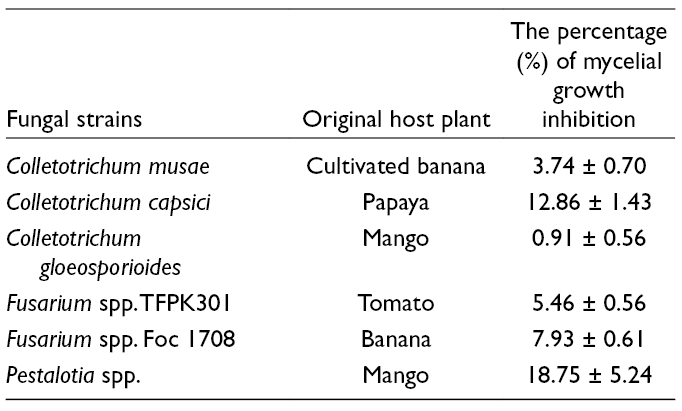
Cordytropolone (1) and (−)-leptosphaerone A (2) were isolated from the culture broth of the fungus Polycephalomyces nipponicus. The structures of these two compounds were elucidated by spectroscopic methods and from a comparison of the spectroscopic data with those reported previously. The structure of 1 was confirmed by X-ray crystallography for the first time while the leptosphaerone class (compound 2) was first isolated as its (+)-antipode from the fungus Polycephalomyces (Cordyceps). The fermentation process was monitored weekly by High performance liquid chromatography analysis for 10 weeks. The predominant compound (1) was produced at ~0.65 mg/mg of dry extract at week 9. Compound 1 exhibited modest antipathogenic fungal activity against Collectrichum musae, Colletotrichum capsici, Colletotrichum gloeosporioides, Fusarium spp. TFPK301, F. spp. FOC1708, and Pestalotia spp. with percentage of mycelial growth inhibition values of 3.74 ± 0.70%, 12.86 ± 1.43%, 0.91 ± 0.56%, 5.46 ± 0.56%, 7.93 ± 0.61%, and 18.75 ± 5.24%, respectively, at 25 μg/mL.
Cordyceps (family Cordycipitaceae) is an insect pathogenic fungus which comprises about 400 species worldwide. This genus has long been used in Chinese and Tibetan traditional medicine from ancient times. There are many reports indicating that the secondary metabolites produced by Cordyceps have interesting biological properties, including antioxidant,1 anti-inflammatory,2,3 antimalarial,4–7 kidney-protective,8 anticancer,9,10 anti-bacterial,11 and cytotoxic12,13 activities. Cordyceps nipponica, which has had a nomenclatural change to Polycephalomyces nipponicus, exhibits antibacterial11 and anticancer14 activities. The isolation of cordypyridones A–D from this species and their antimalarial and cytotoxic activities have been published.4 In our search for bioactive natural products from local fungi, cordytropolone (1) and a new natural product, (−)-leptosphaerone A (2), were isolated from the culture broth of P. nipponicus. The X-ray structure of 1 and the nuclear magnetic resonance (NMR) data of 2, the production of 1 and 2 from this fungus, and the antipathogenic fungal activity of 1 are reported herein.
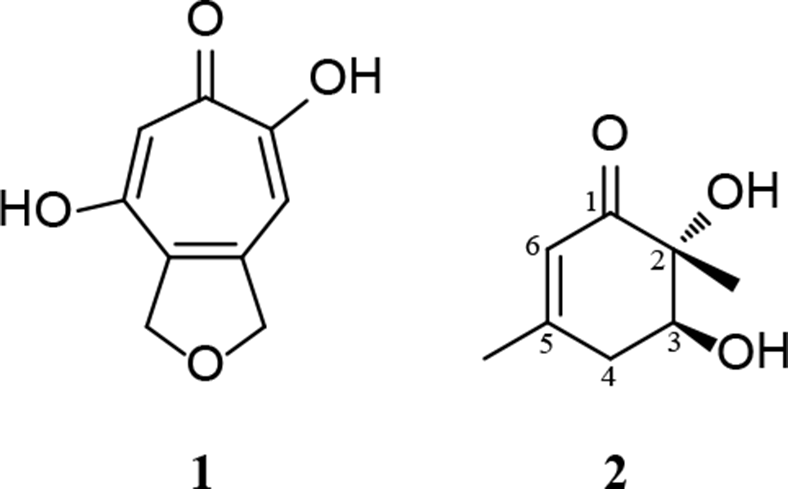
Compound 1, a colorless amorphous powder, was obtained from the dried broth extract from a large-scale fermentation by trituration in ethyl acetate. This compound was identified as cordytropolone (C9H8O4) (Figure 1). Its 1H- and 13C-NMR spectroscopic data in DMSO-d6 and MS were the same with those reported for cordytropolone from the fungus Cordyceps sp. BCC 1681.5 Crystallization of 1 from methanol:water successfully provided small crystals and the structure of 1 was confirmed by X-ray crystallography for the first time (Figure 2). Considering the high amount of 1 (90% yield from the dried extract) produced by P. nipponicus and previous isolation (69% yield from the dried extract) from Cordyceps sp. BCC 1681 these two fungi are considered to be good sources of bioactive cordytropolone.
Structures of 1 and 2.
X-ray structure of 1.
Compound 2, a pale brownish oil, has a molecular formula of C8H12O3 (m/z 179.07 [M + Na]+), as determined by ESIMS. The structure of 2 (Figure 1) was elucidated on the basis of its NMR spectroscopic data in MeOH-d4 (Table S1). This compound had the same structure as leptosphaerones A and B, which have been isolated from the fungus Leptosphaeria herpotrichoides.15 Comparison of the NMR spectroscopic data of 2 (Table S2) with those reported in the literature for leptosphaerone A were the same. However, their optical rotation values were different. Leptosphaerone A showed [α]D + 1.9 (c = 0.47, CHCl3), while 2 displayed a specific rotation of the opposite sign [α]D25 −1.7 (c = 0.49, CHCl3). This information indicated that 2 was the enantiomer of leptosphaerone A (2S, 3R). The lack of a NOESY correlation (in CDCl3, Figure S11) from the methyl protons 2-CH3, δH 1.26 (s), to the nearby oxygenated methine proton H-3, δH 4.00 (dd, J = 10.5, 6.0 Hz) supported the trans relationship between these protons. Hence, the stereogenic centers of 2 were supported as 2R, 3S. The name (−)-leptosphaerone A was given to this new compound.
The antimalarial (P. falciparum, K1) and cytotoxic (KB and BC-1 cell lines) activities of cordytropolone,5 and the cytotoxicity (A-549 cell lines) of leptosphaerone C (same identified structure with leptosphaerone A) isolated from the fungus Penicillium sp. have already been published.16
A time course of a small-scale fermentation of P. nipponicus for 10 weeks was conducted in order to study the production of 1 from the culture broth of this fungus. From our experiment, the weight of the dried mycelium varied from 0.91 to 1.56 g. It obviously increased from weeks 1 to 3, slightly went up from weeks 3 to 5, and then started to decrease from weeks 5 to 10 (Table 1).
Weight of Dried Mycelium and Extraction Yield of Culture Broth of the P. nipponicus From the Small Scale Fermentation.
Fermentation (week)
Weight of dried mycelium (g)
Extraction yield of culture broth(mg/100 mL of broth)
1
1.02
28.53
2
1.40
23.75
3
1.49
40.89
4
1.50
15.90
5
1.56
31.83
6
1.37
48.00
7
1.30
26.72
8
1.27
30.22
9
0.92
20.57
10
0.91
25.00
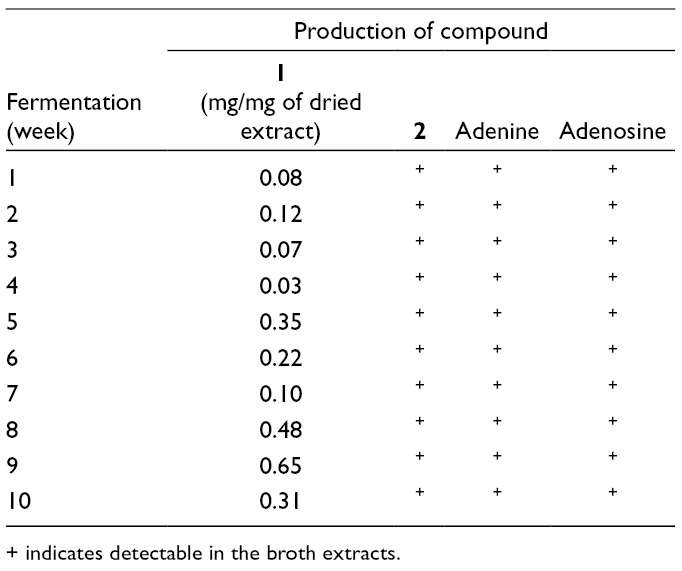
The production of 1 and 2 in the broth extracts was monitored by high performance liquid chromatography (HPLC) analysis using pure compounds 1 (tR 2.52 min) and 2 (tR 11.20 min), and also adenine (tR 7.63 min) and adenosine (tR 12.35 min) as reference standards (Figures S13-S16). This study showed that compounds 1 and 2 were produced by P. nipponicus in every week of the fermentation, as well as adenine and adenosine (Table 2). The quantity of 1 in the extracts was determined from its peak area calculated based on a standard linear equation of pure compound 1 (r2 0.988), which showed the highest at week 9 (~0.65 mg/mg of dried extract). Surprisingly, the production of 1 at week 3 (21 days), the optimal fermentation condition for growing this fungus on induced medium, was very low when compared with most other weeks. These results were different from our large-scale fermentation (20 days) results. Therefore, a second large-scale fermentation was started and the isolation of 1 under the same procedure was repeated. Only 756.2 mg (55% yield from 1.38 g of the dried extract) of 1 was obtained. These results confirmed that 1 is produced by this fungus, but the compound is not stable under the same fermentation conditions.
Production of 1, 2, Adenine and Adenosine From Small-Scale Fermentation of P. nipponicus.
Fermentation(week)
Production of compound
1(mg/mg of dried extract)
2
Adenine
Adenosine
1
0.08
+
+
+
2
0.12
+
+
+
3
0.07
+
+
+
4
0.03
+
+
+
5
0.35
+
+
+
6
0.22
+
+
+
7
0.10
+
+
+
8
0.48
+
+
+
9
0.65
+
+
+
10
0.31
+
+
+
+ indicates detectable in the broth extracts.
The antifungal activity of 1 at 25 µg/mL was tested against 6 plant pathogenic fungi. The results revealed that 1 had a slight inhibitory effect against fungal mycelial growth, as shown in Table 3. The antifungal activity of 1 against plant pathogenic fungi including Colletotrichum capsici and Colletotrichum gloeosporioides has been reported recently.17 However, the mechanisms of action against fungal pathogens have not yet been described.
the Percentage of PGI of 1 Against Six Plant Pathogenic Fungi.
Fungal strains
Original host plant
The percentage (%) of mycelial growth inhibition
Colletotrichum musae
Cultivated banana
3.74 ± 0.70
Colletotrichum capsici
Papaya
12.86 ± 1.43
Colletotrichum gloeosporioides
Mango
0.91 ± 0.56
Fusarium spp. TFPK301
Tomato
5.46 ± 0.56
Fusarium spp. Foc 1708
Banana
7.93 ± 0.61
Pestalotia spp.
Mango
18.75 ± 5.24
Compound 2 was tested for its cytotoxicity against human breast cancer (MCF-7), oral human epidermoid carcinoma cancer (KB) and Vero (African green monkey kidney fibroblasts) cell lines, and antivirus activity against Herpes simplex virus type-1 (HSV-1) at the National Center for Genetic Engineering and Biotechnology (BIOTEC), Thailand. It was found to be inactive in all these tests.
Cordytropolone (1) and the new natural product (−)-leptosphaerone A (2) were isolated from the culture broth of the fungus P. nipponicus. The structure of 1 was confirmed by X-ray crystallography for the first time. The NMR spectroscopic data of 2 and the isolation of the leptosphaerones from the fungus Polycephalomyces (Cordyceps) are reported for the first time. Compounds 1 and 2 were produced regularly during 10 weeks of fermentation, but the content of 1 was not stable. The antifungal activity of 1 was considered weak against 6 plant pathogenic fungi, while 2 was found to lack cytotoxicity against MCF-7 and KB cancer cell lines and was also inactive against HSV-1. Owing to the high quantity of 1 produced by this fungus, fermentation conditions that can lead to the stable production of 1 and structure modification of this compound on its biological activity are our interests.
Experimental
General
1H-NMR (500 MHz), 13C-NMR (125 MHz), and 2D NMR spectra were recorded in DMSO-d6 for 1, and MeOH-d4 and CDCl3 for 2 on a Bruker 500-MHz Avance Neo NMR Spectrometer. Optical rotations were determined using a JASCO DIP-1000 digital polarimeter. Mass spectra were obtained from a Bruker micrOTOF mass spectrometer. Column chromatography was carried out on Merck silica gel 60 and Pharmacia Fine chemicals Sephadex G-75. HPLC was performed on a Shimadzu system with a SLC-10AD controller and detector using a diode array detector (SPD-M20A; Shimadzu). A C18 column (250 × 4.6 mm, 5 µm) was used in this analysis.
Fungal Material and Fermentation
Polycephalomyces nipponicus (Cod-MK1201) was isolated from a dead cicada nymph and collected from Muang District, Maha Sarakham Province, northeast Thailand. The fungus was isolated and identified by one of the authors (A.S.). The culture used throughout the experiment was maintained on potato dextrose agar (PDA) slants at 28°C. For inoculum preparation, the fungus was initially grown at 28°C on a PDA plate for 14 days. The hyphal tip of the fungal strain was cut with a sterile cork borer and transferred to 25 mL of induced medium (35 g/L of sucrose, 5 g/L of peptone, 2.5 g/L of yeast extract, 0.5 g/L of MgSO4, 1 g/L of KH2PO4, and 0.05 g/L of vitamin B1, adjusted to pH 5.2 in a 250 mL flask, and grown at 28°C.18 The culture broth was collected from 5 flasks every week for the small-scale time course fermentation (1, 10 weeks), while for the large-scale fermentation, 100 flasks were collected at day 20. The culture broth was filtered through a 0.2 µm filter membrane before extraction.
Extraction and Isolation
For the small-scale extraction, the culture broth was extracted twice with an equal volume of ethyl acetate. The combined ethyl acetate layer was concentrated under reduced pressure to obtain an extract which was further subjected to analyses by HPLC. For the large-scale extraction, the culture broth (2.5 L) was divided into 5 portions (500 mL) and each portion was extracted with ethyl acetate (300 mL, ×3). The collected ethyl acetate layer was combined and concentrated under reduced pressure to obtain the ethyl acetate extract (8.9 g). This was further triturated in ethyl acetate at room temperature and stirred for 24 hours (50 mL, ×2) to yield 1 (8.0 g) and an ethyl acetate-soluble fraction (520 mg). The ethyl acetate-soluble fraction was subjected to purification by silica gel and Sephadex G-75 CC to give 2 (12. 3 mg) as a pale brownish oil.
The extract samples for HPLC were prepared at 2 mg/mL in methanol: water (1:1) while the standard compounds were prepared at 100 µg/mL in water. Each sample solution of 50 µL was injected onto a reversed phase C18 column. Methanol: milli-Q water (15:85 v/v) was used as a mobile phase with a constant flow rate of 1 mL/min. The column temperature was set at 30°C and the total run time was 30 min. All samples were detected at 254 nm using a diode array detector.
Antifungal Activity Assay (Pore Plate Technique)
Compound 1 (5 mL) as a solution in water at a concentration of 500 µg/mL was mixed in 95 mL PDA medium before being plated onto 90 mm Petri dishes. Seven-day-old mycelial discs of 6 plant pathogenic fungal pathogens (Colletotrichum musae, Colletotrichum capsici, Colletotrichum gloeosporioides, Pestalotia spp., Fusarium spp. TFPK301 and Fusarium spp. Foc 1708) were cut with a 7 mm sterilized cork borer under aseptic conditions and placed onto 25 ml PDA plates containing 25 µg/mL (final concentration) of 1. The plates were incubated at 28°C and the mycelial growth was determined at day 7. The percentage of mycelial growth inhibition (PGI) was calculated using the formula; PGI (%) =R− R1/R × 100, where R represents the fungal growth radius (mm) of the control culture and R1 represents the fungal growth radius distance (mm) in the treatment culture.19 The experiment was replicated 5 times, and the fungus grown on the PDA plate was used as the control.
Biological Assay
Anti- HSV-1 and cytotoxicity assays against human breast cancer (MCF-7), oral human epidermoid carcinoma (KB), and Vero (African green monkey kidney fibroblasts) cell lines were evaluated using a colorimetric method.20
Crystal Data of 1
A single crystal of 1 was mounted on the end of a hollow glass fiber. X-ray diffraction data were collected using a Bruker D8 VENTURE and operating at T = 296(2) K. Data were measured using ω and ϕ scans and using Cu-Kα radiation (λ = 1.54056 Å). The total number of runs and images was based on the strategy calculation from the program APEX3 and unit cell indexing was refined using SAINT (V8.38A, Bruker, 2016). Data reduction and scaling were performed using SAINT (V8.38A), and SADABS-2016/2 was used for absorption correction (APEX3, SADABS and SAINT; Bruker AXS Inc., Madison, Wisconsin, USA, 2016). The structure was solved with the ShelXT structure solution program using combined Patterson and dual-space recycling methods.21 The structure was refined by least squares using ShelXL.22 All non-hydrogen atoms were refined anisotropically. The hydrogen atoms of organic ligands were placed in calculated positions and refined using a riding model on attached atoms with isotropic thermal parameters 1.2 times those of their carrier atoms. The O−H hydrogen atoms were located in difference Fourier maps, but refined with O−H = 0.82 ± 0.01 Å. The data have been deposited with the Cambridge Crystallographic Data Center (CCDC) with CCDC number 1 843 739.
Footnotes
Acknowledgements
N. Surapong acknowledges the Science Achievement Scholarship of Thailand (SAST) and MSU Outbound Internship in 2015 and the School of Chemistry, University of Wollongong (UOW), Australia. P. Seephonkai thanks the Center of Excellence for Innovation in Chemistry (PERC-CIC) and Faculty of Science, Mahasarakam University for partial financial support for this project. We thank Dr Praphat Kawicha from Kasetsart University Chalermphrakiat Sakonnakhon Province Campus for providing the plant pathogenic fungi Fusarium spp. TFPK301 and Fusarium spp. Foc 1708.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article: received partial financial support from Faculty of Science, Mahasarakham University.
Supplemental Material
References
1.
SPL.LiP.DongTTX.TsimKWK. Anti-oxidation activity of different types of natural Cordyceps sinensis and cultured Cordyceps mycelia. Phytomedicine. 2001;8(3):207–212.
2.
KimHG.ShresthaB.LimSYet al. Cordycepin inhibits lipopolysaccharide-induced inflammation by the suppression of NF-κB through Akt and p38 inhibition in RAW 264.7 macrophage cells. Eur J Pharmacol. 2006;545(2-3):192–199.doi:10.1016/j.ejphar.2006.06.047
IsakaM.TanticharoenM.KongsaereeP.ThebtaranonthY. Structures of cordypyridones A-D, antimalarial N-hydroxy- and N-methoxy-2-pyridones from the insect pathogenic fungus Cordycepsnipponica. J Org Chem. 2001;66(14):4803–4808.doi:10.1021/jo0100906
5.
SeephonkaiP.IsakaM.KittakoopPet al. A new tropolone from the insect pathogenic fungus Cordyceps sp. BCC 1681. J Antibiot. 2001;54(9):751–752.doi:10.7164/antibiotics.54.751
6.
RukachaisirikulV.PramjitS.PakawatchaiC.IsakaM.SupothinaS. 10-membered macrolides from the insect pathogenic fungus Cordycepsmilitaris BCC 2816. J Nat Prod. 2004;67(11):1953–1955.doi:10.1021/np0401415
7.
IsakaM.BoonkhaoB.RachtaweeP.AuncharoenP. A xanthocillin-like alkaloid from the insect pathogenic fungus Cordyceps brunnearubra BCC 1395. J Nat Prod. 2007;70(4):656–658.doi:10.1021/np060509t
8.
WangY.YinH.LvX.WangY.GaoH.WangM. Protection of chronic renal failure by a polysaccharide from Cordyceps sinensis. Fitoterapia. 2010;81(5):397–402.doi:10.1016/j.fitote.2009.11.008
9.
AramwitP.BangN.RatanavarapornJ.NakphengT.SrichanaT. An anti-cancer cordycepin produced by Cordyceps militaris growing on the dead larva of Bombyx mori silkworm. J Agric Sci. 2014;6(6):41–53.doi:10.5539/jas.v6n6p41
10.
NakamuraK.ShinozukaK.YoshikawaN. Anticancer and antimetastatic effects of cordycepin, an active component of Cordyceps sinensis. J Pharmacol Sci. 2015;127(127):53–56.doi:10.1016/j.jphs.2014.09.001
11.
SangdeeK.NakbanpoteW.SangdeeA. Isolation of the entomopathogenic fungal strain Cod-MK1201 from a cicada nymph and assessment of its antibacterial activities. Int J Med Mushrooms. 2015;17(1):51–63.doi:10.1615/IntJMedMushrooms.v17.i1.60
12.
BunyapaiboonsriT.YoiprommaratS.IntereyaK.KocharinK. New diphenyl ethers from the insect pathogenic fungus Cordyceps sp. BCC 1861. Chem PharmBull. 2007;55(2):304–307.doi:10.1248/cpb.55.304
13.
WangJ-hui.ZhangZ-liang.WangY-qinet al. Chemical constituents from mycelia and spores of fungus Cordyceps cicadae. Chinese Herbal Medicines. 2017;9(2):188–192.doi:10.1016/S1674-6384(17)60094-7
14.
SangdeeK.SeephonkaiP.BuranratB.SurapongN.SangdeeA. Effects of ethyl acetate extracts from the Polycephalomyces nipponicus isolate Cod-MK1201 (Ascomycetes) against human pathogenic bacteria and a breast cancer cell line. Int J Med Mushrooms. 2016;18(8):733–743.doi:10.1615/IntJMedMushrooms.v18.i8.90
15.
AyerWA.GökdemirT.MiaoS.TrifonovLS.LeptosphaeronesA. Leptosphaerones A and B, new cyclohexenones from Leptosphaeria herpotrichoides. J Nat Prod. 1993;56(9):1647–1650.doi:10.1021/np50099a034
16.
LinZ.ZhuT.FangY.GuQ.ZhuW. Polyketides from Penicillium sp. JP-1, an endophytic fungus associated with the mangrove plant Aegiceras corniculatum. Phytochemistry. 2008;69(5):1273–1278.doi:10.1016/j.phytochem.2007.10.030
17.
JaihanP.SangdeeK.SangdeeA. Disease suppressive activity of extracts from entomopathogenic fungus Ophiocordyceps sobolifera against chili anthracnose fungi Colletotrichum spp. in a pot experiment. J Gen Plant Pathol. 2018;84(3):237–242.doi:10.1007/s10327-018-0778-3
18.
HuangL.QZL.ChenYY.WangXF.ZhouXW. Determination and analysis of cordycepin and adenosine in the products of Cordyceps spp. Afr J Microbiol Res. 2009;3:957–961.
19.
KumerAS.Eswara ReddyNP.Hariprasad ReddyK.Charitha DeviM. Evaluation of fungicidal resistance among Colletotrichum gloeosporioides isolates causing mango anthracnose in agri export zone of Andhra Pradesh, India. Plant Pathol Bull. 2007;16:157–160.
20.
SkehanP.StorengR.ScudieroDet al. New colorimetric cytotoxicity assay for anticancer-drug screening. J Natl Cancer Inst. 1990;82(13):1107–1112.doi:10.1093/jnci/82.13.1107
21.
SheldrickGM. SHELXT - integrated space-group and crystal-structure determination. Acta Crystallogr A Found Adv. 2015;71(1):3–8.doi:10.1107/S2053273314026370
22.
SheldrickGM. Crystal structure refirement with SHELXL. Acta Crystallogr C Struct Chem. 2015;7(Pt 1):3–8.
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.