
Abstract
Lactuca indica L. has been traditionally used as a wild vegetable and as a medicinal plant for centuries. The various compounds present in it and their biological activities have been extensively reported. Hairy-root culture combined with agrobacterium-meditated metabolic engineering is a useful technique to achieve stable production of biologically active plant compounds. Here, we evaluated the enhancement of secondary metabolites in L. indica L. and their bioactivities by testing culture media composition and the use of an elicitor. Hairy roots were induced and cultured in MS or SH liquid media for 2 weeks prior to treatment with various concentrations of MeJa, for different periods. The resulting phenolic contents and physiological activities were analyzed. Higher total phenolic, flavonoid, and hydroxycinnamic acids contents were attained by elicitation with MeJa. Metabolite accumulation, especially in SH media and in the presence of MeJa, was time dependent. Particularly, accumulation of chicoric acid increased markedly with time. Similarly, we observed time dependent positive and negative responses of antioxidant activity in DPPH and ABTS assays, respectively. As in previous studies, the highest correlation was found between total phenolic content and total flavonoid content. Further, 3,5-DCQA showed the highest correlation with total phenolic content, total flavonoid content, and antioxidant activities in hydroxycinnamic acids. Our data effectively identified optimal culture conditions to increase the accumulation of secondary metabolites and antioxidant activity in hairy roots cultures of L. indica L.
Lactuca indica L. belongs to the family Compositae and is widely distributed in Asian countries, including Korea. It has traditionally been used as a wild vegetable and medicinal plant as it contains various biologically active compounds. L. indica L. contains triterpene acetate, quinic acid derivatives, and flavonoids, which reportedly show antioxidant activity and reduce serum levels of total cholesterol, and low-density lipoprotein-cholesterol; additionally, they show hepatoprotective and anti-mutagenic activity and stimulation of differentiation and epithelial cell protection from Escherichia coli infection. 1 –4
Traditionally, plants have been cultivated to produce not only food but also useful and stable secondary metabolites. Open-field cultivation is still the prevalent plant production system everywhere, and many cultivars of medicinal plants have been cultivated using this system, whereby yield and quality are dependent on uncontrolled environmental conditions such as temperature, solar radiation, and photo period. 5 Therefore, changes in the content of bioactive substances during the plant life cycle due to diverse abiotic factors affect quality control in the cosmetics, functional food, and pharmaceutical industries that use plant secondary metabolites. The interactions between plants and abiotic conditions described above have been observed in the previous studies in which L. indica L. was grown in the open environment. 6 Thus, complete control over all abiotic factors of the plant culture environment is required to attain high production of plant bioactive compounds with stable quality characteristics.
Hairy roots in plants which result from infection caused by Agrobacterium rhizogenes are produced in complete controlled environment. 7 Hairy roots have been investigated as an attractive system for producing valuable secondary metabolites from plants because of their characteristic high growth rate, genetic and biochemical stability, hormone independence, similar biosynthetic potential to parent plant, and suitability for scaling up for mass production. 7,8 Moreover, through in vitro culture, we can design a strategic approach, such as media formulation and elicitation, 9 and control the abiotic environment to specifically enhance the accumulation of secondary metabolites. Methyl jasmonate (MeJa), a volatile methylester of jasmonic acid, is an extensively used effective elicitor due to its outstanding role in signal transduction pathways. Indeed, various studies have demonstrated enhanced phytochemical contents in plants and hairy roots by MeJa treatment. 10
To the best of our knowledge, this is the first report on induced hairy roots from L. indica L. subjected to varying culture conditions regarding culture medium, elicitor concentration, and treatment duration with the aim to define the optimum conditions for promoting the accumulation of bio-antioxidant activities as natural products for use as raw material in health- and food-related industries.
Tissues from cut stems and leaves of L. indica L. were cultured in hormone-free Murashige-Skoog (MS) medium. Hairy roots were induced in the leaves after 1 week of culture (Figure 1a,b). Thereafter, they were subcultured once every 2 weeks in cefotaxime-treated MS solid medium for 2 months (Figure 1c). Samples (100 mg) of fast growing hairy roots were selected and cultured in 30 mL of MS or Schenk-Hildebrandt (SH) liquid media (Figure 1d). rol genes of the Ri-plasmid in A. rhizogenes are responsible for the induction of hairy roots from plant species. To determine whether these genes had successfully been inserted, we performed a polymerase chain reaction (PCR) using primers that the hairy root showed the expected bands for rol A (304 bp), B (797 bp), and C (550 bp) genes (Figure 1e).

Hairy roots of Lactuca indica L. obtained after infection by Agrobacterium rhizogenes and HPLC chromatograms of methanol extracts. (a) Explant of cotyledon leaves after inoculation with Agrobacterium rhizogenes R1000; (b) hairy roots 30 days after induction in leaves; (c) hairy root culture after 60 days growing on MS-agar medium; (d) hairy root culture in liquid culture medium; (e) results of PCR analysis to detect the rol A, rol B, and rol C genes in hairy root lines; and (f) HPLC chromatograms of methanol extracts from freeze-dried Lactuca indica L. leaves, roots, and hairy roots.
High-performance liquid chromatography (HPLC) chromatograms of the root extracts showed differences in hydroxycinnamic acids content between leaves and roots of the wild-type plant. Chlorogenic and chicoric acids were higher in the leaves, whereas 3,5-dicaffeoylquinic acid (DCQA) was more abundant in the roots. In turn, induced hairy roots showed a similar fluctuation pattern in hydroxycinnamic acids content to that described for the wild-type plant; furthermore, 3,5-DCQA was higher in the induced hairy roots than in roots of the wild-type plant (Figure 1f).
Similar responses were observed for total phenolic content (TPC) and total flavonoid content (TFC) to MeJA elicitation, as they both increased by exposure of MeJa and typically showed higher contents in SH than MS medium (Figure 2a,b). Although TPC and TFC fluctuated with exposure time, they both increased with extended duration of exposure. However, this was not a dose-dependent response. Highest TPC and TFCs under cultivation in SH media and after 72 hours exposure to MeJa were 0.028 ± 0.00004 and 0.022 ± 0.00023 µg/mL, respectively; such contents were 236.6% and 348.0% higher than in untreated cultures, respectively.

Accumulation of total phenolic (a) and flavonoid (b) content in the hairy root cultures of Lactuca indica L. in different media and elicitation conditions using methyl jasmonate (time of exposure, 0-72 h and concentration, 0-300 ppm).
In this study, 3 types of hydroxycinnamic acids were analyzed, namely, chlorogenic acid, 3,5-DCQA, and chicoric acid. Chlorogenic acid accumulated to a larger extent upon MeJa elicitation treatment in SH than in MS medium, although not in a time- or dose-dependent manner. Highest accumulation under the shorter duration or lower concentration of the elicitation treatment yielded chlorogenic acid content 220.6% (exposure to MeJa for 1 hour), 217.8% (exposure to MeJa for 2 hours), and 222.4% (exposure to MeJa at 50 ppm) higher than in untreated cultures (P < 0.05, Figure 3a).
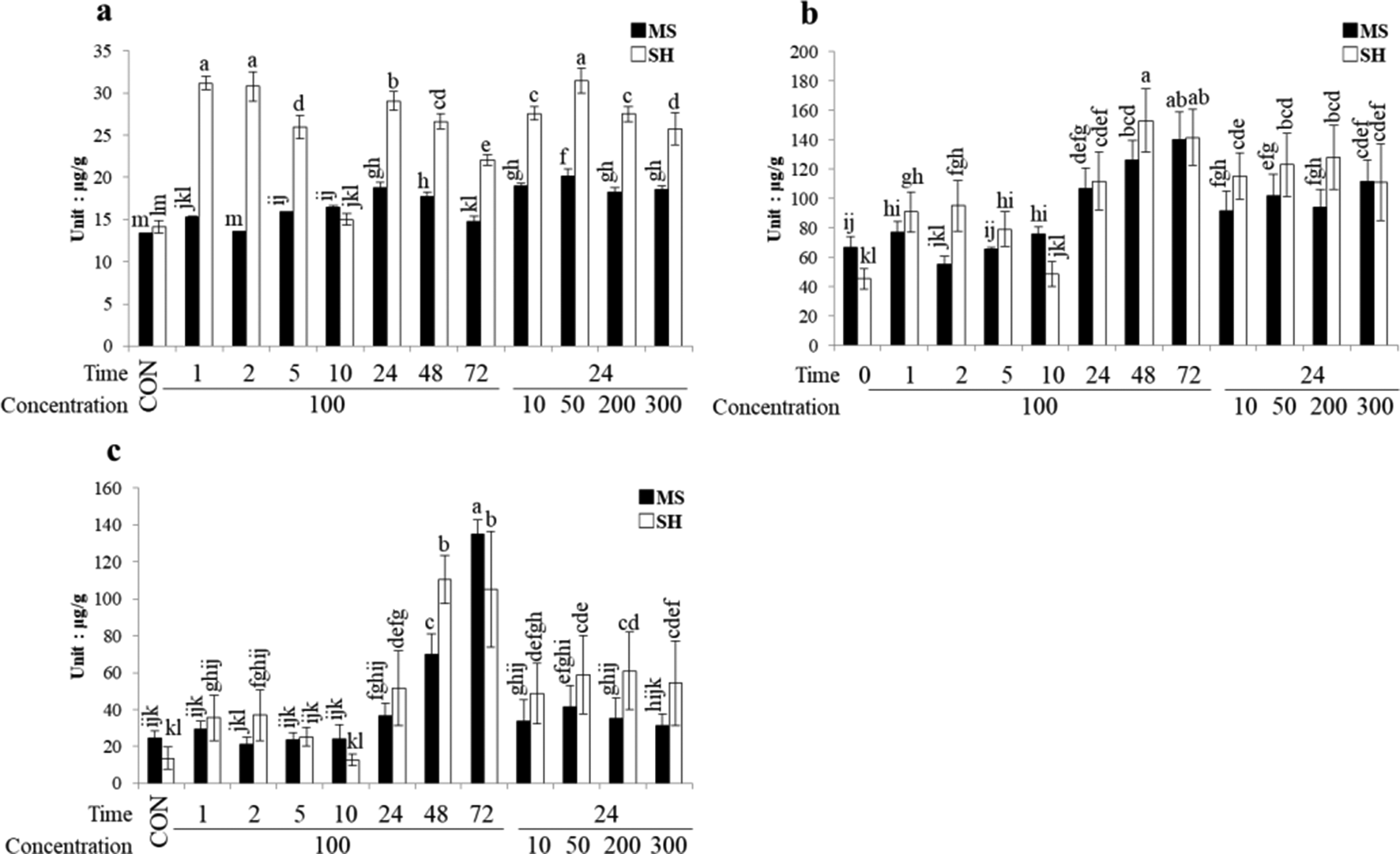
Accumulation of hydroxycinnamic acids in induced hairy root cultures of Lactuca indica L in different culture media and elicitation conditions using methyl jasmonate (time of exposure, 0-72 h and concentration, 0-300 ppm). (a) chlorogenic acid, (b) 3,5-DCQA, and (c) chicoric acid.
As for 3,5-DCQA and chicoric acid, neither TPC nor TFC differed between SH and MS media. In this case, the response was time dependent but not dose dependent. Maximum accumulation of 3,5-DCQA occurred after exposure to MeJa for 48 and 72 hours in SH medium or after exposure to MeJa for 72 hours in MS medium (P < 0.05, Figure 3b). The highest chicoric acid content was 553.4% greater than in untreated cultures and was obtained in cultures exposed to MeJa for 72 hours in MS medium (P < 0.05, Figure 3c).
Antioxidant activities present in induced L. indica L. hairy roots under the experimental culture conditions were assessed using the 2,2-diphenyl-1-picrylhydrazyl (DPPH) (Figure 4a) and 2,2ʹ-azino-bis-3-ethylbenzo-thiazoline-6-sulfonic acid (ABTS) (Figure 4b) assays. Higher antioxidant activities were measured by the DPPH assay in roots cultured in SH media and treated with MeJa for 48 (increase of 1.86-fold) and 72 hours (increase of 1.88-fold) (Figure 4a) than in untreated cultures. Conversely, a decrease in antioxidant activity was measured by the ABTS assay under the elicitation conditions tested, with inhibition rates ranging between 97.5% and 99.5% in both culture media (Figure 4b).

Antioxidant activities evaluated using 2,2-diphenyl-1-picrylhydrazyl (a) and 2,2ʹ-azino-bis-3-ethylbenzo-thiazoline-6-sulfonic acid (b) assays in the hairy root cultures of Lactuca indica L. in different culture media and elicitation conditions using methyl jasmonate (time of exposure, 0-72 h and concentration, 0-300 ppm).
Several significant relationships were observed between secondary metabolites, antioxidant activities, and treatment conditions (Table 1). The highest significant correlation was observed between TPC and TFC (r = 0.988, P < 0.01). Further, TPC and TFC were both significantly and positively correlated with 3,5-DCQA (r = 0.891 and 0.880, P < 0.01), chicoric acid (r = 0.865 and 0.870, P < 0.01), and DPPH scavenging activity (r = 0.942 and 0.933). In turn, MeJa treatment duration and 3,5-DCQA were closely associated with chicoric acid content. There was a significant positive correlation between treatment duration (r = 0.836, P < 0.01) and 3,5-DCQA (r = 0.875, P < 0.01). Conversely, ABTS scavenging activity showed a negative correlation with all other contents. In particular, significant negative correlations were detected between ABTS scavenging activity and TPC (r = −0.868, P < 0.01), TFC (r = −0.876, P < 0.01), and DPPH scavenging activity (r = −0.803, P < 0.01) (Table 1).
Correlation Analysis Among Phenolic Composition and Antioxidant Activity, Culture Medium, and Methyl Jasmonate Elicitation in Hairy Root Cultures of Lactuca Indica L.

ABTS, 2,2ʹ-azino-bis-3-ethylbenzo-thiazoline-6-sulfonic acid; DPPH, 2,2-diphenyl-1-picrylhydrazyl.
Significant correlation coefficient are shown in bold type (positive: red, negative: blue) based on Pearson’s correlation at *P < 0.05 and **P < 0.01.
Lactuca indica L. has been used as a wild vegetable and as a medicinal plant for centuries. Further, the demand for the species is currently on the rise owing to recent reports about its various biological activities presumably related to the effective promotion of human health. 1 However, the conventional field cultivated product cannot satisfy the industrial quality standards required, as crop yield and quality are strongly dependent on the environmental conditions prevailing during the plant growing season. Therefore, in the present study, we aimed to elucidate the optimum conditions to increase secondary metabolites and antioxidant activity contents in L. indica L. using hairy root cultures obtained under strictly controlled environmental conditions.
The ABTS and DPPH assays are widely used as they are 2 fast and easy methods to determine the antioxidant capacity of plant extracts, 11 and the results obtained from them are reportedly, strongly, and positively correlated. 12 Numerous reports have indicated that the ABTS assay is more effective in reflecting the antioxidant activity in various foods and is more strongly correlated with TFC than the DPPH assay. 13,14 In contrast, in the present study, we observed that ABTS results correlated negatively with DPPH results, as well as with other phenolic compounds and culture conditions, while its values were in very high and narrow range (97.5%-99.5%) of antioxidant activity under different culture conditions. Therefore, we concluded that the ATBS assay was not the best way of evaluating antioxidant activity due to a reduction of the ABTS or DPPH radical scavenging activity depending on a variety of polyphenols present in plant extracts 15 or to the likely involvement of other antioxidants that are polar or non-polar constituents other than phenolic compounds in ABTS radical scavenging activity. 2 Therefore, the use of more than 1 method for the evaluation of antioxidant activity in plant extracts seemed to be a more reliable approach. Accordingly, in the present study, DPPH assay was used as an alternative evaluation method of antioxidant activity in hairy root cultures of L. indica L.
Phenolic compounds are a group of secondary metabolites, including flavonoid and phenolic acids (eg, hydroxycinnamic acids), with high antioxidant potential, 16 and are the major contributors to the antioxidant activity in vegetables. 17 Strong correlations were found among TPC, TFC, and antioxidant activity (DPPH), as frequently reported in the previous studies. 16,18–21 Hydroxycinnamic acids have higher antioxidant capacity among phenolic acids such as hydroxybenzoic, hydroxyphenylacetic, and hydroxycinnamic acids. 22 Significant correlations between antioxidant activities and hydroxycinnamic acids were found in the present study. We believe that hydroxycinnamic acids contributed to antioxidant activity in the following ranking order: 3,5-DCQA > chicoric acid > chlorogenic acids, according to the same order of the correlation coefficients of hydroxycinnamic acids with DPPH, TPC, and TFC. This finding was in agreement with the observation that 3,5- DCQA showed stronger correlation with DPPH than with chlorogenic or chicoric acid, as reported by Kweon et al 23 and Ohnishi et al. 24 Therefore, our elicitation strategy proved an effective method which considered the contribution rate of target compounds to increased antioxidant activity in hairy root cultures.
An optimum culture medium is an important determinant of adventitious root productivity in terms of secondary metabolite production by in vitro culture systems. 25 However, it is difficult to predict the effect of the growth media as results will vary depending on the plant species. Therefore, the selection of the culture medium is important and needs to be carefully evaluated on a case by case basis to improve the production and quality of economic, plant-derived secondary metabolites, and antioxidant activity.
Elicitor concentration and time of exposure strongly influence secondary metabolite synthesis. 26 The previous studies reported similar results as the present study in that elicitation with MeJa significantly enhanced TPC and TFC in hairy root cultures of other plants. 11,27,28 Elicitation also enhanced the accumulation of hydroxycinnamic acids; particularly, MeJa significantly enhanced chicoric acid accumulation in L. indica L. Similar results were reported for Echinacea purpurea 29 and Echinacea angustifolia. 30 Furthermore, secondary metabolites were reportedly enhanced by increasing exposure duration in hairy root cultures of Salvia miltiorrhiza, 31 Salvia virgata Jacq., 27 and Rubia tinctorum. 32 In addition, an elicitor-concentration-dependent increase was observed in American ginseng 28 and in Solanum triobatum L., 10 whereas a combination of exposure duration and concentration was determinant in the case of S. virgata Jacq. 27
These results indicate that optimum culture and elicitation methods in hairy root cultures depend on plant species; accordingly, they should be determined systematically, based on empirical results. Regarding the commercial aspect, empirical optimization through a trial and error approach is expensive and time consuming, 33 although hairy root cultures are quite remarkable for their stability and reproducibility. Through machine learning, such as artificial neural networks or deep learning, it should be possible to reduce the time and the expense needed to optimize conditions for high yield and best quality of secondary metabolite accumulation in in vitro cultures. Other research groups have tried to use artificial neural networks for in vitro studies. 34,35 Future studies will have to consider that an optimum protocol for the whole process, from induction of the hairy root culture through the optimization of results, will be best determined using the machine learning approach to establish the best production conditions of highly functional materials.
In conclusion, culture medium and the presence of a strong elicitor of hairy roots positively influenced phenolic compound production and antioxidant activities by in vitro hairy root cultures. The optimum conditions to enhance these phytochemicals and bioactivities in hairy root cultures of L. indica L. were established. We expect that these results will increase the presence of plant natural products in the market through stable mass production of functional materials from L. indica L. using metabolic engineering for specific target compounds.
Experimental
Plant Material
Lactuca indica L. seeds were disinfected by immersion in 70% ethanol for 1 minute, followed by immersion in 2% sodium hypochlorite solution and gentle shaking for 10 minutes. Thereafter, the seeds were washed 3 times with sterilized water and 10-seed batches were cultured in petri dishes containing 25 mL of MS 36 solid medium. Culture conditions were maintained at 25°C under incandescent light for 16 hours.
Induction and Culture of Hairy Roots
Agrobacterium rhizogenes R1000 was used for the induction of hairy roots. Bacterial cultures were grown in Luria-Bertani liquid medium (1% [w/v] tryptone, 0.5% [w/v] yeast extract, 1% [w/v] NaCl, pH 7.0) for 16 hours in the dark at 28°C in a rotary shaker at 180 rpm. Cultures were then centrifuged at 1500 rpm for 10 minutes and the collected bacteria were adjusted to a density of A 600 = 1.0 with liquid MS medium.
Stems and leaves of L. indica L. grown for 30 days were cut and cocultured in liquid MS medium diluted with A. rhizogenes R1000 for approximately 15 minutes. After removing bacteria from the sterilized filter paper, plant tissues were transferred to the solid medium and cocultured in the dark for 2 days. Thereafter, the tissues were washed 3 times with sterile water to remove bacteria and cultured in MS solid medium treated with 400 mg/L of antibiotic cefotaxime, which had not been treated with any hormone. Hairy roots were induced in the leaves 1 week after culturing; thereafter, they were subcultured once every 2 weeks in cefotaxime-treated solid MS medium for 2 months.
Two different media, namely, MS and SH, 37 were used to evaluate the optimal media for production of secondary metabolites by L. indica L. hairy root cultures. Fast growing hairy roots were selected and cultured in these 2 media. Hairy roots (100 mg) were cultured for 2 weeks in 125 mL Erlenmeyer flasks containing 30 mL of liquid MS or SH medium at 25°C in a 100 rpm rotary shaker.
Each clone of hairy roots was incubated continuously to obtain enough biomass for the further experiment and then 1 line selected through the detection of rol A, B, and C genes (rol A: Forward 5′- CATGTTTCAGAATGGAATTA-3′, Reverse 5′-AGCCACGTGCGTATTAATCC-3′; rol B: Forward 5′-TCACAATGGATCCCAAATTG-3′, Reverse 5′-TTCAAGTCGGCTTTAGGCTT-3′; and rol C: Forward 5′-ATGGCTGAAGACGACCTGTGT-3′, Reverse 5′-TTAGCCGATTGCAAACTTGCA-3′). Genomic DNA of hairy roots was extracted and PCR amplified included 95°C for 2 minutes, 30 cycles of 95°C for 30 seconds, 55°C for 45 seconds, 72°C for 1 minute, and a final extension of 10 minutes at 72°C. After the amplification, amplified PCR products were mixed with loading dye and electrophoresed on 1% agarose gels prepared in Tris/Borate/EDTA buffer. Gels were analyzed using gel documentation system for the determination of respective rol genes (rol A, B, and C) fragment sizes.
Elicitation of Hairy Roots With MeJa
Elicitor MeJa was dissolved initially in 100% ethanol at an optimal concentration. To investigate the effect of MeJa concentration, hairy roots were cultured in MeJa-treated medium at one of the following concentrations: 0, 10, 50, 100, 200, or 300 ppm for 24 hours. To examine the effects of elicitor treatment duration, hairy roots were treated with 100 µM MeJa under the same conditions as described above, for 0, 1, 3, 6, 12, 24, 48, or 72 hours. Elicitor-treated induced hairy roots were collected and frozen in sealed, clear polyethylene plastic bags at −80°C until use. Each treatment was performed in 3 flasks and the experiment was repeated 3 times.
HPLC Analysis
Hairy roots grown under each experimental condition described above were frozen in liquid nitrogen and then ground to a fine powder using a mortar and pestle. Ten milligrams of each sample was extracted with 1 mL of 50% ethanol for 24 hours at 30°C in a water bath. After centrifugation, the supernatant was filtered through a 0.45 µm syringe filter and used for physiological activity measurements and HPLC analysis.
To analyze phenolic compound for evaluating the optimal combination of elicitation conditions and culture medium, the extracts were separated on a C18 column (250 mm × 4.6 mm, particle size 5 µm; Shimadzu, Kyoto, Japan) using a Prominence HPLC system (Shimadzu, Kyoto, Japan) equipped with a diode array UV-vis detector. Solvent A was water (formic acid 0.1%) and solvent B was acetonitrile (formic acid 0.1%). The flow rate was 0.7 mL/min, injection volume was 10 µL, and column temperature was maintained at 40°C. Analytical wavelengths were 280 and 330 nm. Initially, solvent B started at 12%, increased to 30% at 20 minutes and to 80% at 50 minutes. All solvents were purchased from Daejung Chemicals (Siheung, Korea) and standards were purchased from Sigma-Aldrich (St Louis, MO, United States). Calibration curves were drawn using 5 concentrations between 10 and 200 µg/mL of standard material, and the area of each component was quantitatively analyzed using a calibration curve.
Physiological Activity Measurement
TPC was measured using the modified methods of Anesini, Ferraro, and Filip. 38 Briefly, Folin-Ciocalteu phenol reagent was converted to molybdenum blue as a result of the polyphenol compound reduction in the extract. Fifty microliters of Folin-Ciocalteu reagent was added to 100 µL of each extract and reacted for 3 minutes; next, 300 µL of 20% Na2CO3 solution was added. After 15 minutes, 1000 µL of distilled water was added, and the supernatant was transferred to a 96-well plate. The absorbance value was measured at 738 nm and a calibration curve (R² = 0.998) was prepared using gallic acid as a reference material. The result was substituted into the calibration curve and expressed as gallic acid equivalents mg/g.
Total flavonoid content was determined by a modification of the methods used by Re, Pellegrini, Proteggente, Pannala, Yang, and Rice-Evans. 39 Briefly, 100 µL of 10% aluminum nitrate and 100 µL of 1 M potassium acetate were added to 500 µL of each extract and reacted for 40 minutes. The reaction solution was transferred to a 96-well plate and measured at 405 nm. The calibration curve (R² = 0.9993) was prepared using quercetin, which is a standard substance, and the results were substituted into the calibration curve and expressed as quercetin equivalents mg/g.
2,2-Diphenyl-1-picrylhydrazyl radical scavenging activity was analyzed using a modification of the methods used by Re, Pellegrini, Proteggente, Pannala, Yang, and Rice-Evans. 39 Briefly, 100 µL of each sample extract was added to 100 µL of 0.15 mM DPPH solution (dissolved in 99.9% ethanol) and reacted for 30 minutes. Absorbance reduction was measured at 519 nm and ascorbic acid was used as a standard.
2,2ʹ-Azino-bis-3-ethylbenzo-thiazoline-6-sulfonic acid radical scavenging activity was analyzed by a modification of the methods used by Re et al. 39 Briefly, a 1:1 ratio of 7.4 mM ABTS and 2.6 mM potassium persulfate were mixed and left standing overnight in a dark room to form ABTS cations. Then, phosphate-buffered saline buffer was diluted to 0.7 ± 0.03 at 738 nm. After adding 190 µL of the diluted ABTS solution to 10 µL of each sample extract, the reaction was allowed to proceed for 10 minutes in the dark, and the change in absorbance was measured at 738 nm. Ascorbic acid was used as a standard.
Statistical Analysis
SPSS 18.0 software (SPSS, Chicago, IL, United States) was used for the statistical analysis. Analysis of variance was used for treatment group comparisons. Pearson correlation was determined for all pairs of medium, treatment duration and concentration, secondary metabolites, and antioxidative activity based on treatment mean values. Data in figures and tables are means ± SD.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported partially by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2017R1D1A1B03030685).