
Abstract
Background:
Programmed death-ligand 1 (PD-L1) may be induced by oncogenic signals or can be upregulated via interferon gamma (IFN-γ). We have explored whether the expression of IFNG, the gene encoding IFN-γ, is associated with clinical response to the immune checkpoint blockade in non-small cell lung cancer (NSCLC) and melanoma patients. The role of inflammation-associated transcription factors STAT3, IKBKE, STAT1 and other associated genes has also been examined.
Methods:
Total RNA from 17 NSCLC and 21 melanoma patients was analyzed by quantitative reverse transcription PCR. STAT3 and Rantes, YAP1 and CXCL5, DNMT1, RIG1 and TET1, EOMES, IFNG, PD-L1 and CTLA4, IKBKE and NFATC1 mRNA were examined. PD-L1 protein expression in tumor and immune cells and stromal infiltration of CD8+ T-cells were also evaluated. Progression-free survival and overall survival were estimated.
Results:
A total of 17 NSCLC patients received nivolumab and 21 melanoma patients received pembrolizumab. Progression-free survival with nivolumab was significantly longer in NSCLC patients with high versus low IFNG expression (5.1 months versus 2 months, p = 0.0124). Progression-free survival with pembrolizumab was significantly longer in melanoma patients with high versus low IFNG expression (5.0 months versus 1.9 months, p = 0.0099). Significantly longer overall survival was observed for melanoma patients with high versus low IFNG expression (not reached versus 10.2 months p = 0.0183). There was a trend for longer overall survival for NSCLC patients with high versus low IFNG expression.
Conclusions:
IFN-γ is an important marker for prediction of response to immune checkpoint blockade. Further research is warranted in order to validate whether IFNG is more accurate than PD-L1.
Introduction
The inhibition of T-cell inhibitory receptors, mainly the programmed death 1/programmed death-ligand 1 (PD1/PD-L1) and the cytotoxic T-lymphocyte-associated antigen 4 (CTLA4), has led to a paradigm shift in the treatment of cancer. Monoclonal antibodies that target these immune checkpoint receptors have demonstrated promising antitumor activity and have achieved regulatory approvals for the treatment of multiple types of tumors.1–13 However, responses to PD-1⁄PD-L1 inhibition range from 20% to 40%.5,14–17 Until now, PD-L1 protein testing by immunohistochemistry (IHC) is the commonly used biomarker for selecting patients for immune checkpoint blockade therapy, at least for lung cancer patients. The PD-L1 IHC 22C3 PharmDx kit (Dako North America, Carpinteria, CA, USA) is United States Food and Drug Administration (FDA)-approved as a companion diagnostic for pembrolizumab in non-small cell lung cancer (NSCLC). The PD-L1 28-8 PharmDx kit (Dako North America) and the PD-L1 SP142 Ventana test (Ventana Medical Systems Inc., Tucson, AZ, USA) are approved as complementary diagnostics for nivolumab and atezolizumab, respectively. The PD-L1 SP263 Ventana test (Ventana Medical Systems Inc.) is approved for durvalumab. 18 The overall mutation load that is linked to smoking in lung cancer, or mismatch repair deficiency in colon cancer, have also been correlated with response to the immune checkpoint blockade.19–23
PD-L1 expression may be induced by oncogenic signals or can be upregulated via interferon gamma (IFN-γ). Both CD8+ T-cells and IFN-γ are critical for antitumor immunity. 24 Tumor cells are able to abolish interferon signaling signaling and therefore avoid antigen presentation. 25 Melanoma cells that are resistant to the immune checkpoint blockade do not respond to IFN-γ treatment with expression and activation of signal transducer and activator of transcription 1 (STAT1), or expression of interferon regulatory factors (IRFs) and downstream interferon targets, like PD-L1 and major histocompatibility complex (MHC) class I. 25 This interferon ‘insensitivity’ can also occur through epigenetic silencing of interferon signaling components or increased expression of negative regulators. For instance, activation of STAT3 activates the DNA methyltransferase 1 (DNMT1), which methylates the promoters and, therefore, silences the expression of IRFs and human leukocyte antigen (HLA) molecules. Simultaneously, STAT3 inhibits the expression and activation of STAT1 and further reduces antigen presentation 26 (Figure 1).

Immune suppressive and immune stimulating signaling pathways explored in our study. Signal transducer and activator of transcription 3 (STAT3) activates the DNA methyltransferase 1 (DNMT1), which silences the expression of interferon regulatory factors (IRFs), HLA molecules and subunits of the immunoproteasome complex (PSMB8 and PSMB9). The immunoproteasome facilitates antigen presentation for CD8+ T-cell responses. Ten eleven translocation 1 (TET1) enhances the activity of DNMT inhibitors. TET1 activity is upregulated by ascorbic acid. STAT3 can also indirectly inhibit the expression and activation of STAT1 and further reduce antigen presentation. Retinoic acid inducible gene-1 (RIG1) is one of the three RNA-sensing proteins [the other two are toll-like receptor-3 (TLR3) and melanoma differentiation associated gene-5 (MDA-5)] that induces interferon type I responses. Key transcription factors involved in RIG1 signaling include IRF3, IRF7 and NF-κB. Yes-Associated Protein 1 (YAP1) through a CXCL5–CXCR2 signaling axis drives myeloid-derived suppressor cell (MDSC) recruitment. STAT3 regulates the expression of Rantes (regulated upon activation, normal T-cell expressed and presumably secreted), also known as CCL5 and contributes to the recruitment of MDSCs. STAT3 increases the expression of genes encoding the calcium-binding proinflammatory proteins S100A and S100B. S100A and B activate cell surface receptors like toll-like receptor 4 (TLR4). A key downstream signal effect of TLR4 is mediated by activation of nuclear factor-kappa B (NF-κB), which produces large amounts of inflammatory cytokines like IL-6 and eventually STAT3 activation. Activated STAT3 has been demonstrated to bind to multiple sites in the arginase I promoter. High levels of arginase activity suppress CD8+ T-cell function and mediate MDSCs immune suppressive action. The T-box transcription factor eomesodermin (EOMES) maintains exhausted CD8+ T-cells. The noncanonical IκB kinase family member IKBKE (also called IKKε and IKKi), induced by tobacco components, regulates immune response by phosphorylating and activating STAT1. IKBKE can also negatively regulate T-cell immune responses through phosphorylating the nuclear factor of activated T-cells c1 (NFATc1) which complexes with STAT3 and cooperates in transcriptional regulation.
Treatment with DNMT inhibitors can sensitize cells to immune checkpoint blockade by derepressing endogenous retroviral sequences that are usually epigenetically silenced. Endogenous retroviral sequences trigger the double-stranded RNA (dsRNA) sensing pathway that through the RNA-sensing proteins toll-like receptor-3 (TLR3), melanoma differentiation associated gene-5 (MDA-5) and retinoic acid inducible gene-1 (RIG1) induce interferon type I responses. Tumors with high expression levels of the viral defense set of genes are more likely to respond to immune checkpoint blockade.27–29 Ten eleven translocation 1 (TET1) that is activated by vitamin C enhances the activity of DNMT inhibitors and contributes to endogenous retroviruses upregulatation in the dsRNA form and the induction of viral defense pathways 30 (Figure 1).
We have shown that STAT3 and Yes-Associated Protein 1 (YAP1) are related with innate resistance to epidermal growth factor receptor (EGFR) inhibition in EGFR-mutant NSCLC. 31 In prostate cancer, a disease with de novo resistance to immune checkpoint blockade,32,33 YAP1 through a CXCL5-CXCR2 signaling axis drives myeloid-derived suppressor cell (MDSC) recruitment, which ultimately suppress T-cell activity 34 (Figure 1). Rantes (regulated upon activation, normal T-cell expressed and presumably secreted), also known as CCL5, is a chemokine that its production is dependent on STAT3 35 and contributes to the recruitment of MDSCs and regulatory T-cells (Tregs) 36 (Figure 1). The T-box transcription factor eomesodermin (EOMES) maintains exhausted CD8+ T-cells defined by the expression of PD-1 37 (Figure 1).
The noncanonical IκB kinase family member, IKBKE (also called IKKε and IKKi), induced by tobacco components, inflammatory factors and viral infections, regulates immune response by phosphorylating and activating IRF3, IRF7 and STAT1. 38 However, IKBKE is a double edged sword since it can also negatively regulate T-cell immune responses through phosphorylating the nuclear factor of activated T-cells c1 (NFATc1). 39 NFATc1 complexes with STAT3 and cooperates in transcriptional regulation 40 (Figure 1).
In an effort to discover biomarkers that can predict the outcome to immune checkpoint blockade, we have examined archival tumor samples of NSCLC and melanoma patients who were treated with nivolumab and pembrolizumab, respectively, and we have analyzed the potential influence on survival and response of the mRNA expression of several transcripts that directly or indirectly influence tumor immunogenicity (Figure 1). The influence of the protein expression of PD-L1 on tumor cell and immune cells and the stromal infiltration of CD8+ T-cells were additionally examined.
Methods
Clinical samples
Pretreatment tumor archival specimens from advanced NSCLC patients and metastatic melanoma patients were retrospectively collected from the the Institute of Oncology Rosell (IOR), Quiron Dexeus University Hospital, Barcelona, Spain and from Hospitals of the Spanish Melanoma Group (GEM). All samples were derived either from the primary tumor or a metastatic biopsy performed >3 months before anti-PD-1 therapy. All patients provided written informed Ethics Committee (EC)/Institutional Review Board/(IRB)-approved consent before study entry.
Gene expression analyses
All analyses were performed centrally at the Pangaea Oncology SA laboratory, Barcelona, Spain, an ISO 15189 accredited laboratory by the Spanish Accreditation Body (ENAC). RNA was isolated from formalin-fixed paraffin-embedded (FFPE) tissue specimens in accordance with a proprietary procedure (European patent number EP1945764-B1) as previously described. 41 Quantification of gene expression was performed using the ABI Prism 7900HT Sequence Detection System (Applied Biosystems) and calculated according to the comparative Ct method. The primer and probe sets for each gene were designed using Primer Express 3.0 Software (Applied Biosystems) according to their Ref Seq (http://www.ncbi.nlm.nih.gov/LocusLink) respectively. The sequences of the primers and probes used were as follows: STAT3 forward 5′-CACCTTCAGGATGTCCGGAA-3′, reverse 5′-ATCCTGGAGATTCTCTACCACTTTCA-3′, probe 6FAM 5′-AGAGTGCAGGATCTAGA-3′ MGB; Rantes forward 5′-CATCTGCCTCCCCATATTCCT-3′, reverse 5′-AGTGGGCGGGCAATGTAG-3′, probe 6FAM 5′-ACACCACACCCTGCTG-3′ MGB; YAP1 forward 5′-TTGGGAGATGGCAAAGACATC-3′, reverse 5′-GCCATGTTGTTGTCTGATCGA-3′, probe 6FAM 5′-TCAGAGATACTTCTTAAATCACA-3′ MGB; CXCL5 forward 5′-CGCCATAGGCCCACAGTG-3′, reverse 5′-ATTTCCTTCCCGTTCTTCAGG-3′, probe 6FAM 5′-AGGTGGAAGTGGTAGCCT-3′ MGB; TET1 forward 5′-AAACCATCTGTTGTTGTGCCTCT-3′, reverse 5′-TTTGGGCTTCTTTTCCCTCTG-3′, probe 6FAM 5′-GGAGGTTATAAAGGAAAAC-3′ MGB; EOMES forward 5′-AATAACATGCAGGGCAACAAAA-3′, reverse 5′-CTCATCCAGTGGGAACCAGT-3′, probe 6FAM 5′-ATGTTCACCCAGAGTCT-3′ MGB; IFNG (that encodes for IFN-γ) forward 5′-TTAGGCATTTTGAAGAATTGGAAA-3′, reverse 5′-GGAGACAATTTGGCTCTGCATT-3′, probe 6FAM 5′-AGGAGAGTGACAGAAAA-3′ MGB; PD-L1 forward 5′-AGCTATGGTGGTGCCGACTA-3′, reverse 5′-TTGATTTTGTTGTATGGGGCATT-3′, probe 6FAM 5′-AGCGAATTACTGTGAAAGT-3′ MGB; IKBKE forward 5′-TCAAGCTCTTTGCGGTGGA-3′, reverse 5′-TGGAGCAGTACTCCATCACCAGTA-3′, probe 6FAM 5′-GAGACGGGCGGAAG-3′ MGB; NFATC1 forward 5′-CTACGTCCTACATGAGCCCGA-3′, reverse 5′-AGCTCATACGGGCCTGAGTG-3′, probe 6FAM 5′-GCCCTGGACTGGCA-3′ MGB and β-actin (internal reference gene) forward 5′-TGAGCGCGGCTACAGCTT-3′ reverse 5′-TCCTTAATGTCACGCACGATTT-3′, probe 6 FAM 5’-ACCACCACGGCCGAGCGG-3’ TAMRA. Gene expression of DNMT1, RIG1 (DDX58) and CTLA4 was analysed with Hs00154749_m1, Hs01061436_m1 and Hs00175480_m1 (Applied Biosystems), respectively.
Immunohistochemistry
Four microns thick sections were cut from FFPE tissue specimens and IHC was performed using the following antibodies: anti-PD-L1 rabbit monoclonal antibody (SP142, Ventana Medical Systems, Inc.) and CONFIRM anti-CD8 rabbit monoclonal primary antibody (SP57, Ventana) on an automated staining platform (Benchmark ULTRA, Ventana) with antigen retrieval and antibody dilutions following manufacturer’s recommendations. In the negative control, the primary antibody was omitted. Human tonsil was used as a positive control.
Statistical analysis
The primary endpoint of the study was to examine the potential effects of gene mRNA expression levels on survival and response. Progression-free survival and overall survival were estimated by means of the Kaplan–Meier method and compared with a nonparametric log-rank test. Progression-free survival was calculated from the start of treatment with immune checkpoint blockade to the date of disease progression or death from any cause. Overall survival was recorded from the first day of treatment with immune checkpoint blockade until death or was censored on the date of the last follow-up consultation. In addition to analysing gene expression as a continuous variable, expression levels were divided into tertiles. A multivariate Cox proportional hazard regression model was applied with potential risk factors as covariates, obtaining hazard ratios (HR) and their 95% confidence interval (CI). Each analysis was performed with the use of a two-sided 5% significance level and a 95% CI. Association between biomarkers was assessed using a Pearson correlation analysis. The statistical analyses were performed using SAS version 9.4.
Results
Gene and protein expression analysis and correlations among the biomarkers examined
The clinical characteristics of the patients included in this study are shown in Tables 1 and 2. In archival samples of 17 NSCLC patients treated with nivolumab and 21 melanoma patients treated with pembrolizumab we conducted a comprehensive analysis of the mRNA levels of key genes (Figure 1): STAT3 and Rantes, YAP1 and CXCL5, DNMT1, RIG1 and TET1, EOMES, IFNG, PD-L1 and CTLA4, IKBKE and NFATC1. This analysis is among the most comprehensive molecular biomarker studies in immunotherapy treated patients that has been reported to date. Both NSCLC and melanoma patients were heavily pretreated, with >50% having received at least two previous lines of therapy for metastatic disease (Tables 1 and 2). Gene expression levels were grouped based on the tertiles (Q33, Q66) and divided into high (>Q66 and Q33–Q66) and low (<Q33). As presented in Table 3, some of the correlations among the biomarkers reconfirmed the biological rational behind our study. In the melanoma cohort, STAT1 was strongly and significantly correlated with IFNG (r = 0.80, p < 0.0001). In both cohorts of patients, IKBKE was correlated with NFATc1 (r = 0.81, p = 0.0005 and r = 0.60, p = 0.0093, for NSCLC and melanoma, respectively) and STAT3 with YAP1 (r = 0.80, p < 0.0001). IFNG was positively and significantly correlated with CTLA4 (both cohorts) and with STAT1 and IKBKE in the melanoma group of patients (Table 3). We also evaluated the PD-L1 IHC expression on tumor cells and immune cells as well as the stromal infiltration of CD8+ T-cells. Greater number of CD8+ T-cells was found in melanoma patients with high IFNG mRNA expression (p = 0.0341).
Characteristics and treatment response to immune checkpoint blockade for the 17 NSCLC patients treated with nivolumab.

ECOG PS, Eastern Cooperative Oncology Group performance status; IHC, immunohistochemistry; mOS, median overall survival; mPFS, median progression-free survival; NSCLC, non-small cell lung cancer; PD-L1, programmed death-ligand 1. *Large cell carcinoma (1), adenosquamous (1).
Characteristics and treatment response to immune checkpoint blockade for the 21 melanoma patients treated with pembrolizumab.

ECOG PS, Eastern Cooperative Oncology Group performance status; IHC, immunohistochemistry; LDH, lactate dehydrogenase; mOS, median overall survival; mPFS, median progression-free survival; NSCLC, non-small cell lung cancer; PD-L1, programmed death-ligand 1.
Statistically significant Pearson linear correlation coefficients among the biomarkers explored.

Survival analysis for the NSCLC patients treated with nivolumab
For the NSCLC patients treated with nivolumab, IFNG mRNA expression emerged as the only biomarker that significantly influenced treatment outcome. With a median follow up of 15.7 (95% CI 3.4–23.0) months, median progression-free survival was 2 months (95% CI 0.5–3.1) and 5.1 (95% CI 1.4–15.2) months for patients with low and high IFNG mRNA, respectively (p = 0.0124), [HR for disease progression, 6.66; 95% CI 1.20–36.79; p = 0.0297; Figure 2(a); Table 4]. Univariate analysis revealed that low IFNG mRNA expression was significantly associated with shorter progression-free survival, whereas none of the other biomarkers examined or the clinical parameters were related with progression-free survival (Table 4).
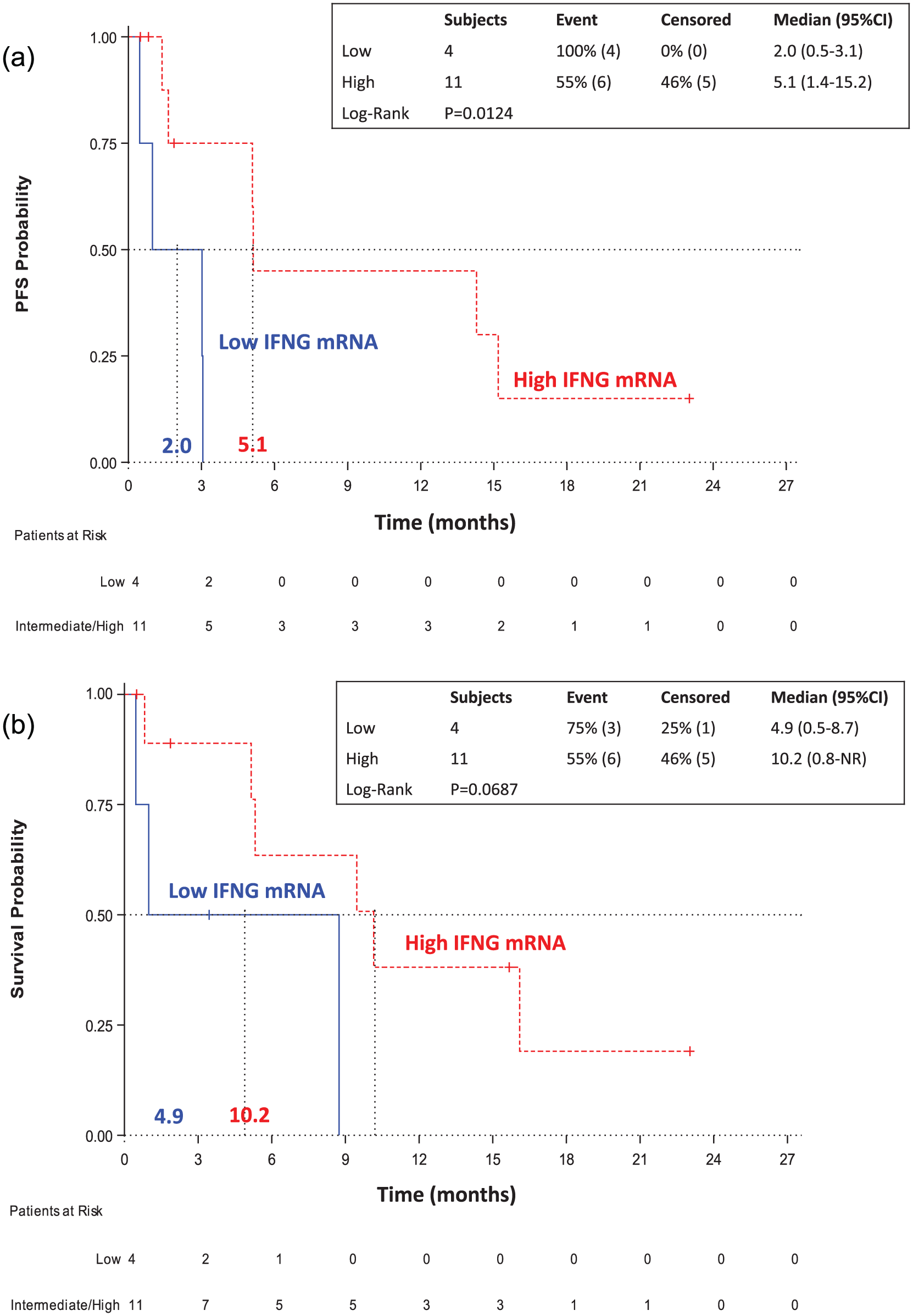
Survival analysis according to the expression levels of IFNG in NSCLC patients treated with nivolumab. (a) Kaplan–Meier curves for progression-free survival for NSCLC patients with low and high IFNG mRNA expression. (b) Kaplan–Meier curves for overall survival for NSCLC patients with low and high IFNG mRNA expression.
Univariate and multivariate analyses of clinicopathologic factors associated with PFS and OS in the cohort of NSCLC patients treated with nivolumab.

Chisq, Chi-square; CI, confidence interval; ECOG PS, Eastern Cooperative Oncology Group performance status; IHC, immunohistochemistry; NR, not reached; NSCLC, non-small cell lung cancer; OS, overall survival; PD-L1, programmed death-ligand 1; PFS, progression-free survival ; Pr>ChiSq, P value for the Wald Chi-Square test.
The statistical significant differences are highlighted in bold.
IFNG mRNA expression was not significantly associated with the survival of the patients but a clinically relevant difference was found. Specifically, median overall survival was 4.9 months (95% CI 0.5–8.7) and 10.2 [95% CI 0.8–NR (not reached)] months for patients with low and high IFNG mRNA, respectively (p = 0.0687), (HR for death, 4.10; 95% CI 0.80–20.83; p = 0.0911) [Figure 2(b); Table 4]. From the clinical parameters, the presence of brain metastases was significantly associated with shorter overall survival (HR for death, 6.12; 95% CI 1.22–31.03; p = 0.0286) (Table 4).
Survival analysis for the melanoma patients treated with pembrolizumab
Similarly to the NSCLC cohort of patients, IFNG was among the genes that most significantly influenced the outcome of the melanoma patients treated with pembrolizumab. With a median follow up of 12.4 (95% CI 7.3–18.0) months, median progression-free survival was 1.9 months (95% CI 0.0–5.1) and 5.0 (95% CI 1.5–14.1) months for patients with low and high IFNG mRNA, respectively (p = 0.0099), (HR for disease progression, 3.77; 95% CI 1.23–11.16; p = 0.0164) [Figure 3(a); Table 5]. Univariate analysis revealed that low IFNG mRNA expression and elevated lactate dehydrogenase (LDH) levels were significantly associated with shorter progression-free survival (Table 5). Paradoxically, low Rantes mRNA expression was also significantly associated with shorter progression-free survival. However, in the Cox’s regression model, only LDH levels remained a significant predictor of progression-free survival (HR for disease progression, 3.13; 95% CI 1.04–9.42; p = 0.0419; Table 5). There was a trend for IFNG levels to be associated with progression-free survival (HR for disease progression, 3.29; 95% CI 0.83–13.08; p = 0.0910).

Survival analysis according to the expression levels of IFNG in melanoma patients treated with pembrolizumab. (a) Kaplan–Meier curves for progression-free survival for NSCLC patients with low and high IFNG mRNA expression. (b) Kaplan–Meier curves for overall survival for NSCLC patients with low and high IFNG mRNA expression.
Univariate and multivariate analyses of clinicopathologic factors associated with PFS and OS in the cohort of melanoma patients treated with pembrolizumab.

Chisq, Chi-square; CI, confidence interval; ECOG PS, Eastern Cooperative Oncology Group performance status; IHC, immunohistochemistry; LDH, lactate dehydrogenase; NR, not reached; NSCLC, non-small cell lung cancer; OS, overall survival; PD-L1, programmed death-ligand 1; PFS, progression-free survival; Pr>ChiSq, P value for the Wald Chi-Square test. The statistical significant differences are highlighted in bold.
IFNG mRNA expression was significantly associated with the overall survival of the melanoma patients treated with pembrolizumab. Median overall survival was 3.1 months (95% CI 0.0–11.8) for patients with low IFNG mRNA while it was not reached (95% CI 2.6–NR) for patients with high IFNG mRNA expression (p = 0.0183), (HR for death, 3.50; 95% CI 1.16–10.60; p = 0.0265) [Figure 3(b); Table 5]. Median overall survival was 3.6 months (95% CI 0.03–5.4) for patients with low Rantes mRNA and 11.79 months (95% CI 2.6–NR) for patients with high Rantes mRNA expression (p = 0.0082), (HR for death, 4.75; 95% CI 1.34–16.86; p = 0.0159). From the clinical parameters, Eastern Cooperative Oncology Group (ECOG) performance status (PS) and LDH were associated with shorter overall survival (HRs for death, 3.34; 95% CI 1.01–18.99; p = 0.0473 and 6.40; 95% CI 1.89–21.70; p = 0.0029, respectively). In the multivariate analysis only LDH levels remained a significant predictor of overall survival (HR for disease progression, 5.45; 95% CI 1.28–23.28; p = 0.0221) (Table 5).
Response rate and IFNG mRNA expression
Considering that IFNG was consistently influencing the survival of both cohorts of patients, we then explored whether it may have an effect on the response to the immune checkpoint blockade. A total of nine NSCLC patients with IFNG expression levels determined were evaluable for response. Disease control rate was 71.43% for those with high IFNG expression (five out of seven patients with partial response) compared with 0% for patients with low IFNG expression (two out of two patients with disease progression) [Figure 4(a)]. A total of 19 melanoma patients with IFNG expression levels determined were evaluable for response. Disease control rate was 71.43% for those with high IFNG expression (out of 14 patients, 8 had stable disease, 1 had partial response and 1 had complete response) compared with 20% for patients with low IFNG expression (1 out of 5 patients had stable disease) [Figure 4(b)]. The above results should be taken cautiously considering the low number of patients being evaluated.

Best response according to IFNG expression. (a) NSCLC cohort. (b) Melanoma cohort.
Discussion
IFN-γ is a cytokine that was initially discovered as crucial for the host response to viral infections and recently has been recognized as having a key role in cancer related immunity. 42 Defects in the interferon signaling pathway are one of the main mechanisms of resistance to immune checkpoint blockade.25,43 IFN-γ secreted by immune cells in the tumor microenvironment causes growth arrest, augments MHC class I expression, contributes to the recruitment of effector cells, causes T-reg fragility and coordinates the process of innate and adaptive antitumor response.25,44–46 At the same time, the same IFN-γ signaling compromises antitumor immunity and activates PD-1 activity. 47 IFN-γ induces the expression of PD-L1, through increasing STAT1 signaling and decreasing STAT3 activation. 48 IFN-γ can also upregulate the expression of other immune suppressive molecules. 49 In our study we found a strong positive correlation of IFNG with STAT1 mRNA expression, while in both cohorts, a strong correlation was detected between IFNG and the immune suppressive molecule CTLA4. NSCLC and melanoma patients with high mRNA expression of IFNG exhibited longer progression-free and overall survival and higher disease control rates with anti-PD-1 therapies, nivolumab and pembrolizumab.
PD-L1 expression on tumor cells has been extensively explored as a biomarker to identify patients more likely to respond to immunotherapy. Response rates to immune checkpoint blockade range from 36% to 100% for PD-L1 positive tumors, while patients whose tumors do not express PD-L1 can experience a response rate ranging from 0% to 17%. 50 Although PD-L1 has been assessed within most of the pivotal studies as a predictive biomarker for immunotherapy, it is still unclear whether PD-L1 expression correlates with treatment outcomes. There are several caveats with the use of PD-L1 expression as a biomarker that indeed can be applicable only to patients who are treated with anti-PD-1⁄PD-L1 antibodies and no other types of immunotherapy. The first and most important problem is the use of various PD-L1 detection methods, like IHC, flow cytometry, mRNA expression that does not allow the standardization of the technique. Then, it is still not clear what levels of PD-L1 expression define positivity, while PD-L1 expression can occur, not only on the tumor cells, but also on the nonmalignant cells of the tumor microenvironment. 51 PD-L1 is an inducible and highly dynamic receptor that can change over time. 52 Finally, differential PD-L1 expression may occur according to tumor type or histological subtype. 52 The Blueprint PD-L1 IHC Assay Comparison Project compared four PD-L1 IHC assays (22C3, 28-8, SP142, and SP263) and found that the SP142 is inferior in comparison with the rest. 53 A previous study found high concordance among six PD-L1 monoclonal antibodies, including SP142. 54 However when Rimm and colleagues compared in 90 archival NSCLCs four PD-L1 platforms (22C3, 28-8, SP142, and E1L3N), they found that SP142 is an outlier that can detect less PD-L1 expression in comparison with the rest of the assays. 55 We have found that IFNG mRNA expression can be predictive of response with >70% of NSCLC or melanoma patients achieving response or stabilization of their disease. PD-L1 expression neither by protein (SP142 Ventana test) nor by mRNA expression was associated with treatment outcome.
High tumor mutational burden can predict favorable outcome to the immune checkpoint blockade. 56 But still, the presence of high mutational loads does not always correlate with responses. Is there any interaction between IFN-γ signaling and tumor mutational load? The downstream interferon target, transporter associated with antigen processing 1 (TAP1), 25 permits the entrance of short peptides, produced by the ubiquitylation and proteasome degradation of mutant proteins, into the endoplasmic reticulum. 57 In the endoplasmic reticulum the peptides bind to MHC class I, and then are transferred to the plasma membrane where they can be recognized by CD8+ T-cells. 57 This indicates that high tumor mutational burden requires an active interferon signaling for the post-proteosomal trimming of antigen epitopes, 58 in order to induce immune mediated tumor elimination.
In our study, paradoxically, high expression of Rantes (CCL5) was related with longer progression-free and overall survival for the melanoma patients treated with pembrolizumab, although in the multivariate analysis, only LDH remained significant modulator of treatment outcome. This finding can be due to the dual role of Rantes. On one side Rantes is a chemokine that contributes to the recruitment of MDSCs and Tregs 36 and on the other side its production by dendritic cells and macrophages, under the effect of IFN-γ, contributes to the recruitment of CD8+ T-cells. 59 None of the rest of the biomarkers explored in our study was associated with treatment outcome.
Our findings reinforce previous knowledge on the fact that response to immune checkpoint blockade occurs mainly in patients with a preexisting intratumoral T-cell adaptive immune response.49,60–63 Antoni Ribas was among the first to show that an IFN-γ signature is associated with better survival for pembrolizumab treated melanoma patients. 64 Several efforts are subsequently ongoing to identify essential genes in cancer cells that can facilitate immune selection. Applying an 18-gene IFN-γ signature with the Nanostring nCounter platform in RNA isolated from FFPE tissue, Ayers and colleagues were able to separate responders from nonresponders to pembrolizumab, across multiple solid tumors. 65 The signature consisted of the following genes: TIGIT, CD27, CD8A, PD-L2, LAG3, PD-L1, CXCR6, CMKLR1, NKG7, CCL5, PSMB10, IDO1, CXCL9, HLA.DQA1, CD276, STAT1, HLA.DRB1 and HLA. Their signature performed favorably in comparison with PD-L1 IHC (22C3 PharmDx kit). 65 In the POPLAR study, comparing atezolizumab versus docetaxel for previously treated NSCLC patients, those with high T-effector-IFNγ-associated gene expression had improved overall survival with atezolizumab. 7 High levels of a baseline IFNG gene expression signature was associated with greater benefit from durvalumab in NSCLC patients. 66 Prat and colleagues, with the Nanostring nCounter platform, reported a 23-gene signature related to T-cell and IFNγ that could predict response and progression-free survival to nivolumab and pembrolizumab. 67 Loss-of-function mutations in APLNR, encoding the apelin receptor, have been found in patient tumors that were refractory to immunotherapy. 68 APLNR interacts with JAK1 to augment IFN-γ response. 68 In pancreatic tumor models that lack IFN-γ, CKLF-like MARVEL transmembrane domain containing protein 6 (CMTM6) has been described as a regulator of PD-L1 expression.69,70
Our study has some limitations. First, it is a retrospective study with a limited sample size. Second, we used the SP142 assay for PD-L1 IHC expression, which now is considered less consistent in PD-L1 tumor cell staining in comparison with the Dako 22C3, Dako 28-8, and VENTANA SP263 assays. In our study, approximately 20% of the patients in both cohorts (NSCLC and melanoma) were stained positive for PD-L1 expression. Still, PD-L1 expression by both IHC and mRNA were not predictive of treatment outcome. Finally, the patients included in our study were heavily pretreated and biomarker assessment was performed for most of the cases in the diagnostic sample that could be far from the immune checkpoint blockade therapy initiation.
In summary, our results show that single mRNA expression of IFNG could be predictive of response and survival to immune checkpoint blockade. These results are consistent with previous findings and the concept that the presence of a preexisting adaptive immune environment predicts clinical outcome. Ongoing clinical studies are evaluating immune-related gene expression profiles, but we also consider that single IFNG mRNA expression warrants further clinical validation. In our molecular oncology laboratory, Pangaea Oncology, we are screening our patients for IFNG mRNA expression and we are in the process of validating a 7-gene IFN-γ signature (IFNG, CD274, CD4, CD8A, FOXP3, PDCD1 and GZMM) with the Nanostring nCounter platform 71 as a predictor of response to immunotherapy.
Footnotes
Acknowledgements
The initial results of this study were presented in an oral presentation at the American Society of Clinical Oncology (ASCO) meeting in 2017.
NK, MGC, GC and RR designed the study, did the literature search, and wrote the manuscript; NK, EA, AJC and CT performed the experiments; NK, MGC, GC, AD and RR and RR analyzed and interpreted data; NK, MGC, GC, SV, MDLLG, SMA, EPR, IMR, DRA, RB, TP, MAR, and RR collected data; all authors contributed to the writing, review, and approval of the final manuscript
Funding
This work was funded by La Caixa Foundation, Barcelona, Spain.
Conflict of interest statement
The authors declare that there is no conflict of interest.