold-shocked cells (Table 1). On the other hand, TPC, TFC, and PBP content in cold-stressed extracts were lower than that in heat-stressed extracts (Tables 1 and 2). Next, we determined the intracellular levels of glycine betaine (GB), which is known as a major osmoprotectant exclusively produced in Halothece sp. PCC7418 (Figure 2). The chemical structure of GB is shown in Figure 1. We found that both cold and heat shocks enhanced GB content. The analysis of cyanobacterial secondary metabolite levels upon reversal changes in temperature would be an interesting subject to be clarified in future studies.

Chemical structures of mycosporine-2-glycine (a) and glycine betaine (b).
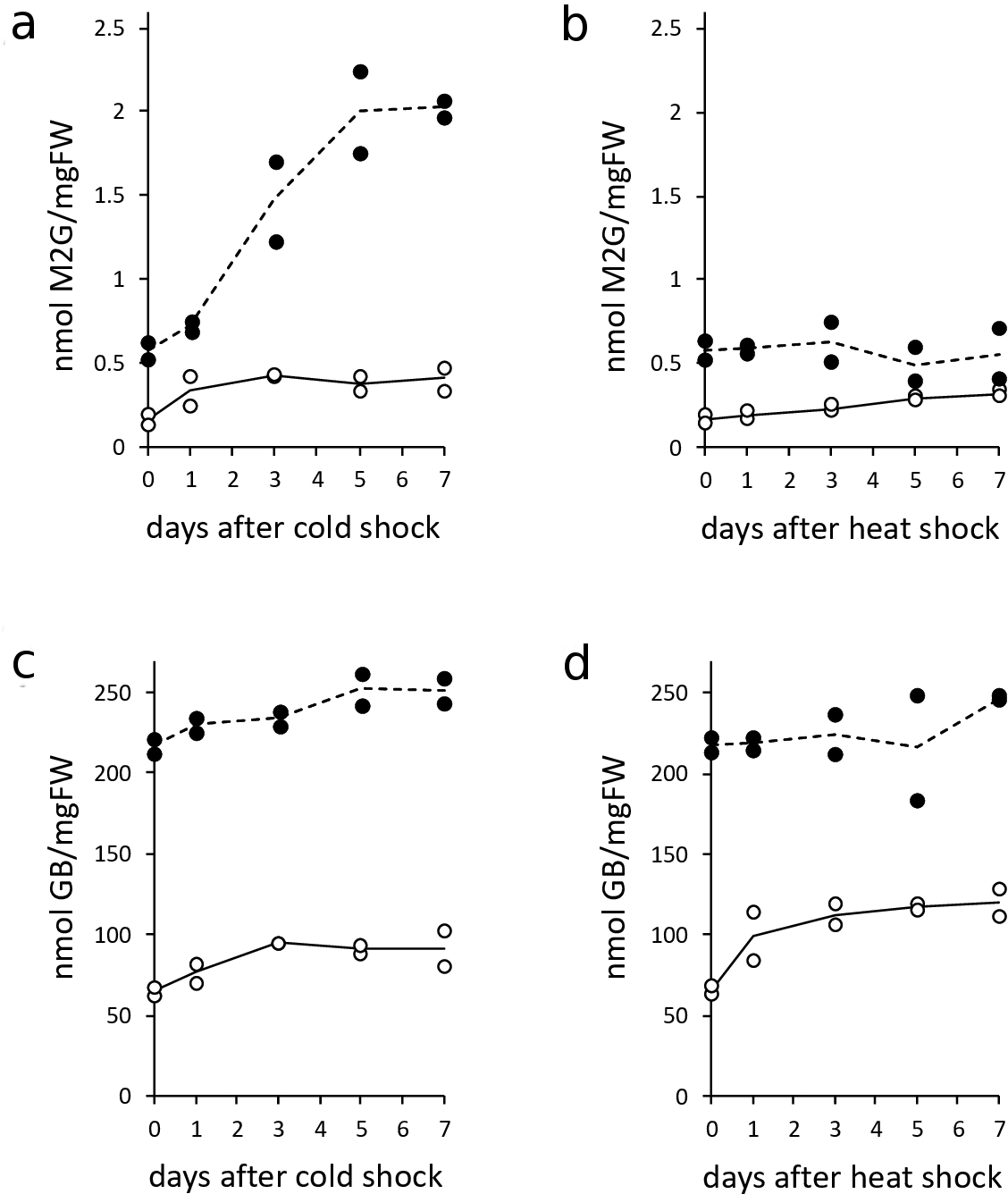
The accumulation of mycosporine-2-glycine in Halothece sp. PCC7418 cells grown under cold stress (a) or heat stress (b) conditions. The accumulation of glycine betaine in Halothece sp. PCC7418 cells grown under cold stress (c) or heat stress (d) conditions. Two independent experiments were performed for each condition, and the levels of these metabolites are indicated by circles. The concentration of NaCl supplemented into the culture media is indicated by open (0.5 M) and filled (2.5 M) circles. The solid and dashed lines show plots of average values.
In conclusion, the antioxidative power of aqueous extracts derived from Halothece sp. PCC7418 cells was enhanced by exposing the cells to temperature stress. We found that the optimal conditions for maximum induction of antioxidative power were a combination of cold shock and higher NaCl concentration. The quantification of constituent antioxidant molecules revealed that the amounts of phenolic compounds and PBPs were induced by temperature stress. In addition, cold stress enhanced the bioproduction of M2G, which suggested that M2G was partially responsible for the higher antioxidative activity of aqueous extracts derived from cold-shocked Halothece sp. PCC7418 cells. Since M2G has several potential therapeutic applications, such as antioxidative, 18,19 DNA protecting, 18 anti-inflammatory, 13 and antiglycation activity, 20 we have made efforts to develop the bioproduction process of M2G by utilizing Halothece sp. PCC7418 cells 19 and Escherichia coli cells. 17 Our optimized conditions for the production of M2G in Halothece sp. PCC7418 cells by cold stress may allow for the industrial production of M2G in the future because higher efficiency of accumulation of target compounds in microbial cells is a critical step.
Experimental
Chemicals/Reagents
Glycine betaine, gallic acid, and methanol were purchased from Wako (Osaka, Japan). Trolox and ABTS were obtained from Tokyo Chemical Industry (Tokyo, Japan). Folin-Ciocalteu reagent, aluminum chloride, and acetonitrile were purchased from Kanto Chemical (Tokyo, Japan). Potassium persulfate, 18-crown-6, and choline were purchased from Sigma-Aldrich (St Louis, MO, United States). Quercetin was obtained from Cayman Chemical Company (Ann Arbor, MI, United States).
Cyanobacterial Culture Conditions
Halothece sp. PCC7418 cells were grown photoautotrophically in liquid BG11 media containing the indicated NaCl concentrations and Turk Island salt solution under continuous light intensity of 70 µE m−2 s−1 at 30°C. The media were prepared as previously described. 12
Preparation of Aqueous Extracts From Halothece sp. PCC7418 Cells
Cyanobacterial cells were harvested by centrifugation at 2290 × g for 15 minutes at 4°C. The cell pellets were stored at −80°C before use. For preparation of aqueous extracts, the cells were resuspended in distilled water at a concentration of 50 mg fresh weight/mL (50 mgFW/mL). Thereafter, the cell suspensions were disrupted by sonication using a VP-5s sonicator (Taitec, Saitama, Japan). During the sonication, the cells were kept on ice and disrupted for 30 seconds with the output power set to 5. Next, the cells were further disrupted by sonication for 10 seconds with the output power set to 7. After centrifugation at 1000 × g for 5 minutes at 4°C, the supernatants were transferred into new tubes and subjected to further analyses.
2,2-Azino-Bis(3-Ethylbenzothiazoline-6-Sulfonic Acid) Diammonium Salt Assay
The ABTS assay was performed as described previously with minor modifications. 19 Briefly, the ABTS radical solution was generated by mixing 7 mM ABTS stock solution in water with 2.45 mM potassium persulfate in water. This mixture was then incubated in the dark for 16 hours at 25°C. Prior to the assay, the ABTS solution was diluted in water to generate an absorbance of approximately 0.5 at 415 nm. Test samples (10 µL) or trolox standards were added to 90 µL of diluted ABTS solution and mixed. After incubation for 15 minutes at 25°C, the absorbance at 415 nm was measured. Quantification was performed based on the trolox standard curve. The antioxidative power values were expressed in TEs µg/mgFW.
Total Phenolic Content Assay
The TPC was quantified by a previously described colorimetric method 21 with minor modifications. Briefly, 40 µL of each cell-free extract was mixed with 25 µL of distilled water and 105 µL of 10% (v/v) Folin-Ciocalteu reagent. After incubation at 25°C for 3 minutes, 80 µL of 7.5% (w/v) sodium carbonate was added and the mixture was incubated at 25°C for 15 minutes. Next, the mixtures were diluted with an equal volume (250 µL) of distilled water, and the absorbance at 760 nm was measured. Quantification was performed based upon the gallic acid standard curve. The TPC values were expressed in GAEs µg/mgFW.
Total Flavonoid Content Assay
The TFC was quantified by a previously described colorimetric method 22 with minor modifications. Briefly, 25 µL of each cell-free extract was mixed with 7.5 µL of 5% (w/v) sodium nitrate and 100 µL of water. After incubation at 25°C for 5 minutes, 7.5 µL of 10% (w/v) aluminum chloride was added and the mixture was incubated at 25°C for 5 minutes. Next, 100 µL of 4% (w/v) sodium hydroxide and 10 µL of water were added and the mixture was incubated at 25°C for 15 minutes. The absorbance of the mixtures was measured at 510 nm. Quantification was performed based upon the quercetin standard curve. The TFC values were expressed in QEs µg/mgFW.
Quantification of PBPs
Absorption spectra of Halothece sp. PCC7418 aqueous extracts were measured with a BioSpec-nano spectrophotometer (Shimadzu, Kyoto, Japan). Quantities of PE, PC, and APC were calculated from the absorbance at 562, 615, and 652 nm, respectively, using the following equations 23,24 :


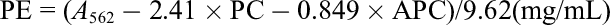
Extraction of M2G and GB
The extraction of M2G and GB was performed according to a previously described protocol with minor modifications. 10 Briefly, cyanobacterial cells were harvested by centrifugation at 2290 × g for 15 minutes at 4°C. Next, 10 to 20 mgFW of Halothece sp. PCC7418 cells were resuspended in 300 µL of methanol. Thereafter, the cells were sonicated for 10 seconds using the VP-5s sonicator with the output power set to 5. After centrifugation at 22 000 × g for 10 minutes at 4°C, supernatants were collected. After the addition of 200 µL of methanol to the remaining cell debris, sonication was conducted again for 10 seconds with the output power set to 5. The samples were then centrifuged as described above. The second supernatant was mixed with the first supernatant. Next, the methanol extracts were desiccated and redissolved in 200 µL of distilled water. After centrifugation at 22 000 × g for 10 minutes at 4°C, the supernatants were passed through Amicon Ultra Ultracel 3K centrifugal filters (Merck Millipore, Darmstadt, Germany). The resulting flow-through fractions were used to quantify M2G and GB.
Quantification of M2G
For quantification of M2G, high-performance liquid chromatography (HPLC) analysis was conducted as previously reported and the M2G authentic standard was prepared as described previously. 19
Quantification of GB
For detection of GB by HPLC analysis, GB was derivatized with p-bromophenacyl bromide and crown ether was utilized as the catalyst. GB extracts (50 µL) were mixed with 100 mM KH2PO4. Next, 900 µL of derivatizing solution (containing 66 mg of 18-Crown-6 and 1390 mg of 4-bromophenacyl bromide dissolved in 100 mL of acetonitrile) was added to the samples. The samples were then mixed and heated at 80°C for 60 minutes followed by centrifugation at 1000 × g for 5 minutes at 25°C. The supernatants (10 µL) containing the GB phenacyl esters were directly subjected to HPLC analysis. The HPLC analysis was conducted according to a previous report. 25 Briefly, a Supelcosil LC-SCX column (5 µm, 25 cm × 4.6 mm; Supelco, Bellefonte, PA, United States) was used for separation at 40°C. The mobile phase was run at a flow rate of 1.5 mL/min using 22 mM choline in 900 mL/L acetonitrile and 100 mL/L water. The derivatized GB was detected using a UV-visible detector at 254 nm.
Footnotes
Acknowledgments
The authors acknowledge financial support for this work generously provided by the Cosmetology Research Foundation and Hoyu Science Foundation.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was financially supported by the Cosmetology Research Foundation and Hoyu Science Foundation.