
Abstract
Melanin plays a role in determining human skin color of a person, and a large amount of melanin makes the skin color look darkened. The proper amount of melanin formation protects our skin from UV radiation, but excessive melanin production causes hyperpigmentation and leads to freckles, melasma, and lentigo. In this study, we investigated the inhibitory effect of hydroxyectoine on melanogenesis and its mechanism in B16F10 cells. Melanin content and cellular tyrosinase activity were determined. The expression of microphthalmia-associated transcription factor (MITF), and the activities of tyrosinase and other melanogenesis-related enzymes, such as tyrosinase-related protein 1 (TRP-1) and tyrosinase-related protein 2, were also examined. Hydroxyectoine treatment significantly inhibited melanin production and intracellular tyrosinase activity in a dose-dependent manner. Western blot analysis showed that hydroxyectoine also reduced the expressions of tyrosinase and TRP-1. In addition, hydroxyectoine significantly reduced the expression of MITF, a major regulator of melanin production, and inhibited the phosphorylation of p38, c-Jun N-terminal kinase, and activated the protein kinase B. The results demonstrated that hydroxyectoine inhibits the expression of MITF through the inhibition or activation of melanin-related signaling pathways and downregulates melanogenesis by inhibiting melanogenic enzyme expression and tyrosinase activity. Hydroxyectoine has potential value in functional cosmetics applications, such as whitening.
Keywords
Melanin refers to the black or brown pigment present in the tissues of skin or eyes of many animals. The primary role of melanin is to protect the skin by blocking excess ultraviolet (UV) radiation. Also, the quantity of melanin present determines the skin color. Melanin is produced in melanocytes and is transferred through the dendrites of melanocytes to specific epidermal cells (keratinocytes). In these epidermal cells, melanin forms a hat-like structure around the nucleus to protect genes from UV radiation, scavenge free radicals, and protect cellular proteins. However, excess production of melanin can lead to skin diseases that include melisma, freckles, lentigo, age spots, and other hyperpigmentation syndrome. 1,2
In melanocytes, tyrosinase oxidizes the amino acid tyrosine as a substrate to form the dark brown pigment termed eumelanin. Tyrosinase is the key enzyme in the first 2 steps of melanin biosynthesis in which l-tyrosine is hydroxylated to the o-diphenol product l-3,4-dihydroxyphenylalanine (l-DOPA). l-3,4-Dihydroxyphenylalanine is further oxidized into the corresponding o-quinone. 3 The activation of microphthalmia-associated transcription factor (MITF) has been reported to contribute to melanogenesis by increasing the expression of melanin-related enzymes, such as tyrosine-related protein (TRP)-1 and -2 and tyrosinase. 4 -6 In addition, MITF is regulated by various signal pathways, which include the mitogen-activated protein kinase (MAPK), cyclic adenine monophosphate (cAMP), and phosphoinositide 3-kinase/protein kinase B (PI3K/AKT) signaling pathways. Recent studies have shown that the inhibition of p38 and c-Jun N-terminal kinase (JNK) phosphorylation in these MAPK pathways is associated with decreased melanin production. On the other hand, increased AKT phosphorylation in the AKT signaling pathway has been reported to inhibit melanin production. 7 -9
Recent studies have sought to develop tyrosinase inhibitors and biological reductants, such as kojic acid, arbutin, hydroquinone, and ascorbic acid, to treat hyperpigmentation and pigmented diseases. 10,11 However, tyrosinase inhibiting whitening agents have some side effects that include cytotoxicity, low stability to oxygen and water, and cutaneous irritation and dermatitis. These side effects limit their application. 12,13 Safer and more effective agents for skin whitening are a priority in cosmetics research and development.
Hydroxyectoine (Figure 1) is a (4S,5S)-5-hydroxy-2-methyl-1,4,5,6-tetrahydropyrimidine-4-carboxylic acid produced by high-density fermentation of salt-tolerant bacteria. It is now being produced on an industrial scale and has generated considerable biotechnological interest. 14 The compound is currently being used in a variety of ways, including as enhancers in polymerase chain reaction, protein stabilizers in vitro, and protein folding catalysts in vivo, whole cell stabilizers, and skin care products. 14,15
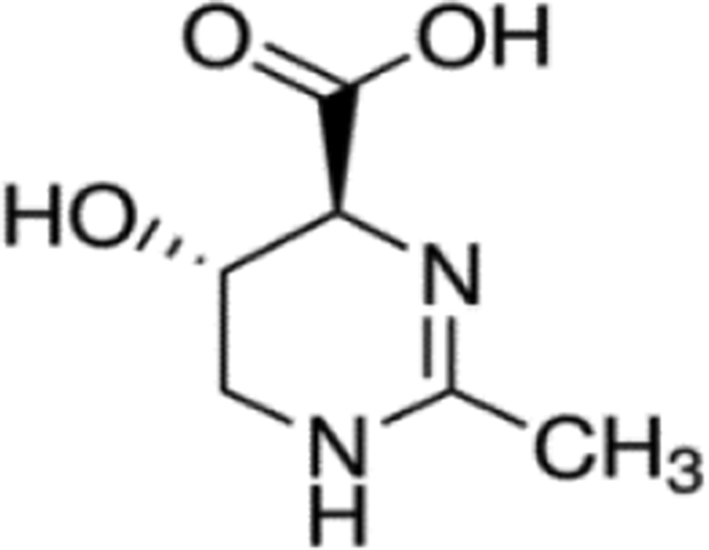
Chemical structure of hydroxyectoine.
Synthesis of hydroxyectoine depends on the prior production of ectoine. 16 Ecotoine is a natural organic compound found in halophilic organisms. Hydroxyectoine and ectoine are closely related chemically. Both function in compatible solutes. 17 -19 Cellular accumulation of these compounds through de novo synthesis or transport processes can protect cells from salt, heat stress, and dehydration. 20,21 However, the effects of hydroxyectoine and ectoine on biological macromolecules and cells may differ in several ways. Recently, ectoine has been reported to have a whitening effect. 22 Whether hydroxyectoine has a whitening effect is not known. This suggests that the response of hydroxyectoine involves inhibition rather than melanin production. Therefore, this study investigated the mechanism and the inhibitory effect of hydroxyectoine on melanogenesis through the inhibition of melanogenesis in B16F10 cells.
The relationship between concentrations of hydroxyectoine and the inhibition of melanogenesis in B16F10 cells and cell viability was investigated by the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. The assay is based on the ability of mitochondria in viable cells to reduce the aqueous soluble substrate of MTT tetrazolium to a purple, water-insoluble MTT formazan by dehydrogenase activity. The assay revealed that hydroxyectoine was not cytotoxic in B16F10 cells (Figure 2).

Influence of hydroxyectoine on viability of B16F10 cells. 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay was performed after incubation of B16F10 cell was treated with various concentrations (250, 500, and 1000 µM) of hydroxyectoine for 72 hours at 37°C in a 5% CO2 atmosphere.
Next, to investigate if hydroxyectoine inhibited melanin production, the melanin contents experiments were determined in B16F10 cells. Melanocytes are stimulated by stimulating factors that include UV radiation and alpha–melanocyte-stimulating hormone (α-MSH), which cause DNA photoproduction. 23 -25 We used α-MSH (200 nM) to stimulate melanogenesis in B16F10 cells. As shown in Figure 3, cells treated only with α-MSH displayed marked increases in melanin contents compared with cells treated with hydroxyectoine and arbutin. Hydroxyectoine inhibited α–MSH-induced melanin production in a dose-dependent fashion in B16F10 melanoma cells.

Hydroxyectoine inhibition of melanogenesis in B16F10 cells. The cells (2.0 × 104 cells/well) were pre-incubated for 18 hours and melanin content was determined after incubation of B16F10 cells treated with alpha–melanocyte-stimulating hormone (200 nM), arbutin (100 µM), or hydroxyectoine (250, 500, and 1000 µM) for 72 hours at 37°C in a 5% CO2 atmosphere. Absorbance was measured at 405 nm using an enzyme-linked immunosorbent assay P < 0.05 (*).
In melanosomes, tyrosinase acts as a catalyst in the early 2 stages of the melanin production mechanism. Overexpression of this enzyme leads to excessive production of melanin and pathological diseases, such as hyperpigmentation. 4 Inhibition of tyrosinase can reduce the content of melanin. The results of the determination of melanin contents indicated that melanogenesis is associated with the activity of tyrosinase involved in melanin synthesis.
Tyrosinase inhibitors that inhibit tyr osine-catalyzed oxidation and prevent pigmentation are becoming increasingly important in manufacture of pharmaceuticals and cosmetics. 26,27 Since melanin synthesis is regulated by tyrosinase, we examined intracellular tyrosinase activity in B16F10 cells to determine the tyrosinase inhibition of hydroxyectoine. As shown in Figure 4, hydroxyectoine decreased the intracellular tyrosinase activity in a dose-dependent manner. The findings indicated an association between decreased melanin contents and the inhibition of tyrosinase activity.

Hydroxyectoine inhibition of intracellular tyrosinase activity in B16F10 cells. The cells (1.0 × 105 cells/dish) were pre-incubated for 18 hours and then treated with alpha–melanocyte-stimulating hormone (200 nM) and hydroxyectoine (250, 500, and 1000 µM) for 72 hours at 37°C in a 5% CO2 atmosphere; P < 0.05 (*) or P < 0.01 (**).
Melanogenesis is controlled by an enzymatic cascade, which is regulated at the expression level of tyrosinase, TRP-1, TRP-2, and MITF. 28
Specifically, MITF is an important transcriptional regulator of tyrosinase and other melanogenic enzymes. 29 When present in greater amounts, α-MSH binds to its receptor, which increases the level of cAMP and subsequently activates cAMP-dependent protein kinase A (PKA). The activated PKA induces the phosphorylation of cAMP response element-binding protein and increases MITF protein levels. Microphthalmia-associated transcription factor induces melanin production by increasing the expression of tyrosinase and other melanogenic enzymes, such as TRP-1 and TRP-2. Therefore, reducing the expression of these enzymes reduces melanogenesis and promotes skin whitening. 30 We investigated how the expressions of the melanogenesis-related enzymes, MITF, tyrosinase, TRP-1, and TRP-2, are regulated by hydroxyectoine in B16F10 cells. As expected, hydroxyectoine significantly suppressed the expressions of MITF, tyrosinase, and TRP-1 at the transcriptional and translational levels (Figure 5). However, TRP-2 expression was not significantly changed. These data indicated that hydroxyectoine inhibits tyrosinase and TRP-1 expression by the downregulation of MITF protein expression.

Hydroxyectoine inhibition of tyrosinase-related protein 1, tyrosinase-related protein 2, tyrosinase, and microphthalmia-associated transcription factor proteins in B16F10 cells. The cells were stimulated with alpha–melanocyte-stimulating hormone (200 nM) in the presence of arbutin (100 µM) and hydroxyectoine (250, 500, and 1000 µM) for 24 hours. The protein levels were determined by immunoblotting.
Increasing phosphorylation of AKT in the PI3K/AKT signaling pathway decreases MITF expression via the inhibition of glycogen synthase kinase 3 beta, which downregulates melanogenesis. 31 To determine the effect of hydroxyectoine on AKT phosphorylation in B16F10 cells, western blot analysis was performed after treatment of cells with varying concentrations of hydroxyectoine. As shown in Figure 6, hydroxyectoine significantly increased the phosphorylation of AKT. The result indicated that hydroxyectoine can downregulate melanogenesis via the AKT signaling pathway.

Hydroxyectoine inhibition of the protein level of P-AKT in B16F10 cells. The cells (1.0 × 105 cells/dish) were pre-incubated for 18 hours and then stimulated with alpha–melanocyte-stimulating hormone (200 nM) in the presence of hydroxyectoine for 4 hours. Protein levels were determined using immunoblotting.
Recent studies linked the phosphorylation of p38 and JNK in MAPK signaling pathways to the upregulation of melanogenesis via increased MITF expression. 7,8 As shown in Figure 7, hydroxyectoine suppressed the phosphorylation of p38 and JNK, especially at the highest concentration (1000 µM). The results suggested that hydroxyectoine inhibits the expression of MITF by suppressing p38 and JNK phosphorylation, which downregulates melanogenesis.
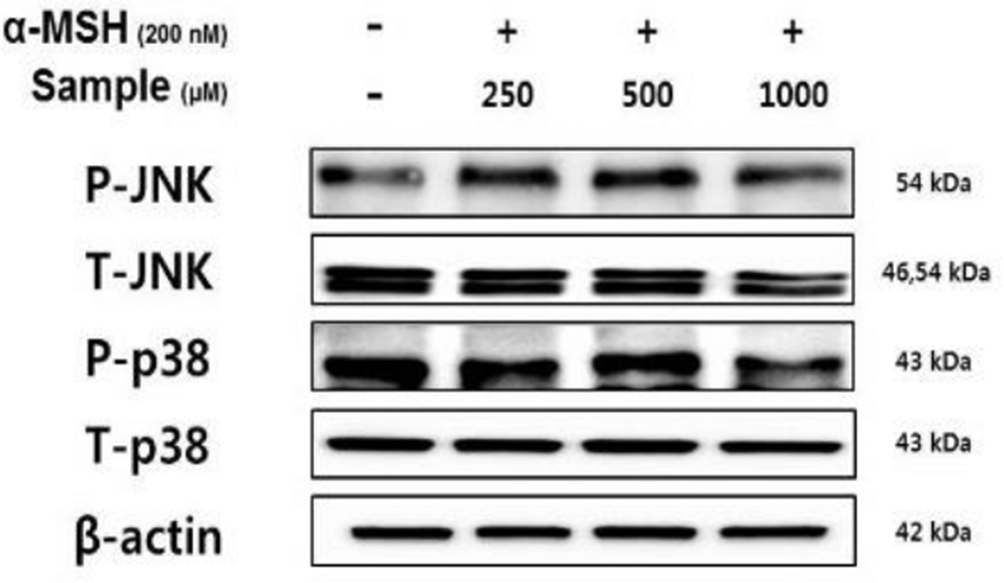
Hydroxyectoine inhibition of the protein levels of P-JNK and P-p38 in B16F10 cells. The cells were stimulated with alpha–melanocyte-stimulating hormone (200 nM) in the presence of hydroxyectoine for 4 hours. Protein levels were determined using immunoblotting.
In conclusion, hydroxyectoine can inhibit melanin synthesis and cellular tyrosinase activity in α–MSH-induced B16F10 cells, and also can suppress the expressions of MITF, tyrosinase, and TRP-1 expression. Furthermore, hydroxyectoine can decrease p38 and JNK phosphorylation and increase AKT phosphorylation. These results suggest that hydroxyectoine inhibits melanogenesis by inhibiting tyrosinase, TRP-1, and MITF expression through the JNK, p38, and AKT signaling pathways in α–MSH-induced B16F10 cells. The collective findings indicate the potential value of hydroxyectoine as a skin whitening agent.
Experimental
Cell Culture
The B16F10 murine melanoma cell line was purchased from the Korean Cell Line Bank (Seoul, Korea). Cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM) containing 10% fetal bovine serum, penicillin (100 units/mL), and streptomycin (100 µg/mL) at 37°C in a humidified atmosphere with an atmosphere of 5% CO2.
3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide Assay
Cell viability was measured by the MTT assays. B16F10 cells were cultured in 24-well plates for 18 hours and then treated with hydroxyectoine (250, 500, and 1000 µM) for 72 hours. 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide solution was added to each well for 4 hours. The solution was then removed and then the formazan crystals were dissolved in dimethyl sulfoxide. The absorbance was measured at 540 nm using an enzyme-linked immunosorbent assay (ELISA) reader.
Measurement of Melanin Content
The cells grown in DMEM were treated with hydroxyectoine and α-MSH for 72 hours at 37°C. Arbutin (100 µM) was used as the positive control. After removal of DMEM, the B16F10 cells were washed with cold 1× phosphate buffered saline (PBS). The cell pellet was resuspended in 1 N NaOH for 1 hour at 80°C to dissolve the cells. The absorbance of the released melanin was measured at 405 nm using an ELISA reader. Each experiment was performed in triplicate.
Intercellular Tyrosinase Activity
B16F10 cells (1.0 × 105 cells/dish) were seeded in a 60 mm dish for 18 hours, and then treated with various concentrations of hydroxyectoine and 200 nM α-MSH during a 72 hours incubation at 37°C in a humidified atmosphere of 5% CO2. The cells were recovered by centrifugation in an e-tube and each supernatant was removed. The cell pellets were lysed with phosphate buffer containing 1% Triton X-100. The lysates were centrifuged for 15 minutes at 13 000 rpm. After the quantification of protein levels and the adjustment of concentrations using lysis buffer, each lysate containing identical amounts of protein, and the test samples, the indicated concentrations were added to wells of 96-well plates, and 15 mM l-DOPA was added to each well. After incubation at 37°C for 1 hour, the absorbance was measured at 475 nm using an ELISA reader.
Western Blot Analysis
B16F10 cells (1.0 × 105 cells/dish) were incubated for 18 hours, and then treated for 24 hours with α-MSH (200 nM) and various concentrations of hydroxyectoine. The cells were washed twice with 1× PBS. The final cell pellet of each sample was collected in a microtube and the cells were lysed in a lysis buffer containing Radioimmunoprecipitation assay (RIPA) buffer, 1% Nonidet P-40, and 1% protease inhibitor cocktail for 1 hour. Each suspension was centrifuged at 15 000 rpm for 15 minutes at 4°C. Each supernatant was collected and added to a new microtube. The protein content of each cell lysate was adjusted to be the same based on the Bradford assay using a standard curve based on bovine serum albumin. After heating at 70°C for 10 minutes, an equal amount of the cell lysates was separated using 4% to 12% Bis-Tris mini gel electrophoresis (Invitrogen Inc.). The resolved proteins were transferred to nitrocellulose membranes (Invitrogen Inc.). Each membrane was washed using Tris-buffered saline (TBS; 20 mM Tris base, 137 mM NaCl, pH 7.6) containing 0.1% Tween 20 (TBST) and blocked in TBST containing 5% skim milk for 24 hours. Each membrane was incubated overnight at 4°C with the primary antibodies. The membranes were washed 6 times with TBST and then incubated with a secondary peroxidase-conjugated goat immunoglobulin G for 1 hour. The membranes were then washed 6 times with TBST. Chemiluminescence of the target protein was detected using an enhanced chemiluminescence (ECL) kit.
Data Analysis
All data are expressed as the means ± standard deviations of at least 3 independent experiments. Student’s t-test was used to determine statistically significant differences between each treated group and the negative control (α-MSH group). Statistical significance was considered at P < 0.05 (*) or P < 0.01 (**).
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by the Ministry of Trade, Industry & Energy (MOTIE), Korea Institute for Advancement of Technology (KIAT) through the Encouragement Program for The Industries of Economic Cooperation Region (P0006063 and R0002894).