
Abstract
A standardized extract of Asparagus officinalis stem (EAS) is prepared from the lower residual parts of the plant. It is known to have antioxidative and neuroprotective activities. The purpose of the present study was to investigate the effects of EAS on photoaging in the epidermal layer of the skin using cultured keratinocytes. Normal human epidermal keratinocytes were irradiated or unirradiated with UV-B (10 mJ/cm2) and incubated with EAS (0.5 or 2 mg/mL) or vehicle. After 3 or 13 hours, molecular examinations were performed, and after 24 or 48 hours, cell viabilities were determined by a CCK-8 assay. EAS treatment upregulated the mRNA level of heparin-binding EGF-like growth factor in cultured keratinocytes 13 hours after UV-B irradiation. On the other hand, EAS downregulated caspase-9 and endothelin-1 mRNA levels in UV-treated keratinocytes. EAS also increased cell viability on keratinocytes 24 hours after UV-B irradiation. These results suggested that EAS might accelerate the migration of keratinocytes and protect the cells against damage induced by UV-B irradiation. Furthermore, EAS might inhibit melanocyte proliferation and UV-induced hyperpigmentation.
Asparagus officinalis is a perennially growing vegetable native to the western coasts of Europe. It contains many bioactive substances, such as carotenoids, 1 steroidal saponins, 2 and flavonoids. 3 A standardized extract of Asparagus officinalis stem (EAS) is prepared from the lower residual parts of Asparagus. 4 Previous studies have reported several biological activities of EAS, such as neuroprotective effects, 5,6 modulation of sleep stages, 7 beneficial effects against psychological stress, 8 and anti-oxidative activity. 9
Human skin is composed of various types of cells, including keratinocytes, melanocytes, and fibroblasts. Most of the cells constituting the epidermis are keratinocytes, which transmit the external stimulus to the melanocytes in the basal layer of subepidermis. Several important proteins and mediators regulate the proliferation, growth, maintenance, survival, and functions of keratinocytes. Heparin-binding EGF-like growth factor (HB-EGF) is a predominant growth factor that accelerates keratinocyte migration during wound healing. 10 Cell cycle-related protein, Ki-67, is a proliferation marker of epidermal keratinocytes. 11 Heat shock protein 27 (HSP27), a member of the small HSP family, is expressed in different cell types in adults, such as breast, uterus, and skin. 12 Overexpression of HSP27 prevents apoptosis triggered by various stimuli. 13 Furthermore, HSP27 regulates proinflammatory mediator release in keratinocytes. 14
Skin can be adversely affected by exposure to UV radiation, which can cause pigmentation, inflammation, aging, and cancer. UV radiation in sunlight is generally divided into three regions: UV-C (wavelength below 280 nm), UV-B (280-320 nm), and UV-A (320-390 nm). The high-energy wavelengths, such as UV-C, are effectively absorbed by the ozone layer in the stratosphere and, thus, cannot reach the earth’s surface. 15 The amount of UV-B irradiation reaching the earth’s surface is determined only by the stratospheric ozone layer. 15,16 The emission of certain chlorofluorocarbon chemicals contributes to the depletion of the ozone layer. 17 The increase in UV radiation reaching the earth’s surface is of considerable interest because of its impact on biological systems. 15
UV-B irradiation causes premature skin aging, referred to as photoaging, and accelerates aging-related changes, such as dyspigmentation abnormalities (hypermelanosis and hypomelanosis). 18 Some studies suggest that UV-B exposure increases the production of reactive oxygen species (ROS) in the skin. 19,20 When a high volume of ROS overwhelms the antioxidant defense mechanisms, it causes oxidative photodamage of proteins and other macromolecules in the skin. ROS are critical mediators of the photoaging processes. 21 Human keratinocytes secrete interleukin (IL)-1α in response to UV-B irradiation, which, in turn, induces the secretion of endothelin-1 (ET-1) from keratinocytes. 22,23 ET-1 acts as an intrinsic melanogen and mitogen for human melanocytes 24,25 ; therefore, excess ET-1 secretion may cause hyperpigmentation of the skin.
A study demonstrated that EAS has the potential to prevent skin aging by attenuating the oxidative stress responses in dermal fibroblasts. 9 However, there is no study related to the effects of EAS on photoaging in the epidermal layer of the skin. The purpose of the present study was to examine whether EAS has beneficial effects on UV-B-irradiated human keratinocytes by measuring gene expression.
The absorbance at 450 nm of UV-B-irradiated or unirradiated keratinocytes, which were cultured with EAS (0.5 or 2.0 mg/mL) or vehicle for 24 or 48 hours, is presented in Figure 1. Because there was no significant difference in absorbance between unirradiated normal human epidermal keratinocytes (NHEK) with and without EAS, this study evaluated the ratio of UV-B-irradiated NHEK cell viabilities to unirradiated NHEK cell viabilities. Twenty-four and 48 hours after the addition of EAS to keratinocyte medium, the absorbance ratio increased in an EAS concentration-dependent manner. Twenty-four hours after the addition of EAS, there was a significant difference in the absorbance ratio between vehicle and 2.0 mg/mL EAS.

The effects of EAS on cell viabilities of UV-B-irradiated keratinocytes. *P < 0.05 compared to vehicle treated. EAS: extract of Asparagus officinalis stem.
As there was no significant difference in absorbance between UV-B-unirradiated NHEK with and without EAS, we did not analyze mRNA expression levels in UV-B-unirradiated NHEK.
HB-EGF mRNA expression in NHEK cultured with 2 mg/mL EAS was significantly higher than that of untreated cells (P < 0.001) or cells treated with 0.5 mg/mL EAS (P < 0.001) after 13 hours UV-B irradiation (Figure 2a). Ki-67 mRNA expression levels in UV-B-irradiated-NHEK cultured without EAS showed an increase depending on time (Figure 2b). Ki-67 mRNA expression level in NHEK treated with 2 mg/mL EAS was, however, lower than that of untreated NHEK (P < 0.05) at the 13hour time-point post-UV-B irradiation (Figure 2b).

Real-time polymerase chain reaction analysis. (a) HB-EGF and (b) Ki-67 in UV-B-irradiated NHEK cultured with and without EAS at 3 and 13 hours after UV-B irradiation. Relative mRNA expression is presented as normalized to GAPDH or 18S. *P < 0.05, ***P < 0.001 compared to vehicle-treated, ## P < 0.01 compared to EAS-treated at 0.5 mg/mL concentration. EAS: extract of Asparagus officinalis stem; GAPDH: glyceraldehyde 3-phosphate dehydrogenase; HB-EGF: heparin-binding EGF-like growth factor; NHEK: normal human epidermal keratinocytes.
We found that HSP27 mRNA expression levels in NHEK increased in a time-dependent manner after UV treatment (Figure 3a). No significant difference was seen in caspase-3 mRNA expression levels between NHEK cultured with and without EAS at both 3 and 13 hours time-point (Figure 3b). However, 13 hours after UV-B irradiation, caspase-9 mRNA expression level in NHEK cultured with 2 mg/mL EAS was significantly lower than that of untreated cells or 0.5 mg/mL EAS-treated cells (P < 0.05) (Figure 3c).
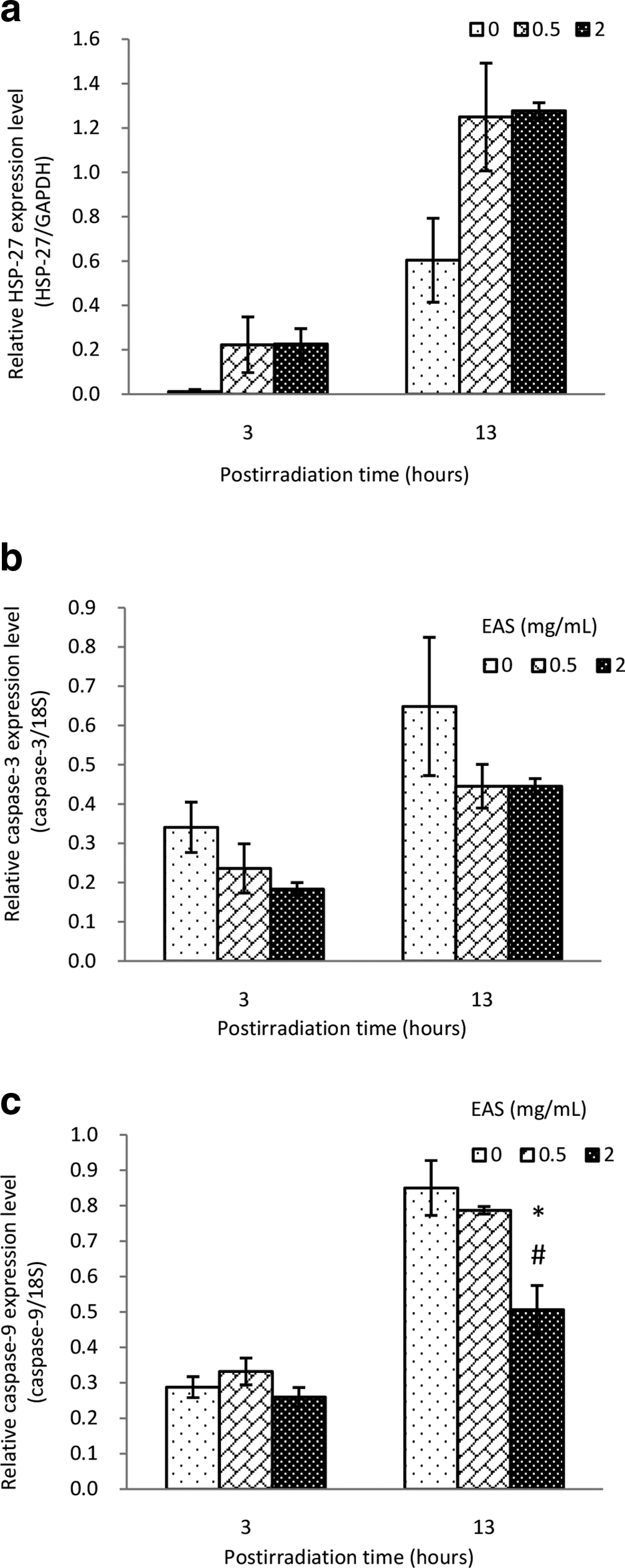
Real-time polymerase chain reaction analysis of HSP27 (a), caspase-3 (b), and caspase-9 (c) in UV-B-irradiated NHEK cultured with and without EAS at 3 and 13 hours after UV-B irradiation. Relative mRNA expression is presented as normalized to GAPDH or 18S. *P < 0.05 compared to vehicle treated, # P < 0.05 compared to EAS treated at 0.5 mg/mL concentration. EAS: extract of Asparagus officinalis stem; GAPDH: glyceraldehyde 3-phosphate dehydrogenase; HSP27: heat shock protein 27; NHEK: normal human epidermal keratinocytes.
No significant difference in mRNA expression levels of IL-1α were observed between NHEK cultured with and without EAS (Figure 4a). ET-1 mRNA expression level of EAS-treated NHEK was lower than that of the untreated cells at 13 hours after UV-B irradiation (P < 0.05) (Figure 4b).

Real-time polymerase chain reaction analysis of IL-1α (a) and ET-1 (b) in UV-B-irradiated NHEK cultured with and without EAS at 3 and 13 hours after UV-B irradiation. Relative mRNA expression is presented as normalized to 18S. *P < 0.05 compared to vehicle treated. EAS: extract of Asparagus officinalis stem; ET-1: endothelin-1; IL: interleukin; NHEK: normal human epidermal keratinocytes.
In the present study, EAS treatment has shown an increase in HB-EGF mRNA expression levels in NHEK, 13 hours post-UV-B irradiation. HB-EGF is a crucial autocrine migration accelerator for human keratinocytes. 10 In addition, we found that Ki-67 mRNA expression levels in UV-B-irradiated-NHEK cultured without EAS increased with time, and this increase was suppressed by EAS treatment at 13 hours after UV-B irradiation. Ki-67 is a cell proliferation-associated human nuclear antigen found in all cell cycle stages and it is a specific marker for multiplying keratinocytes in human epidermis. 11 Repeated UV irradiation has reported to induce keratinocyte proliferation with epithelial hyperplasia (acanthosis) or tumor. 26 Hence, these results suggested that EAS promoted UV-B-irradiated NHEK migration and might inhibit subsequent cell damages.
We observed that EAS significantly downregulated the mRNA expression of caspase-9 in NHEK, and marginally downregulated the mRNA expression of caspase-3, at 13 hours time-point after UV-B treatment. On the other hand, EAS increased the mRNA levels of HSP27. High doses of UV-B irradiation have been reported to induce apoptosis in human keratinocytes. 27 -29 UV irradiation predominantly activates the intrinsic apoptotic pathway, resulting in the activation of procaspase-9. 30 Caspase-9 initiates a caspase activation cascade, leading to procaspase-3 activation, which in turn, directly or indirectly, activates procaspase-8. 30 Caspase-3 activation leads to cleavage of multiple death substrates. 30 HSP27 blocks caspase-dependent apoptotic pathways. 13 In this study, EAS also increased cell viability on keratinocytes 24 hours after UV-B irradiation. Therefore, our results suggest that EAS suppressed the apoptotic pathway in UV-B-irradiated NHEK.
Keratinocytes in the skin were reported to secrete ET-1 in response to UV-B irradiation, accompanied by a significant secretion of IL-1α. 23 ET-1 produced and secreted by the keratinocytes plays an essential role in the maintenance of melanocyte proliferation and UV-induced hyperpigmentation in the epidermis. 23 Our study showed that EAS significantly downregulated the mRNA expression of ET-1 in NHEK, 13 hours after UV-B irradiation, suggesting that EAS may help prevent increased melanin pigmentation. In conclusion, we demonstrated that treatment with EAS significantly increased the mRNA expression levels of HB-EGF and decreased Ki-67 mRNA levels. Furthermore, caspase-9 mRNA expression was significantly decreased and this seemed to induce HSP27 mRNA levels. Thus, EAS may promote the migration of UV-B-irradiated NHEK and protect the cells against damage. Also, ET-1 mRNA expression levels significantly decreased with EAS treatment in UV-B-irradiated NHEK. Thus, EAS may have an inhibitory activity against melanocyte proliferation and UV hyperpigmentation in the epidermis.
Experimental
EAS
EAS (Amino Up Co., Ltd., Sapporo, Japan) is a standardized EAS. EAS contains 49.6% pinedex as a stabilizer.
Cell Viabilities
Normal human epidermal keratinocytes (NHEK) were obtained from Kurabo Industries Ltd. (Osaka, Japan) and cultured in serum-free medium, Humedia-KG2 (Kurabo Industries Ltd.), containing insulin (10 µg/mL), human epidermal growth factor (0.1 ng/mL), hydrocortisone (0.5 µg/mL), gentamicin (50 µg/mL), amphotericin B (50 ng/mL), and bovine brain pituitary extract (0.4%, v/v) under a humidified atmosphere of 5% CO2 at 37°C. For UV irradiation, the cells were plated on 24-well microplates, UV-B-irradiated (6 W, 9 seconds) at a dose of 10 mJ/cm2 using a commercial UV-B lamp (Analytic Jena, Jena, Germany), and incubated with EAS (0.5 or 2 mg/mL in dimethyl sulfoxide [DMSO]) or vehicle (pinedex 2 mg/mL in DMSO) for 24 or 48 hours. The dosage of 10 mJ/cm2 was used throughout this study, because keratinocyte growth was moderately inhibited by 8-12 mJ/cm2 and completely inhibited by 16-20 mJ/cm2. 24 The intensity of UV-B was quantitated in mJ/cm2 using an ultraviolet meter (Custom, Tokyo, Japan). CCK-8 reagent (80 µL/well) (Dojindo Molecular Technologies, Inc., Kumamoto, Japan) was added to keratinocytes medium, and the medium was incubated at 37°C for 1.5 hours. The absorbance at 450 nm of UV-B-irradiated or unirradiated keratinocytes was measured using a Benchmark Plus microplate spectrophotometer (Bio-Rad Laboratories, Richmond, CA, USA). The absorbance value was regarded as the relative cell viability. The absorbance ratio is a ratio to the unirradiated data.
Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
NHEK were cultured in serum-free keratinocyte medium and were irradiated with UV-B (10 mJ/cm2) and incubated with EAS (0.5 or 2 mg/mL) or vehicle for 3 or 13 hours to measure mRNA expression of HB-EGF, Ki-67, HSP27, caspase-3, caspase-9, IL-1α, and ET-1 using real-time PCR.
RNA Isolation
The total RNA was isolated from cultured NHEK using NucleoSpin RNA/Protein (Macherey-Nagel GmbH & Co. KG, Duren, Germany) and the concentration of total RNA was estimated using the Qubit 3.0 Fluorometer (Thermo Fisher Scientific, Waltham, MA, USA).
RT-PCR
For each cultured NHEK sample, 50 ng of total RNA was reverse-transcribed using PrimeScript RT Reagent Kit (Takara Bio Inc., Shiga, Japan) in a total reaction volume of 10 µL containing PrimeScript Buffer, PrimeScript RT Enzyme Mix I, and 50 pmol of random primer. The reaction was performed for 15 minutes at 37°C and stopped by heating for 5 seconds at 85°C. Polymerase chain reaction was performed with SsoAdvanced Universal SYBRGreen Supermix (Bio-Rad Laboratories, Hercules, CA, USA) in a total volume of 10 µL containing 4 µL of cDNA (diluted the cDNA solution 50 times with RNase free water) and primers. Primers for HB-EGF, Ki-67, HSP27, caspase-3, caspase-9, IL-1α, ET-1, 18S, and glyceraldehyde 3-phosphate dehydrogenase were obtained from Thermo Fisher Scientific (Waltham, MA, USA). The following PCR cycling conditions were used: 95°C for 30 seconds, 40 cycles at 95°C for 5 seconds, and 56–64°C for 10 seconds. The fluorescence emission from 96-well plate at each cycle was monitored using a CFX ConnectTM Real-Time PCR Detection System (Bio-Rad Laboratories, Hercules, CA, USA). The level of each target gene was calculated using standard curve method.
Data are expressed as the mean ± standard error of mean from 3 independent experiments (n = 3). Statistical significance of the differences between the EAS-treated and vehicle-treated control was determined by one-way analysis of variance followed by Tukey’s post hoc test. P values less than 0.05 were considered statistically significant.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for theresearch, authorship, and/or publication of this article: This study was supported in part by Amino Up Co., Ltd.