
Abstract
Although immune checkpoint inhibitors (ICIs) have greatly improved the prognosis of some cancer patients, the majority still fail to respond adequately, and the available biomarkers cannot reliably predict drug efficacy. The gut microbiota has received widespread attention among the various intrinsic and extrinsic factors contributing to drug resistance. As an essential regulator of physiological function, the impact of gut microbiota on host immunity and response to cancer therapy is increasingly recognized. Several studies have demonstrated significant differences in gut microbiota between responders and nonresponders. The gut microbiota associated with better clinical outcomes is called ‘favorable gut microbiota’. Significantly, interventions can alter the gut microbiota. By shifting the gut microbiota to the ‘favorable’ one through various modifications, preclinical and clinical studies have yielded more pronounced responses and better clinical outcomes when combined with ICIs treatment, providing novel approaches to improve the efficacy of cancer immunotherapy. These findings may be attributed to the effects of gut microbiota and its metabolites on the immune microenvironment and the systemic immune system, but the underlying mechanisms remain to be discovered. In this review, we summarize the clinical evidence that the gut microbiota is strongly associated with the outcomes of ICI treatment and describe the gut microbiota characteristics associated with better clinical outcomes. We then expand on the current prevalent modalities of gut microbiota regulation, provide a comprehensive overview of preclinical and clinical research advances in improving the therapeutic efficacy and prognosis of ICIs by modulating gut microbiota, and suggest fundamental questions we need to address and potential directions for future research expansion.
Introduction
Immune checkpoint inhibitors (ICIs) have revolutionized the field of cancer therapy. Although chemotherapy and radiotherapy remain the primary treatments for most cancers, ICIs are now effective treatments for many unresectable solid or nonsolid tumors, with response rates between 20% and 40% for most tumor indications. 1 ICIs also have excellent performance in neoadjuvant and adjuvant therapy. Neoadjuvant nivolumab did not delay surgery and caused major pathologic responses in nearly half (45%) of resectable non-small-cell lung cancer (NSCLC) with minimal side effects. 2 Pembrolizumab as adjuvant therapy for stage IIB or IIC melanoma significantly reduces the risk of disease recurrence or death compared to placebo while having a manageable safety profile. 3 As the most commonly used immunotherapy, ICIs work by relieving the functional suppression of T cells, inducing potent activation of the immune system and an effective antitumor response. To date, the US Food and Drug Administration has approved ICIs against four different targets for use in humans: antibodies against cytotoxic T lymphocyte-associated protein 4 (CTLA-4), programmed cell death protein 1 (PD-1) and its ligand, PD-L1, and lymphocyte-activation gene 3 protein.4,5 Over the years, the clinical use of anti-PD-1 and PD-L1 antibodies has significantly surpassed that of anti-CTLA-4 antibodies because of greater clinical efficacy and better tolerability.
Although some patients experienced dramatic tumor regression after treatment with ICIs, most did not respond. Among the melanoma patients with the highest response rates to ICIs, 40–50% showed no objective response to the combination of nivolumab and ipilimumab.6,7 In patients with mismatch repair proficient/microsatellite stable metastatic colorectal cancer, no efficacy was observed with ICIs alone, 8 and the objective response rate in a phase II clinical trial of durvalumab combined with tremelimumab was only 1%. 9 Why most patients do not respond to ICIs deserves to be studied. A complex and interconnected set of factors, such as host, tumor, and environmental variables, 10 can contribute to drug resistance, including gut microbiota. The gut microbiota plays a vital role as a regulator of physiological functions, including obtaining nutrients from the diet, maintaining the integrity of the mucosal barrier, and promoting the development and homeostasis of the immune system. 11 It affects the threshold of the anticancer immune response.12,13 Increasing numbers of preclinical and clinical studies have demonstrated that the efficacy and prognosis of ICI treatment are closely related to gut microbiota diversity and variety. 14 Therefore, strategies to modulate gut microbiota [e.g. fecal microbiota transplantation (FMT), probiotics, and dietary interventions] are being developed and used to improve clinical outcomes in patients using ICIs.
In this review, we present evidence for the role of gut microbiota in the therapeutic response to ICIs, outline the potential mechanisms, summarize the advances in research modulating the efficacy of ICIs by intervening with gut microbiota, and provide an outlook on future directions.
The gut microbiota is closely related to the efficacy of ICIs
The gut microbiota regulates cancer at multiple levels of initiation, genetic instability, susceptibility to host immune response, progression, and response to treatment. 15 There is evidence that the gut microbiota may play a role in the response to chemotherapy. 16 The immune system is an essential pathway by which the microbiome influences the chemotherapeutic response.16,17 As a treatment that is most closely associated with the immune response, the relationship between ICIs and the gut microbiota has attracted attention.
Two landmark studies published in 2015 uncovered a link between gut microbiota and ICIs in preclinical murine models. They indicated that CTLA-4 and PD-1 blockade only reduced tumor growth in mice carrying specific commensal bacteria and that modulation of gut microbiota could enhance treatment response.18,19 Three subsequent studies in 2018 further explored the gut microbiota characteristics of responders (Rs) and nonresponders (NRs) to ICIs and successfully reduced tumor volumes in preclinical murine models by FMT from responders.20–22 Moreover, two independent cohorts showed that antibiotic-induced dysbiosis adversely affected the response to ICI therapy.22,23 The importance of specific bacterial composition has been supported by several clinical studies investigating the relationship between different gut microbiota characteristics and clinical benefits (Supplemental Table 1).20–22,24–43 Most of these studies focused on NSCLC, melanoma, and hepatocellular carcinoma, where alpha diversity and some specific gut microbiome constituents have been associated with successful treatment with ICIs, such as Bifidobacterium species, Lactobacillus species, Akkermansia species, Faecalibacterium species, Firmicutes species, and Lachnospiraceae. A favorable gut microbiome improves effector T-cell function in the peripheral and tumor microenvironment (TME).44,45 Bacteria currently identified as more abundant in responders were associated with an increased frequency of dendritic cells (DCs) and greater T helper cell 1 (TH1) responses. 46 For example, signals from commensal Bifidobacterium and Akkermansia muciniphila regulate the activation of DCs at a steady state and improve the effector function of tumor-specific CD8+ T cells.18,47 A. muciniphila generates a ‘balanced’ commensal colonization, which induces local and systemic recall of TH1 immune responses and enhances tumor immunosurveillance.22,46,48
Due to the limited sample size, differences in the characteristic microbiome among cancers were not detected. Even in patients with the same cancers responding to the same ICIs, a low overlap of gut microbiota was observed, which may be related to different reference databases, diverse sample analysis techniques, and individual differences. It may also suggest that, rather than individual strains, the integral gut ecosystem is more likely to be a key determinant of the response to ICIs. The human gut microbiome includes multiple discrete ecologically balanced communities.49,50 Most current studies link individual taxa to medication outcomes, but little is known about the gut ecosystem. McCulloch et al. 28 transformed multidimensional 16S rRNA gene amplicon data into two-dimensional flat data using a t-distributed stochastic neighbor embedding plot to define 27 ‘microbiotypes’ and divided them into two favorable and two unfavorable ‘superclusters’ based on the proportion of patients with different outcomes. 28 This approach presents the data visually and concisely while retaining important features, shifting the focus from individual taxa to gut microbiotypes and translating a large amount of scattered community information into a simplified, well-distinguished general profile that is more useful for predicting outcomes.
In addition, some studies use gut microbiota as a biomarker of therapeutic response. Patients were classified into clusters based on the intrinsic gut microbiota composition at baseline by unsupervised hierarchical clustering, these clusters had different response ratios to immunotherapy.19,20,25,26,42 Peng et al. further developed machine learning models from patients’ baseline characteristics of gut microbiota to predict ICI efficacy effectively. The area under the curve of the elastic net model without polynomial features reached 0.78, 40 which suggested that gut microbiota may not be an excellent stand-alone biomarker and require integration with other known biomarkers to exert its predicted function to the utmost extent.
Modulating the efficacy of ICIs by regulating gut microbiota
There is growing evidence that the gut microbiota has a role in influencing the efficacy of ICIs in preclinical models and cancer patients. In particular, antibiotic treatment prior to ICIs inhibits clinical efficacy, further suggesting that disruption of the balanced gut microbiota (i.e. switch from eubiosis to dysbiosis) and loss of specific bacteria may impede the success of anticancer therapy.23,51 A lack of immunostimulatory bacteria or a dysregulated gut ecosystem containing excessive immunosuppressive bacteria can lead to therapeutic failure. Therefore, modulating gut microbiota composition may be an effective strategy to avert ICI resistance. The gut microbiota composition is determined by host genetics, colonization at birth, type of delivery, individual lifestyle, disease, and drug exposure.52–55 It remains relatively stable in adulthood 56 but can be altered by external interventions or disturbances in the internal environment. Clinical studies preliminarily evaluated the effectiveness of improving immunotherapy outcomes through gut microbiota modulation (Table 1).24,29,57–63 Additional clinical trials have been initiated, which may provide practical and accessible tools for future adjuvant cancer therapy (Table 2).
Clinical studies on modulating gut microbiota composition to improve the efficacy of ICIs.
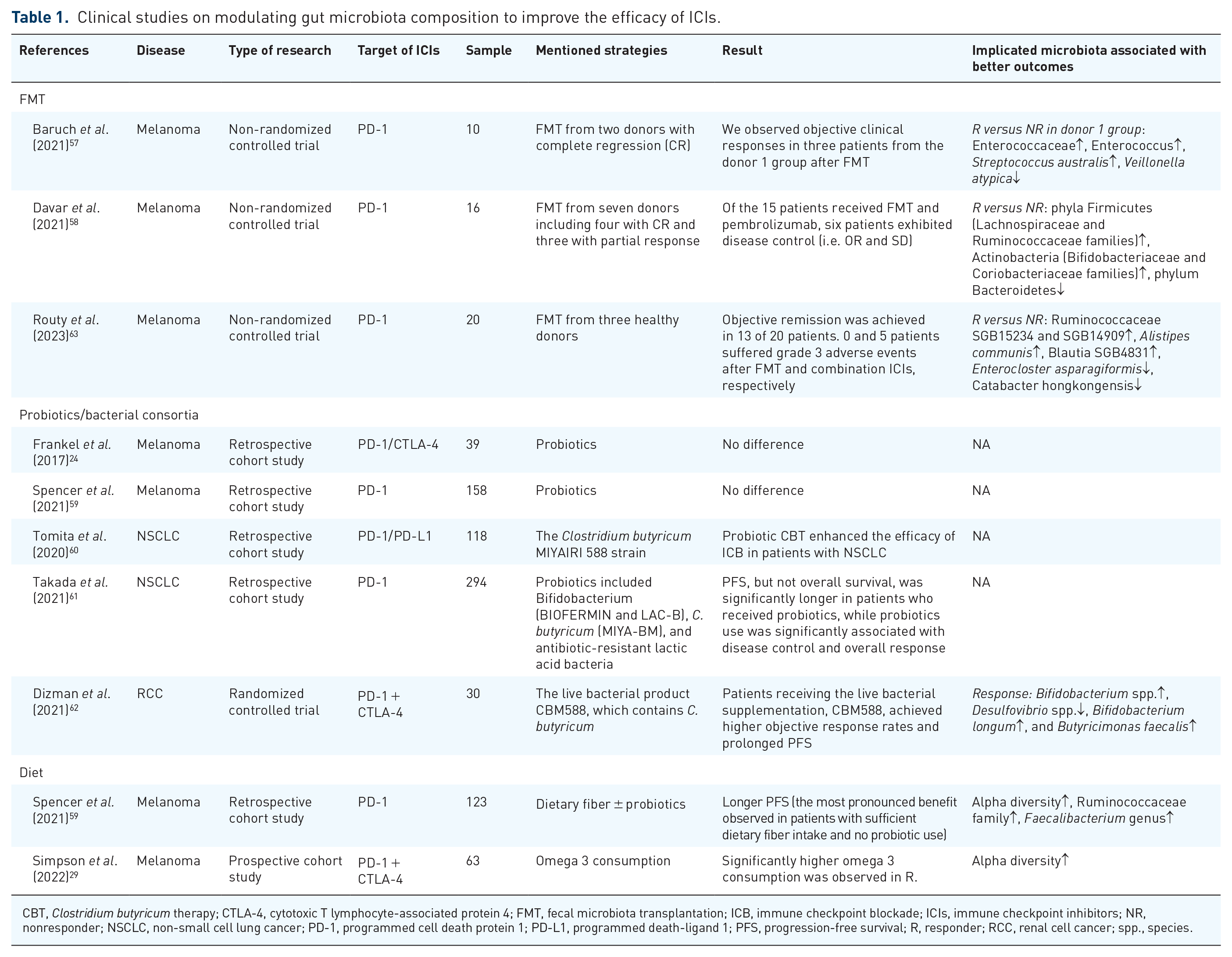
CBT, Clostridium butyricum therapy; CTLA-4, cytotoxic T lymphocyte-associated protein 4; FMT, fecal microbiota transplantation; ICB, immune checkpoint blockade; ICIs, immune checkpoint inhibitors; NR, nonresponder; NSCLC, non-small cell lung cancer; PD-1, programmed cell death protein 1; PD-L1, programmed death-ligand 1; PFS, progression-free survival; R, responder; RCC, renal cell cancer; spp., species.
Ongoing trials of gut microbiome modulation (FMT/probiotics/bacterial consortia) in cancer immunotherapy.

dMMR, mismatch repair deficiency; FMT, fecal microbiota transplantation; ICIs, immune checkpoint inhibitors; MSS, microsatellite stable; NSCLC, non-small-cell lung cancer; PD-1, programmed cell death protein 1; PD-L1, programmed death-ligand 1; pMMR, mismatch repair proficient.
Fecal microbiota transplantation
FMT is the administration of fecal matter from a donor into the intestinal tract of a recipient to directly change the recipient’s microbial composition and confer a health benefit. 64 It is effective in numerous diseases, such as irritable bowel syndrome (IBS), inflammatory bowel disease (IBD), metabolic disorders, autoimmune diseases, and cancer.65,66 FMT transplants the entire gut microbial ecosystem from the donor containing beneficial bacteria and overall diversity, which enables the introduced bacteria to be more stable and less subject to competitive rejection by the recipient microbiota.
Due to imperfect quality control assessment, pathogenic bacteria, especially multidrug-resistant bacteria, and other pathogens may be unknowingly transferred in fecal material, resulting in severe infections and complications. 67 In addition, complicated operating procedures make the products expensive and challenging to access.
Nevertheless, several studies have demonstrated that FMT effectively improved the efficacy and prognosis of ICIs in preclinical murine models (Supplemental Table 2).18–22,34,39,59,63,68–74 Transplantation of fecal matter from Jackson Laboratory (JAX) mice with better responses into Taconic Farms (TAC) mice significantly improved the efficacy of anti-PD-L1 treatment. It increased the abundance of Bifidobacteria in the gut, a highly regarded genus of gut microbiota, whose existence is associated with the accumulation of activated antigen-specific T cells in the TME. 18 FMT from responders combined with ICIs successfully inhibited tumor growth in mice and caused a similar increase in fecal Bifidobacteria abundance. 74 Vetizou et al. 19 demonstrated that FMT from cluster C patients carrying a higher abundance of Bacteroides fragilis combined with anti-CTLA-4 antibodies yielded the most dramatic tumor shrinkage in germ-free (GF) tumor-bearing mice compared to the control group. Dees et al. transferred fecal samples from five healthy human donors into mice to establish human-derived microbiome mice (HuM1–HuM5). When combined with anti-PD-1 treatment, the survival of HuM2 and HuM3 was significantly prolonged. 68 The above preclinical studies suggest that modulation of gut microbiota by FMT could influence the response to immunotherapy. The efficacy of FMT depends on the donors. Different donor-specific gut microbiota characteristics can lead to different clinical outcomes. How to identify the ideal donor is currently the most challenging issue. Some studies chose patients who have responded to previous treatments as donors,20–22,39 some preferred healthy volunteers, 68 and others selected populations carrying specific bacteria.19,71
The combined application of FMT from responders and anti-PD-1 treatment was well tolerated in patients with anti-PD-1 refractory melanoma, resulting in clinical benefit in 6 of 15 patients. FMT combination therapy induced microbiota fluctuations, with posttransplant responders exhibiting an increase in the abundance of taxa previously shown to be associated with anti-PD-1 treatment response. 58 In phase I clinical trial by Baruch et al., 3 of 10 patients with ICI-resistant metastatic melanoma responded successfully after FMT from the complete response population, including two partial responders and one complete responder. All three patients had received fecal matter from donor 1. Unlike the donor 2 group, patients in the donor 1 group had a higher abundance of Bifidobacterium adolescentis after transplantation. Further exploration of the mucosal lamina propria and intratumoral immune cell infiltration associated with the posttransplant gut microbiota revealed a significant increase in lamina propria CD68+ antigen-presenting cells (APCs). By contrast, only a subset of patients exhibited increased intratumoral CD8+ T cells, although they did not all produce responses after therapy. 57 These differences across patients suggest that beneficial gut microbiota is not the only determinant affecting immunotherapeutic response. Achieving the clinical translation of bacteria-related studies may consider the broader immune setting.
The results of the above clinical trials support the fact that FMT from responders could overcome resistance to ICIs in melanoma. Recently, a phase I clinical trial of FMT in combination with immunotherapy (NCT03772899) enrolled 20 ICI-naïve patients with advanced melanoma who underwent FMT via oral capsules derived from healthy donors and were treated with anti-PD-1 therapy 1 week later. Objective remission was achieved in 13 of 20 patients (65%), including 4 complete responders and 9 partial responders. Compared with NRs, Rs enriched in Ruminococcaceae SGB15234 and SGB14909, Alistipes communis, and Blautia SGB4831 1 month after FMT, with trends of higher T-cell infiltration and significant contact enrichment of antigen-experienced T cells to melanoma cells. The safety profile was favorable with 0 and 5 patients suffering grade 3 adverse events after FMT and combination ICIs, respectively. 63 The clinical efficacy data show superior outcomes to anti-PD-1 monotherapy.75–79 Meanwhile, combination therapy did not increase the incidence of immune-related adverse events (irAEs) compared with PD-1 inhibitors alone.6,75,80,81 It demonstrated for the first time that FMT from healthy donors combined with PD-1 inhibitors could safely improve patients’ responses in advanced melanoma.
FMT was delivered mainly by two modalities, endoscopy or (and) oral capsules.57,58,63 Due to insufficient clinical experience, comparing the effects of different delivery modalities in immunotherapy is difficult. Each modality can improve the composition of the gut microbiota effectively and has been widely used in digestive diseases other than cancer.82,83 Endoscopic transplantation is the most direct way to accurately transplant larger amounts of intestinal bacteria into the implicated bowel segment with potentially greater clinical efficacy, especially in recurrent Clostridium difficile infection (CDI),84,85 IBS, 85 and IBD.86,87 However, endoscopic operations are more invasive and intolerable. Oral capsules have the advantage of being convenient and minimally invasive, with greater patient preference and compliance, but the efficacy in diseases other than CDI remains to be proven. 88
More clinical trials are currently underway (Table 2). Most trials use fecal matter from responders (e.g. NCT03341143), and a few use healthy individuals with specific favorable strains or with the same microbiota profile as responders as donors (e.g. NCT04163289). Identifying and functionalizing critical bacterial species that drive favorable clinical outcomes in ICIs are crucial and can help select suitable donors. Meanwhile, the optimal route for FMT has yet to be determined. Factors such as patient compliance and cost-effectiveness are important in selecting the appropriate transplantation modality. Whether there is heterogeneity in the favorable strains associated with the efficacy of ICIs in different cancers and whether the modulation of gut microbiota by FMT to improve the efficacy of ICIs is always sensitive and feasible are questions that need further exploration.
Bacterial consortia and probiotics
An alternative approach to modulating the gut microbiota is the oral administration of bacterial consortia or beneficial bacteria (i.e. probiotics). Concentrating on identified bacteria prevents the transfer of other pathogens and may provide stable therapeutic products that can be obtained and applied easily. However, there are substantial problems and discrepancies in bioavailability and standardization quality control. In addition, administration may cause a reduction in the overall diversity of gut microbiota. 89
As early as 2015, Sivan et al. 18 further reduced tumor volume by combining a mixture of B. breve and B. longum strains with anti-PD-L1. Subsequently, Bifidobacteria species were confirmed to have the potential to promote antitumor T-cell responses.21,38,90 A strain called B. animalis lactis (EDP1503) has been exploited in a clinical trial (NCT03775850) for adjuvant ICI therapy, demonstrating the ability of an orally delivered noncolonizing monoclonal microbe to enhance antitumor immunity. Various bacterial consortia have been shown to enhance the efficacy of ICIs in murine models (Supplemental Table 3).18,19,22,39,71,90–92 Specific B. bifidum strains (B. bif_K57, B. bif_K18, and B. bif_M31), A. muciniphila (Akk), Enterococcus hirae, A. indistinctus, or a mix of four Clostridiales strains (CC4) worked synergistically with anti-PD-1 to reduce tumor growth.22,90,92 Ampicillin + colistin + streptomycin (ACS)-treated mice orally fed B. thetaiotaomicron, B. fragilis, B. cepacia, or the combination of B. fragilis and B. cepacia recovered the anticancer response to anti-CTLA-4 antibodies. 19 Oral administration of immunostimulatory A. muciniphila or B. salyersiae before each cycle of ICIs compensated for the unsuccessful induction of sensitivity to treatment with NR-FMT. 39 Derosa et al. 71 found that resistance to anti-PD-1 treatment in mice was associated with the absence of detectable A. muciniphila in the FMT donor, and supplementation with exogenous Akkp2261 could shift the microbiome toward the favorable Akk-associated collateral ecosystem and reverse the resistance. In brief, bacterial consortia showed great potential in inducing effective anticancer responses in combination with ICIs, reversing drug tolerance caused by antibiotics, and compensating for insensitivity after ineffective FMT treatment.
Apart from conventional bacteria, Griffin et al. revealed that active enterococci express and secrete NlpC/p60 peptidoglycan hydrolase secreted antigen A (SagA) or its orthologs, which catabolize peptidoglycan to generate immune-active muropeptides and induce immunotherapeutic modulation. Oral administration of the engineered, SagA-expressing strain E. faecalis–SagA or intraperitoneal injection with active muramyl dipeptide (MDP)-L, D isomer successfully enhanced immunotherapeutic efficacy. 91 This indicates that the effectiveness of exogenous bacterial products can be altered by genetic engineering and provides a new direction for developing next-generation adjuvants.
Probiotics are one part of the bacterial consortia. They are living bacteria that provide health benefits to the host and show varying degrees of clinical efficacy in many gastrointestinal disorders.93–95 The efficacy of combination therapy with ICIs and probiotics varies (Table 1).24,59–62 Two retrospective studies in NSCLC patients showed that probiotic use improved immunotherapy efficacy and was associated with favorable prognostic outcomes. These probiotics included the C. butyricum MIYAIRI 588 strain, Bifidobacterium (BIOFERMIN and LAC-B), C. butyricum (MIYA-BM), and antibiotic-resistant lactic acid bacteria.60,61 Two other studies in melanoma patients did not observe a beneficial effect of probiotic intake history.24,59 Spencer et al. 59 even concluded that nonspecific probiotic intake, especially self-purchased over-the-counter probiotic supplements, is detrimental to the effectiveness of immunotherapy. However, they did not explicitly restrict the effective composition of probiotics, leading to unavoidable within-group differences. Given the potential drawbacks of retrospective studies, including selection bias and recall bias, the veracity and validity of these arguments need to be further considered.
Preclinical studies are warranted based on promising early data.18,47,60 Oral probiotics combined with antitumor immunotherapy demonstrated superior efficacy in murine models. The commercial cocktail of Bifidobacterium, including B. breve and B. longum, was already shown to reduce tumor growth significantly when combined with anti-PD-L1 in 2015. Five subsequent studies using Lactobacillus-based probiotics similarly yielded preferable results compared to immunotherapy alone, assessed by tumor size and survival.73,96–99 Together, these studies support the need for more careful investigations of current commercially available probiotic formulations (Supplemental Table 3).59,73,96–99 Nonetheless, the self-administration of probiotics during immunotherapy is discouraged because our understanding of how these microorganisms affect the immune system and therapeutic response is limited. These supplements are generally unregulated in the European Union and the United States. There may be significant differences in quality, 100 which affect the efficacy and safety of these products.
Only one interventional clinical trial has presented preliminary results. The open-label, single-center, randomized, controlled study showed that 19 treatment-naïve patients with metastatic renal cell carcinoma receiving nivolumab–ipilimumab plus probiotics (CBM588) achieved higher objective response rates (58% versus 20%, p = 0.06) and prolonged median progression-free survival (PFS; 12.7 months versus 2.5 months, p = 0.001) compared with 10 patients receiving nivolumab–ipilimumab alone. 62 CBM588 contains a butyrate-producing anaerobic spore-forming bacterium, C. butyricum. A retrospective study of NSCLC patients receiving ICIs showed that CBM588 significantly affected both PFS and overall survival. 60 Preclinical studies have also demonstrated that C. butyricum can suppress the development of gastrointestinal tumors by inhibiting the Wnt/β-catenin and NF-κB signaling pathways, modulating the intestinal microbiota and tumor development.101–103 In patients receiving CBM588 and responding to treatment, an increase in Bifidobacterium species was observed, but not all patients taking CMB588 produced an effective elevation of Bifidobacterium species. 62 How probiotics can effectively improve the gut microbiota and the efficacy of immunotherapy needs to be further addressed.
Although the clinical trial presented favorable results, the instability of probiotics or other bacterial products in the intestine caused by the resistance of the original bacteria prevented follow-up research and the clinical application of this technology. Previous studies have primarily used antibiotic pretreatment to prepare the gut ecosystem. Han et al. 104 found that smectite promoted the formation of Lactobacillus and Bifidobacterium biofilms on its surface, which could prime DCs through toll-like receptor (TLR) 2 signaling, activate CD8+ T-cells and release interferon-γ (IFN-γ) and interleukin-12 (IL-12), and finally enhance the efficacy of immunotherapy in mice. Using smectite to establish colonization sites specifically for probiotics is safer and more effective in retaining specific strains and is potentially translatable.
Clinical trials of probiotics in combination with immunotherapy are being conducted gradually (Table 2). Several hospitals in China have expanded the research direction to other cancers, such as NCT05220124 and NCT05032014, focusing on bladder and liver cancer, respectively. Indeed, further research is needed to determine how effectively probiotics can improve immunotherapy efficacy. We should take a very cautious position until the safety of commercially available probiotics is thoroughly evaluated in preclinical and clinical studies.
Diet and prebiotics
Dietary intake can contribute to different microbiome compositions, changes in dietary regimen significantly alter the gut microbiota within 5 days. 105 Although responders to ICIs have different bacterial compositions, these taxa have something in common regarding their upstream dietary substrates.106,107 For example, Faecalibacterium prausnitzii and Ruminococcus species consume dietary fiber, A. muciniphila and Bifidobacterium species use polyphenols of food origin as substrates. Therefore, there is a solid case to evaluate the effect of dietary interventions on ICI efficacy in the context of gut microbiota.
With excellent safety, accessibility, and low cost, dietary interventions are simple ways to regulate the gut microbiota in patients using ICIs. However, behavioral changes, particularly dietary patterns, are notoriously tricky, resulting in a paucity of prospective interventional studies and evidence-based dietary guidelines for patients to follow. As a widely known healthy diet, a high-fiber diet can be helpful for immunotherapy.59,69 It could enhance the response to ICIs in preclinical mouse models, which is closely related to increased fiber-fermenting bacteria.
Dietary fiber is the main subject of current diet-related research. Indeed, consuming specific dietary components, such as dietary fiber and prebiotics, is a common way to regulate the microbiota. Intervention clinical trials have shown that dietary fiber intake increases gut bacterial diversity. 108 Dietary fiber intake may affect clinical outcomes in patients treated with ICIs. A retrospective cohort study showed that patients treated with ICIs taking sufficient dietary fiber intake had significantly longer PFS than those taking insufficient dietary fiber intake (median PFS not reached versus 13 months), with the longest PFS observed in patients reporting sufficient dietary fiber intake and no probiotic use, in which microbial alpha diversity, Ruminococcaceae family, and Faecalibacterium genus abundances were numerically higher, although the results did not reach statistical significance. 59
Bindels et al. 109 defined prebiotics as ‘a nondigestible compound that, through its metabolization by microorganisms in the gut, modulates composition and activity of the gut microbiota, thus conferring a beneficial physiological effect on the host’. As the definition says, prebiotics are rich in variety, and most of them belong to dietary fiber. Different prebiotics have different effects on the regulation of gut microbiota. For example, the plant polysaccharide inulin can stimulate the growth of fiber-fermenting Faecalibacterium and Bifidobacterium species.110,111 Supplementation with inulin and fructo-oligosaccharide may select beneficial bacteria such as Lactobacillus and Bifidobacterium. 111 These taxa are associated with enhanced responses to ICIs.20,21 Inulin, a heterogeneous mixture of fructose polymers, has been recognized as the most widely known prebiotic with nutritional and therapeutic effects to improve health. Han et al. 112 enhanced the antitumor activity of anti-PD-1 antibodies by combining inulin gel in CT26 murine models, which increased the relative abundance of crucial commensal microbiota such as Akkermansia, Lactobacillus, Roseburia, and their short-chain fatty acid (SCFA) metabolites, and induced systemic memory T-cell responses.
In addition, Simpson et al. 29 found that omega-3 intake was significantly higher in patients who responded to immunotherapy. A variety of other prebiotics, specific diets, nutrients, or bioactive compounds from them may also suppress tumors and enhance ICI efficacy.74,113–115 These interventions have value for further exploration (Supplemental Table 4).35,59,69,70,74,91,104,112,115,116
Modulation of gut microbiota affects irAEs of immune checkpoint inhibitors
Research has demonstrated that irAE occurrence is connected to better ICI response and long-term benefits,117,118 making early and appropriate management a critical task. Nevertheless, the relationship between specific genera and irAEs has yielded controversial results. Certain bacteria have been identified to facilitate ICI responses and reduce ICI toxicities, including Akkermansia, Bifidobacterium, etc. 119 Chaput et al. 25 showed that patients treated with ipilimumab with increased representation of Faecalibacterium and Firmicutes had increased efficacy of ICIs, as well as increased risk of developing colitis or other ICI-induced toxicities, which is known as the efficacy-toxicity coupling effect.
While improving the efficacy of immunotherapy through modulating gut bacteria, the diversity and abundance of microbes might also influence ICI therapy’s immunotoxicity. In a currently published clinical trial, the efficacy of ICIs can be safely modulated by FMT. 63 However, it is difficult to draw a definite conclusion regarding highly diverse and multifaceted microbial-immune system interactions. We still need to monitor the possible induction or attenuation of irAEs in immunotherapy and find the balance between ICI response and irAEs step by step.
Gut microbiota modulates the function of immune checkpoint inhibitors by several potential mechanisms
Although studies suggest that the gut microbiota may be an important factor in the therapeutic response to ICIs, the mode of action remains to be elucidated. We summarize the potential mechanisms as follows (Figure 1).

Gut microbiota modulates the antitumor immunity by several potential mechanisms in immunotherapy. (a) Microbial proteins cause T-cell cross-reactivity by mimicking tumor antigens. (b) Gut microbiota creates a relatively “hot” tumor microenvironment by regulating the expression and distribution of immune checkpoint molecules. (c) Microbes or microbial products activate PRRs, promoting tumor cell death by directly inducing or indirectly activating adaptive immune cells via cytokines released from surrounding cells or recruitment and activation of antigen-presenting cells to prime effector T cells. (d) Gut microbiome-derived SCFAs increase specific metabolites in the blood circulation of mice, particularly α-ketoglutaric acid, N-acetyl-l-glutamic acid, and pyridoxine, promoting the infiltration and activation of cytotoxic T lymphocytes and inhibiting the function of Tregs. (e) The intratumoral microbiome could be modulated by the intestinal microbiome, which might translocate to, colonize, and persist within tumors. Their peptides can be presented by the HLA-I and HLA-II molecules of melanoma cells and elicit T-cell cross-reactivity. Intratumoral Lactobacillus reuteri promotes antitumor T-cytotoxic 1 immunity via its metabolites, I3A, which acts through CD8+ T-cell-specific AhR signaling to promote IFN-γ and Gzm-B production in a CREB-dependent manner. (f) Inflammatory pathways can be detected in most of the pathways of immunotherapy regulated by gut microbiota, especially the T-cell receptor signaling pathway.
Regulate the expression and distribution of immune checkpoint molecules
The TME consists of many different cell types that play an essential role in tumor tolerance and evasion of immune surveillance 120 and largely influence the efficacy of ICIs,121,122 including T cells, fibroblasts, natural killer (NK) cells, and DCs.123–125
Chaput et al. found that the commensal bacterium F. prausnitzii was associated with a decrease in circulating α4+β7+ T cells and CD4+ Tregs. In Crohn’s disease-based studies, F. prausnitzii correlated with the induction of regulatory T cells in the gut. 126 Therefore, it was speculated that F. prausnitzii might be related to the sequestration of a4+b7+ T cells and Tregs in the intestine and tumor bed. 25 Tregs express high levels of CTLA-4 and are susceptible to ipilimumab. 127 In the presence of F. prausnitzii, ipilimumab induces the inactivation of a more extensive range of CTLA-4+ Treg cells, reactivating effector T cells,128,129 and ultimately improving the efficacy of the treatment. In addition, Gopalakrishnan et al. showed that transplantation of favorable gut microbiota upregulated PD-L1 expression in the TME in murine models. It induced a ‘hot’ TME and caused more pronounced tumor shrinkage. 20 The gut microbiota creates a relatively ‘hot’ microenvironment for ICIs by inducing aggregation or elevated expression of immune checkpoint molecules, which improves efficacy.
Programmed death ligand 2 (PD-L2) is a member of the same B7 family proteins as PD-L1 and exerts immunomodulatory effects by binding to PD-1 and repulsive guidance molecule b (RGMb) as a ligand. 130 In 2017, Yearley et al. 131 revealed that the expression level of PD-L2 was closely associated with the clinical efficacy of pembrolizumab for head and neck squamous cell carcinoma, and the efficacy prediction effect was independent of PD-L1 expression. 131 Besides, PD-L2 expressed on DCs can bind to RGMb on CD8+ T cells, inhibiting CD8+ T-cell responses. Specific gut microbiota, especially Coprobacillus cateniformis, inhibits PD-L2 expression on DCs, activates antitumor immunity, and improves the efficacy of ICIs. 132 Therefore, downregulation of the PD-L2-RGMb pathway may serve as a mechanism by which gut microbiota modulates the response to ICIs.
Activate antitumor immunity
One pathway is that microbial proteins cause T-cell cross-reactivity by mimicking tumor antigens. The theory of ‘molecular mimicry’ suggests that specific CD4+ or CD8+ T lymphocytes induced by major histocompatibility complex class I and class II binding epitopes encoded by bacterial or viral genomes may unexpectedly recognize autoantigens and attack tumor tissues.133,134
Microbial-specific T lymphocytes can activate an effective antitumor immune response. Deficits in the efficacy of ICIs were overcome by the adoptive transfer of B. fragilis-specific T-cells. 19 Memory responses of IFN-γ-secreting CD4+ and CD8+ T cells specifically targeting E. hirae, B. fragilis, and A. muciniphila are associated with good clinical outcomes in cancer patients.18,19,135,136 A retrospective analysis of long-term pancreatic cancer survivors showed sequence homology of high-quality neoantigens with microbial epitopes, 133 suggesting the basis for T-cell cross-reactivity.
Until 2020, Bessell et al. 137 found that T cells targeting an epitope expressed in the commensal bacterium B. breve SVYRYYGL cross-reacted with the model neoantigen SIYRYYGL expressed in B16.SIY tumor. 137 Ragone et al. 138 further showed a high homology in the linear sequence as well as in structure and conformation between tumor-associated antigens and peptides derived from the microbiota of the Firmicutes and the Bacteroidetes phyla, possessing the basis for cross-reactive T-cell responses. 138 TCC88, a specific tumor-infiltrating lymphocyte (TIL)-derived CD4+ T-cell clone, recognizes multiple glioblastoma-derived peptides and responds strongly to a broad spectrum of gut microbiota-derived targets, especially peptides from B. fragilis, Ruminococcus bromii, Alistipes, and Eubacterium genus, which elicit cross-reactive T-cell responses in bulk TILs and peripheral blood memory T cells. 139 These observations suggest T-cell cross-reactivity as a possible mechanism of effective immunotherapies due to certain gut microbiota composition.
The other pathway is that gut microbes or microbial products activate pattern recognition receptors (PRRs), leading to the activation of immune responses. Several PRR agonists have been proven to overcome resistance to ICI therapy.140–142 PRR stimulation can promote tumor cell death by directly inducing or indirectly activating adaptive immune cells via cytokines released from surrounding cells or recruitment and activation of APCs to prime effector T cells, 143 which shape a ‘hot’ TME and further enhance therapeutic efficacy. For example, gut microbiota triggered type I IFN production in DCs via stimulator of interferon genes (STING), a PRR in the form of endoplasmic reticulum transmembrane proteins, which enhanced cross-stimulation of antitumor CD8+ T cells and improved the efficacy of ICIs. 69 Active enterococci express and secrete orthologs of the NlpC/p60 peptidoglycan hydrolase SagA, which can generate nucleotide-binding oligomerization domain-containing protein 2 (NOD2)-active muropeptides and regulate the efficacy of checkpoint blockade immunotherapy in vivo by activating NOD2, increasing intratumor T-cell distribution. 91 Moreover, TLRs were proven to regulate anticancer immune activity through intestinal bacteria, 104 suggesting that gut microbiota regulation of the PRR pathway to influence therapeutic efficacy is widely available and mediated by multiple receptors. Microbiota species that activate specific PRRs may be leveraged as next-generation adjuvants.
Release metabolites
Metabolites of the gut microbiota suppress inflammation by maintaining the epithelial barrier and intestinal integrity and interact with host cells to influence the immune response.144,145 SCFAs are the most crucial metabolites, including acetate, propionate, and butyrate, which can modulate effector T-cell function by regulating G protein-coupled receptor signaling through a receptor-dependent mechanism.146,147 A cohort study of 52 patients with solid tumors revealed that high fecal acetate, propionate, butyrate, and valerate concentrations were significantly associated with prolonged PFS and higher response rates to anti-PD-1 therapy.20,148 NSCLC patients receiving anti-PD-1 therapy showed similar gut metabolomics, in which fecal SCFAs, particularly propionate, were associated with a better long-term response to ICIs. 31 The supplement of a probiotic strain, Lactobacillus rhamnosus Probio-M9 (Probio-M9), modulates the intestinal microbiome of tumor-bearing mice by increasing beneficial bacteria, promoting the accumulation of acetate, propionate, and butyrate in the intestine. SCFAs produced by these bacteria further increased specific metabolites in the blood circulation of mice, particularly α-ketoglutaric acid, N-acetyl-l-glutamic acid, and pyridoxine, promoting the infiltration and activation of cytotoxic T lymphocytes and inhibiting the function of Tregs, thereby enhancing the ICI response. 149 These results suggest the immunomodulatory capacity of SCFAs and their potential to promote immunotherapy efficacy in tumor–microbe communication.
Other metabolites also contribute to modulating immunotherapy. Trimethylamine N-oxide potentiated type-I IFN pathway and conferred antitumor effects in a type-I IFN-dependent manner, driving immune activation and boosting responses to ICIs in a mouse model of pancreatic ductal adenocarcinoma (PDAC). 150 Inosine produced by the commensal bacterium B. pseudolongum significantly promoted TH1 cell differentiation by exerting adenosine A2A receptors on T cells in the presence of exogenous IFN-γ and enhanced the therapeutic response to ICIs. 151
By specifying the connection between gut microbiota metabolites and immunotherapy, we may pioneer a new approach to adjuvant cancer treatment.
Affect intratumoral microbiome
The intratumoral microbiome constitutes a significant part of TME, influencing tumorigenesis, progression, metastasis, and antitumor immunity, 152 especially the extent of immune infiltration and the degree of activation of CD8+ T cells,153–156 which are important factors affecting ICI efficacy. Intratumor microbiota can promote CD8+ T-cell activation and TH1 differentiation of CD4+ T cells by increasing differentiation of classical activated macrophages (M1) and decreasing myeloid-derived suppressor cells, ultimately increasing PD-1 expression and responsiveness to immunotherapy. 156 Nejman et al. 157 found a higher abundance of intratumoral Clostridium and a lower abundance of intratumoral Gardnerella vaginalis in melanoma patients responding to ICIs. 157
By comparing the microbiomes of stool samples, tumor specimens, and non-tumor adjacent normal tissues from PDAC patients, approximately 25% of the intratumoral microbiome was derived from the gut microbiome. When fecal microbiota from patients with advanced PDAC was transferred to the murine intestine, the bacterial composition of the intratumoral microbiome was significantly altered, and a small proportion of fecal bacteria from human donors could be detected. 153 In the Lewis lung cancer mouse model, exogenous Akk increases the abundance of Akk in the circulation or tumor tissue, affecting the commensal microbiome and reprogramming the tumor metabolic network. 158 These results demonstrate that the intestinal microbiome could modulate the intratumoral microbiome.
Targeting the gut microbiome may reverse the intratumoral immune tolerance environment. Mackenzie et al. found that in preclinical melanoma, probiotic L. reuteri translocates to, colonizes, and persists within tumors where it locally promotes antitumor T-cytotoxic 1 immunity via its released aryl hydrocarbon receptor (AhR) agonist and dietary tryptophan catabolite, indole-3-aldehyde, which acts through CD8+ T-cell-specific AhR signaling to promote IFN-γ production in a cAMP response element-binding protein-dependent manner and enhance ICIs. 159
T-cell cross-reactivity has been mentioned above as a possible mechanism of ICI enhancement by gut microbiota. Whereas a study in melanoma showed that intratumoral bacteria can enter melanoma cells, and that their peptides can be presented by the human leukocyte antigen class I and class II molecules of melanoma tumor cells and elicit immune reactivity, again providing the basis for T-cell cross-reactivity. 160
With the above studies, the authors demonstrated a new way for gut microbiota to modulate immunotherapy through bacteria translocation to gut-distal tumors, providing a rational basis for designing new combination therapeutic strategies.
Regulate inflammatory pathways
The cyclic GMP-AMP synthase (cGAS)/STING signaling pathway has become a key mediator of inflammation, with the capacity to sense and regulate cellular responses to microbes. Gut microbiota associated with a high-fiber diet improves ICI efficacy via a STING-type I IFN-dependent mechanism. 69 Treatment with live L. rhamnosus GG also triggered type I IFN production in DCs through cGAS/STING/TRAF Family Member Associated NF-κB Activator (TANK) binding kinase 1/IFN regulatory factor 7 axis, enhancing the cross-priming of antitumor CD8+ T cells. 97 IL-12 is a heterodimeric pro-inflammatory cytokine that induces IFN-γ production, facilitates TH1 cell differentiation, and coordinates innate resistance and adaptive immunity. Oral supplementation with A. muciniphila restored the efficacy of PD-1 blockade in an IL-12-dependent manner by increasing the recruitment of CCR9+CXCR3+CD4+ T lymphocytes into mouse tumor beds. 22
The inflammatory pathway is an important link in the activation of antitumor immunity. Except for the above, it can be widely detected among the pathways by which the gut microbiota modulates immunotherapy, especially the T-cell receptor signaling pathway. There are far more inflammatory pathways involved, pending further exploration.
Others
MYC, a pivotal signal molecule regulating colorectal cancer proliferation, migration, stemness, and growth, induces PD-L1 expression in tumors, 161 leading to immunosurveillance escape and nonresponse or resistance to ICIs. C. butyricum destabilizes V-Myc Myelocytomatosis Viral Oncogene Homolog (MYC) through enhanced ubiquitination and increases CD8+ T-cell infiltration, suggesting that the intestinal microbiota may promote ICI efficacy by modulating MYC regulatory pathways. 162
Intestinal E. rectale enhances anti-PD-1 efficacy by altering the metabolic environment, which depletes environmental l-serine and activates NK cells via the FBJ murine osteosarcoma viral oncogene homolog (FOS)/FOS-like antigen 2 (FOSL2) signaling pathway. 163
Some components of intestinal bacteria or their metabolites can drive tumorigenesis,15,164,165 induce an immunosuppressive microenvironment, 156 and cause therapeutic tolerance, 20 known as ‘unfavorable gut microbiota’, such as B. fragili, Fusobacterium nucleatum, and Ruminococcus gnavus. F. nucleatum-derived succinic acid suppressed the cGAS-IFN-β pathway, thereby inhibiting the antitumor response by restricting the trafficking of CD8+ T cells to the TME in vivo. Treatment with the antibiotic metronidazole reduced the abundance of F. nucleatum, decreasing serum succinate levels, and resensitizing the tumor to immunotherapy. 166 By altering the microbiota composition, the proportion of unfavorable bacteria can be reduced to some extent, indirectly impacting anticancer therapy.
Future perspectives
The evidence above highlights the potential effect of the gut microbiota on the immunotherapeutic response and supports future efforts to improve treatment efficacy through its modulation.
However, each of these strategies has important variables to be considered. For FMT, it is essential to find expedient preparation and effective delivery methods, and strict policies must guide donor selection. Standardizing commercially available commodities is the most urgent requirement for probiotic or bacterial consortium application, and self-administration of off-the-shelf probiotics for cancer treatment is not recommended. We also have to decide on the type, dose, and formulation based on bacterial colonization to achieve optimal outcomes while avoiding potentially reduced microbiome diversity. Maintaining patients’ dietary patterns over time is the main challenge for dietary interventions. Whether the diet is sufficient to modulate therapeutic efficacy needs to be further demonstrated.
Limitations of this review include the following points. Gut microbiota modulates immunotherapy through multiple mechanisms. We mainly focus on the relevant mechanisms with well-defined evidence and only briefly describe the possible mechanisms that have yet to be rigorously confirmed. Besides, the regulation of gut microbiota is a new approach to enhancing ICIs. Although many clinical studies are underway, the published data are scarce and of limited value for clinical guidance. Research on the biological characteristics, pharmacokinetics, and optimal delivery of the potential interventions could further guide clinical medication. In conclusion, future challenges and directions are as follows:
➢ We need to further explore the relationship between gut microbiota and tumorigenesis, immune function, and antitumor immune response and uncover the biological effects and modes of action of commensal bacteria to lay the foundation for clinical research.
➢ The discovery of reliable microbiome-related biomarkers for predicting outcomes and stratifying patients accordingly could help guide the application of coregulatory interventions for the gut microbiota. The characterization of favorable and unfavorable gut microbiota is crucial, and the identification of microbial characteristics associated with different tumors or different drugs can contribute to the evolution of precision medicine.
➢ We need to determine the optimal modalities to effectively induce and maintain the favorable gut microbiota that is beneficial for immunotherapy over time, as well as factors that disrupt the beneficial bacterial component, and implement appropriate preventive measures.
➢ The evaluation of bacterial metabolites that enhance antitumor immunity can help facilitate the invention of novel products and target new molecular pathways.
➢ Other microbiomes in the gut, such as fungi and viruses, together with bacteria, form an integral part of the microecosystem. However, their roles have been underestimated in current studies. We need to refine relevant studies to confirm their value.
By addressing the above, we expect to establish a convenient and cost-effective approach to clarify the distribution of the gut microbiota and create a reproducible pathway to manipulate the gut ecosystem in combination with immunotherapy to improve patient survival.
Supplemental Material
sj-docx-1-tam-10.1177_17588359231204854 – Supplemental material for Modulation of gut microbiota: a novel approach to enhancing the effects of immune checkpoint inhibitors
Supplemental material, sj-docx-1-tam-10.1177_17588359231204854 for Modulation of gut microbiota: a novel approach to enhancing the effects of immune checkpoint inhibitors by Jinglong Huang, Caifeng Gong and Aiping Zhou in Therapeutic Advances in Medical Oncology
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.