
Abstract
Among the marine animals of the Pacific Ocean, crustaceans and mollusks are promising objects for the pharmaceutical industry. The composition of the total lipid fraction of these hydrobionts, includes poly unsaturated fatty acid and much less studied 1-O-alkyl-2,3-diacyl-sn-glycerols, compounds formed by fatty acids and fatty alcohols. The 1-O-alkyl-sn-glycerols (AG) were isolated from the digestive gland of the squid Berryteuthis magister. Previously, we proposed a simple procedure for obtaining AG with a purity of 99.22%, the main components of which are chimyl (94.39%) and batyl alcohol (4.35%). Purity and structure of the obtained products were confirmed by gas chromatography and mass spectrometry technique. In the present study, we showed that AG at concentrations of 0.1 to 5 µg/mL is a strong stimulator of lysosomal activity. In addition, AG increases the level of reactive oxygen species (ROS), the synthesis of nitric oxide (NO) and interleukin-6 expression in the RAW264.7 murine macrophage cell line. This study demonstrates the immunostimulating activity of AG in an in vitro experiment for the first time.
Marine lipids such as 1-O-alkyl-sn-glycerols (AG) and n-3 polyunsaturated fatty acids are the most well-known and abundant in natural sources. The digestive gland of some mollusk species contains a large amount of ether lipids, particularly 1-O-alkyl-2,3-diacyl-sn-glycerols (DAGE). The content of DAGE fraction is about 50% in the total amount of lipids. 1,2 Ether lipids differ in the length and the number of double bonds of the alkyl fragment in the sn-1 position, as well as differences in the fatty acids composition. C18:1 alkyl fragments (30.0% - 61.4%) are the most common in the DAGE lipids composition of shark liver oil, 3 while chimyl alcohol C16:0 (70.2%) is the main component of the squid Berryteuthis magister digestive gland lipids. 2 AG, the ethers of glycerol and fatty alcohols, are of interest to many researchers. 4
Most studies of AG biological activity have been performed in experiments with tumor therapy. It has been previously established that AG are able to activate macrophages and increase the synthesis of cytokines, such as IL-12 and IFN-γ. IL-12 promotes the secretion of IFN-gamma from naive and activated T- and NK-cells, increases the cytotoxic activity of NK-cells, cytotoxic T-lymphocytes, lymphokine-activated killers and increases the proliferation of T- and NK-cells. 5,6 AG also activates the cytotoxicity of macrophages, which leads to increased Fc-receptor-mediated phagocytosis and increased humoral immune response with a slower response of hypersensitivity. 7 In addition, AG are able to increase [Ca2+]i influx in human Jurkat T-cells presumably by modulating the permeability of calcium channels. 8 Some studies show that AG can be a new therapeutic compound to overcome the blood brain barrier (BBB) in the treatment of central nervous system pathologies. Based on the above data, it can be assumed that some examples of AG are very effective and low-toxic substances that can temporarily increase permeability of the BBB and increase the bioavailability of cytotoxic drugs for the brain. 9
However, despite the large number of studies on AG physiological activity, there are actually no data regarding the mechanisms of the immunostimulating effects of AG obtained from B. magister. This study provides a detailed description of AG immunostimulating activity in vitro.
Alkyl glycerols were derived from the digestive gland of squid B. magister. The main components of the original lipid composition were 1-O-alkyl-2,3-diacyl-sn-glycerols (38.50%) and triacylglycerols (24.26%). The first stage of separation was alkaline hydrolysis of lipid fraction to form a lipid mixture consisting of AG, free fatty acids, and cholesterol. The AG were separated by double recrystallization from acetone at -20°C and 1°C. Thus, a simple procedure has been proposed for obtaining AG with a purity of 99.22%, the main components of which are chimyl (94.39%) and batyl alcohols (4.35%). Purity and structure (Figure 1) of the products were confirmed by gas chromatography (GC) and GC-mass spectrometry (MS) technique. 10 The compounds obtained were tested as an immunostimulant on the RAW264.7 murine macrophage cell line. Johnson et al, presented an extensive toxicological characterization of chimyl and batyl alcohol, demonstrating that these compounds are safe for use in vivo in the concentrations studied. 11

Structures of the alkyl glycerols: 1 – chimyl alcohol; 2 – batyl alcohol.
In our study, in vitro cytotoxic activity of AG against cells RAW264.7 was measured using fluorescent probe, fluorescein diacetate (Figure 2a). Statistical analysis revealed that the EC50 values for cytotoxic activity of AG were 5 to 10 µg/mL. Figure 2(b) shows the morphology of macrophage RAW264.7 exposed to different concentrations of AG vs Escherichia coli lipopolysaccharide (LPS) used as positive control. When AG are added, macrophages show signs of moderate activation, which increases with increasing concentration from 1 to 10 µg/mL (cells become fusiform, some of them did not increase in size), whereas at 50 and 100 µg/mL all macrophages die. At a concentration of 1 µg/mL LPS causes excessive macrophage activation (cells swollen and vacuolated) which is even more enhanced at a concentration of 10 µg/mL.

(a) Cell viability, %. (b) Effect of various concentrations of AG and LPS on macrophage morphology.
During the inflammatory process, the formation of ROS occurs. Excessive formation of ROS increases inflammation, causes damage, and even necrosis of macrophages, which lead to uncontrolled release of cell contents into the extracellular medium and further enhances the inflammatory response. 12 In our study, AG in noncytotoxic doses of 0.1 to 5 µg/mL increased the formation of ROS in macrophage RAW264.7 by about 15% to 20% compared to the negative control (Figure 3a). At the same time, ROS production was not excessive, but only moderately activated the cells, since they maintained a normal morphological state and viability.
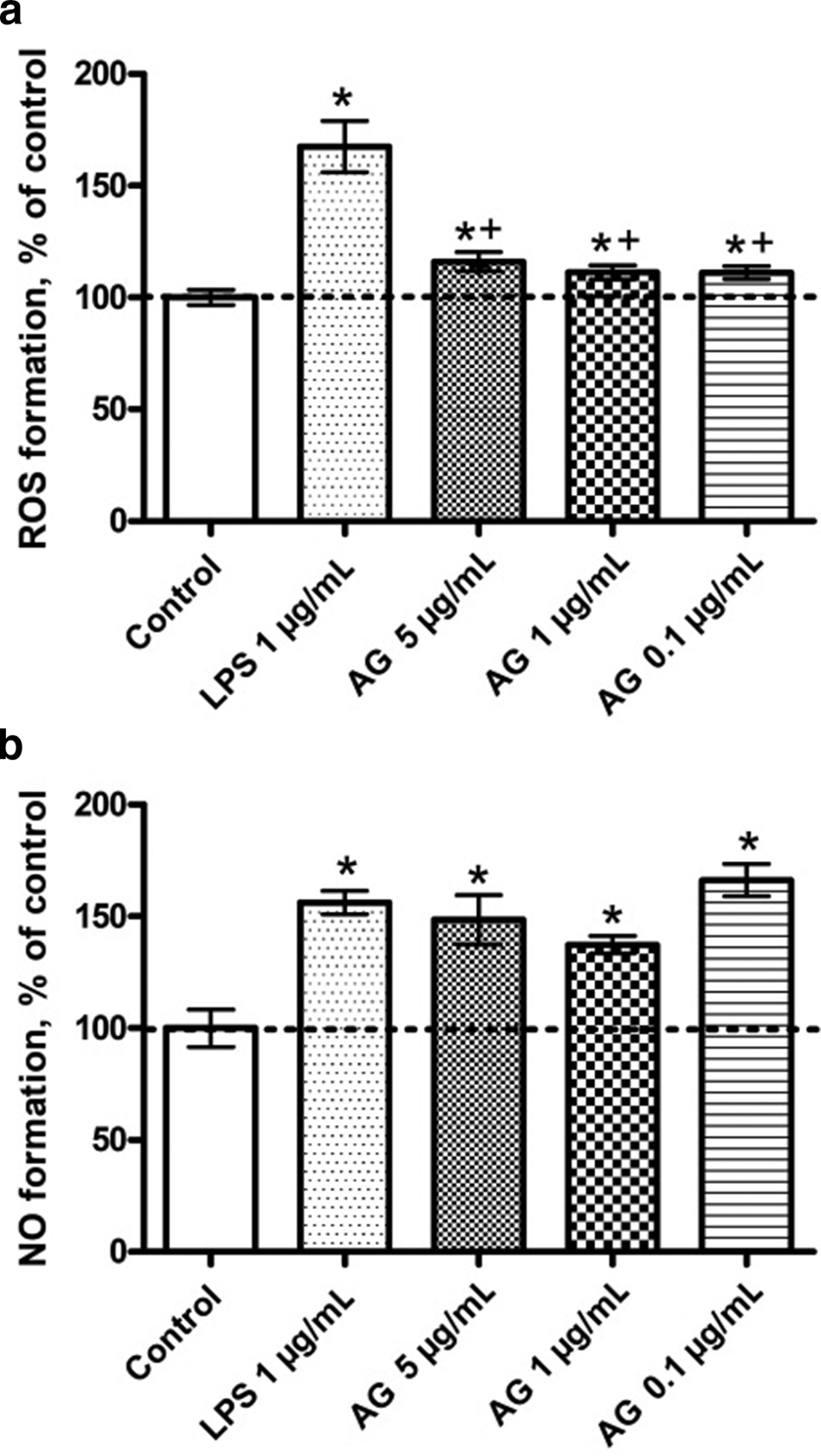
The quantitative analysis of intracellular ROS (a) and NO (b) formation in activated RAW264.7 cells after exposure to AG and LPS. *Significant differences between control and LPS, AG. +Significant differences between LPS and AG. Data are presented as mean ± SEM, P < 0.05, n = 3 (one-way ANOVA).
NO is considered as a mediator of inflammation and immunostimulation in various physiological states of mammals. 13 We found that AG induces an increase NO production in macrophages RAW264.7. The most effective concentration, which enhanced the NO formation in cells up to 60% to 70% compared to the control level, was 0.1 µg/mL. The concentration of 1 and 5 µg/mL increased the synthesis of NO to 40% to 50% (Figure 3b). At the same time, 5 µg/mL concentration of AG induced the synthesis of interleukin-6 in macrophages RAW264.7 compared to the control level (Figure 4a). It was previously shown that activation of NF-κB increase synthesis of TNF-α, interleukin-6 (IL-6) and transcription of the inducible form of NO-synthase, which is a key enzyme involved in the formation of NO. 14 Therefore, it can be assumed that AG enhances the synthesis of IL-6 and NO by direct or indirect activation of the transcription factor NF-κB.

(a) IL6 concentration in the RAW264.7 murine macrophages after exposure to AG and LPS. (b) Intracellular lysosomal activity in the RAW264.7 murine macrophages after exposure to AG and LPS. *Significant differences between control and LPS, AG. +Significant differences between LPS and AG. Data are presented as mean ± SEM, P < 0.05, n = 3 (one-way ANOVA).
It was previously established that AG administration in mice activates Fc receptors on macrophages, resulting in enhancement of phagocytic and cytotoxic activity. 15 In our study, the ability of AG to induce lysosomal activity was first demonstrated. The concentration 5 µg/mL significantly stimulated the lysosomal activity of RAW264.7 cells, by about 15% to 20% compared to the negative control (Figure 4b). Recent studies indicate that the role of lysosomes is not only in the processes of degradation and recycling of cellular waste, but also secretion, restoration of the cell membrane, signaling, and energy metabolism. In addition, the essential role of lysosomes in the autophagic pathway indicates the participation of these organelles in various cellular processes, which is of great importance in the development of various pathophysiological conditions. 16
LPS from E. coli is an endotoxin and causes a strong response from the mammalian immune system. It binds toll-like receptor 4 (TLR-4), activates inflammatory signaling pathways, promotes the secretion of pro-inflammatory cytokines, NO, and eicosanoids in several types of immune cells, and finally causes the classic symptoms of inflammation. 17 In the present study, it was shown that LPS at a concentration of 1 µg/mL increased lysosomal activity, as well as ROS, NO, and interleukin-6 synthesis in RAW264.7 cells (Figures 2–4). In all studied processes, LPS causes excessive activation of macrophages compared to AG in noncytotoxic concentrations.
Thus, AG can be considered a powerful immunostimulant that has a complex effect on the activation of macrophages in a relatively large noncytotoxic concentration range. In previous studies, the low toxicity of AG in vivo experiments was established. 11 Given the data obtained earlier and the results of this experiment, AG are promising drugs for the treatment of various bacterial infections, cancer, and immunodeficiency states. Further study of AG pharmacological activity mechanisms will form the basis for immunostimulating and anticancer drug development.
Experimental
Preparation of the Alkyl-Glycerol
The squid B. magister liver was obtained from Nakhodka Active Marine Fishery Base (Russia) and stored at -20°C. Extraction of total lipids was performed according to Bligh and Dyer. 18 Saponification of lipids was carried out by conventional technique. The lipid mixture after hydrolysis and acidification was dissolved in acetone in a ratio of 1:5 at room temperature and incubated for 24 hours at -20°C. 10 Recrystallization of obtained sediment was performed for complete AG separation from other lipids. The AG content in resulting product was over 99% where chimyl alcohol was the main component – 94% (Table 1).
AG Composition in Final Preparation.

aIndicated by chain length and double bond of alkyl chain in AG.
b% to total components (mean value ± standard deviation [n = 5]).
Cells for Bioassay
The RAW264.7 murine macrophage cell line was obtained from the American Type Culture Collection. The cells were cultured in DMEM supplemented with 10% FBS at 37°C under a humidified 5% CO2 atmosphere in an incubator (MCO-18AIC, Sanyo).
Nonspecific Esterase Activity Assay
RAW264.7 murine macrophages suspensions (1 × 103 cells/well) were incubated with solution AG (0.78, 1.56, 3.12, 6.25, 12.5, 25, 50, and 100 µg/mL) in a CO2 incubator at 37°C for 24 hours. A stock solution of the probe fluorescein diacetate (FDA; Sigma) in DMSO (1 mg/mL) was prepared. After incubation of the cells with the test compound, 10 µL of FDA solution (50 µg/mL) was added to each well and the plate was incubated at 37°C for 15 minutes. Cells were washed with phosphate buffered saline (PBS), and fluorescence was measured with a Fluoroskan Ascent plate reader (Thermo Labsystems, Finland) at λex = 485 nm and λem = 518 nm. All experiments were repeated in triplicate. Cytotoxic activity was expressed as the percent of cell viability.
ROS Level Analysis
RAW264.7 murine macrophages suspensions (1 × 103 cells/well) were incubated with AG solution (0.1, 1, 5 µg/mL) for 1 hour. Cells incubated without AG, and with LPS (1 µg/mL), were used as negative and positive controls, respectively. To study ROS formation, 20 µL of 2,7-dichlorodihydrofluorescein diacetate solution (Molecular Probes, Eugene, OR, USA) was added to each well, such that the final concentration was 10 mM, and the microplate was incubated for an additional 10 minutes at 37°C. 19 The intensity of dichlorofluorescein fluorescence was measured with plate reader PHERAstar FS (BMG Labtech, Ortenberg, Germany) at λex = 485 nm, and λem = 518 nm. The data were processed by MARS Data Analysis v. 3.01R2 (BMG Labtech, Ortenberg, Germany). The results were presented as a percentage of negative control.
Measurement of NO Content
RAW264.7 murine macrophages were plated into 96-well microplates and incubated at 37°C with 5% CO2 for 2 hours. After adhesion, cells were incubated with AG solution (0.1, 1, 5 µg/mL) for 24 hours. For endogenously generated NO detection studies, the cells were coincubated with 10 µM DAF-FM diacetate (Thermo Fisher Scientific, USA) for 40 minutes at 37°C. Prior to fluorescence registration, the cells were washed 3 times with PBS and then bathed in 200 µL/well of PBS. Green fluorescence of cells was registered at λex = 460 nm and λem = 524 nm. In each experiment LPS (1 µg/mL) were used as a positive control. Fluorescent intensity was measured using plate reader PHERAstar FS (BMG Labtech GmbH, Ortenberg, Germany).
Enzyme-Linked Immunosorbent Assay
Enzyme-linked immunosorbent assay (ELISA) was used to quantify level of IL-6 in RAW264.7 murine macrophages. Cell were plated into 96-well microplates and incubated at 37°C with 5% CO2 for 24 hours. After adhesion, cells were incubated with AG solution (5 µg/mL) for 24 hours. Cells incubated without AG, and with LPS (1 µg/mL), were used as negative and positive controls, respectively. Mouse IL-6 ELISA (ab100713, Abcam) kit was used according to the manufacturer’s recommendations. Cells with culture medium were homogenized on ice in the extraction buffer recommended by the manufacturer (100 mM Tris, pH 7.4, 150 mM NaCl, 1 mM EGTA, 1 mM EDTA, 1% Triton X-100, 0.5% sodium deoxycholate) with 1 mg/mL of protease inhibitor cocktail (Complete, Sigma-Aldrich) and 0.01 mg/mL of phosphatase inhibitor cocktail (P5726, Sigma-Aldrich). The protein concentrations were determined using a BCA protein assay kit (Pierce, Rockford, IL). The absorbance at 450 nm was measured with an iMark Microplate Absorbance Reader (Bio-Rad).
Lysosomal Activity Assay
An estimation of intracellular lysosome activity was conducted by staining of lysosomes in live macrophages with a fluorescent dye, acridine orange, followed analysis. RAW264.7 murine macrophages were plated into 96-well microplates and incubated at 37°C with 5% CO2 for 24 hours. After adhesion, cells were incubated with AG (5 µg/mL) for another 24 hours. Lysosomal activity was estimated as described earlier. 20 For this purpose 20 µL of aqueous solution of acridine orange (AO) (Sigma-Aldrich, A6014) was added to each final concentration 1.0 µg/mL, and the microplate was incubated additionally 30 minutes at 37°C. Intensity of AO fluorescence was measured at λex/λem = 485/538 nm. Prior to fluorescence registration, the cells were washed 3 times with PBS and then bathed in 200 µL/well of PBS. In each experiment LPS (1 µg/mL) was used as a positive control. Fluorescent intensity was measured using a PHERAstar FS plate reader (BMG Labtech).
Statistical Analysis
Each experiment was performed independently at least 3 times. The data were subjected to statistical analysis using one-way ANOVA tests followed by a post hoc Tukey’s multiple comparison test. Data were shown as mean ± SEM and P < 0.05 was taken as statistically significant. All statistical tests were performed using the GraphPadPrism 4.00 software.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work is supported by the Russian Science Foundation under grant 17-74-20006.