
Abstract
Understanding habitat selection patterns in congeneric species has raised special interest due to its importance when designing conservation strategies for the species. We analysed reproductive habitat selection of three coexisting warbler species in the eastern Iberian Peninsula. We detected the individuals by their vocalizations at 40 listening points in two study areas (Carcaixent and Alcoy). We used satellite images to identify different vegetation types and their availability. Reproductive habitat selection was analysed by the Ivlev selectivity index and a compositional analysis. In addition, we determined the effect of different habitat variables (dominant vegetation, vegetation height, cover and site) on the presence of the species fitting a binomial GLM with logistic linking function. We found that the Sardinian Warbler (Curruca Melanocephala) selected scrub-dominated areas; while the Dartford Warbler (C. undata) preferred areas with low (<50%) vegetation cover; and the Subalpine Warbler (C. cantillans) was present in areas with scrubs and low (<50%) vegetation cover, avoiding crops. The compositional analysis showed that the species used different vegetation types, in relation to their availability and that the selection differed between species. The Sardinian Warbler was the only species which showed spatial variation in reproductive habitat selection. Our results evidence that the studied warbler species select and avoid different types of vegetation in relation to their specific requirements and the present habitat structure. This represents valuable information to understand the composition of local communities and the reproductive habitat selection of the studied species.
Introduction
Habitat selection is defined as the ability to choose the environment in which to live. 1 From the moment when the environment is elected, it turns into a hierarchical process where the animal has to make a series of innate and learned behavioural decisions on different levels of the environment. 2 This is a process in which the species choose among the different available resources, following their preferences. 3 The habitat selection of a species can be influenced by a diversity of factors such as the amount and quality of environments or landscape elements, the phenotypical plasticity of individuals, the population structure or the social organization of the species present in a certain habitat.4–11
In passerine birds, the mechanisms of coexistence differ according to the life cycle of each species. Interspecific differences in habitat use and selection act on a spatial and temporal scale. 12 In this case, the segregation may be a sub-product of resource partition, especially if the preferred resources are distributed unequally among the different habitats.13,14
Curruca is a genus of the Sylviidae family, which represents a relatively large group of insectivorous passerines. 15 These species can be found in a wide spectrum of habitats, from desert scrub to deciduous and temperate forests. In the Mediterranean region, these species have a high level of sympatry, finding up to seven species of the genus on a regional scale, or up to five species in a certain habitat type. 16
Previous studies on the coexistence of different warbler species explain this fact by an ecological segregation of the habitat and nesting sites.17,18 Furthermore, the importance of the interaction between congeneric and non-congeneric species in the dispersion and habitat selection was also studied.6,7,10 In addition, differences in habitat selection between migrant and sedentary species were also evaluated.5,9,11 Other studies on the habitat selection dynamics of warblers considered factors such as anthropic disturbance, 19 seasonality 20 and social behaviour. 21 Finally, Coreau & Martin 22 performed a multiscale analysis in order to improve the knowledge about bird distribution and changes in their habitat selection.
The Subalpine Warbler (Curruca cantillans) which is a migratory species breeding in the area, in the Iberian Peninsula seems to select Mediterranean scrub habitats, regardless of height or level of coverage.20,23 This species is abundant in dense evergreen oak forests, 24 remaining in the upper layers of vegetation, but also in areas with a high density of trees. 7 The Sardinian Warbler (C. melanocephala) is a species resident in Spain and probably one of the most common warblers in the Mediterranean, it is present in all types of habitats at low altitudes.20,25 Most studies show its close relationship with the Mediterranean scrubland,26–28 habitats with different degrees of structural complexity, 18 areas with agricultural use and urban areas. 11 Finally, the Dartford Warbler (Curruca undata), resident in the Iberian Peninsula, is linked to bushes or dense, homogeneous scrub,29,30 while urban areas have a negative effect on the species.29,30 Therefore, we expect/hypothesize differences in breeding habitat type selection between the species, especially between the Sardinian Warbler (generalist) and the Dartford Warbler (specialist), and that this selection changes with the presence of the Subalpine Warbler.
Thus, the aim of this study is to analyse the reproductive vegetation selection of three sympatric species of Mediterranean Warblers (Dartford Warbler, Sardinian Warbler, and Subalpine Warbler) by defining the spatial distribution of habitat type of these three species at a territory scale. To do so, we determined if the selection of the vegetation is linked to specific requirements (height, dominant vegetation and cover) in relation to the plant structure of the site, and identified if the habitat selection made by resident species is maintained regardless of the presence of the migratory species, since Ieronymidou et al. 9 found evidence of niche change between species of the same genus.
Methods
Study area
The study was carried out in the east of the Iberian Peninsula (Valencian Community), in two sites within the municipalities of Alcoy (Alicante) and Carcaixent (Valencia) (Figure 1). The first site is located east of Carrascal Natural Park of Font Roja, Alcoy (38° 40' 15" N, 00° 28' 36" W), the park has an area of 115 ha (11.6 ha sampled area) and situated 912 m a. s. l. This park is one of the natural enclaves of the Valencian Community where the Mediterranean forest is still present. Its altitude ranges between 600 and 1365 m a. s. l. The mean annual temperatures in 2014 and 2015 were 19.0°C and 19.1°C, respectively (available at https://www.avamet.org, accessed 16 of February 2021). The mean annual precipitation can reach up to 843 mm/year, with a strong peak in October, in which more than a half of the total precipitation is commonly recorded as torrential falls.
31
There are different vegetation formations, mainly Pine (Pinus halepensis) forest and Oak (Quercus ilex) forest, forming a dense and continuous mantl.
32
Location of the study areas in Spain. (A) Font Roja Evergreen Oak Forest Natural Park, Alcoy. (B) Municipal Natural Park of the Font de la Parra-Hort de Soriano, Carcaixent. Type of vegetation: open cropland (oC), dense cropland (dC), open scrubland (oS), open pine-scrubland (oPS), dense pine-scrubland (dPS), open scrubland-pinewood (oSP), dense scrubland-pinewood (dSP).
The second site is situated in the Font de la Parra-Hort de Soriano Municipal Natural Park, Carcaixent (39° 04' 10'' N, 00° 24' 36" W), the park has an area of about 113 ha (11.3 ha sampled area) and situated 112 m a. s. l. The mean annual precipitation in the area oscillates between 400 and 700 mm/year, mainly raining in spring and fall, with a mean annual temperature around 17°C. 33 The vegetation is conditioned by the occurrence of numerous forest fires, leading to a predominance of thermomediterranean scrub formations with Kermes Oak (Quercus coccifera) and Mastic Tree (Pistacia lentiscus), accompanied by labiate formations, dominated by Rosemary (Rosmarinus officinalis) and Thyme (Thymus vulgaris). In addition, there are large areas of mainly orange crops (Citrus x sinensis). 34
Fieldwork
Bird sampling
Bird censuses were carried out in spring (April – May) of 2014 and 2015. We did a total of 16 field trips, eight per study area and four per year. Data were collected using the point count method. 35 Prior to sampling, 40 points near accessible pathways were randomly selected (20 per area) (Figure 1). Each point was located with a minimum distance of 200 m from other points, in order to guarantee the independence of the surveys and to avoid an overestimation of the individuals. 36 The surveys of each group of 20 points began 30 minutes after sunrise, and were carried out during the following 4 hours. To avoid biases we varied the order in which the points were sampled. At each point, one researcher (EAMM) listened for five minutes and registered all detected individuals within a 100 m radius. This method is recommended when the interest of the study focuses on comparing the relative differences of species between habitats during the reproductive season. 36 Also, this methodology is mainly based on auditory cues, which is very useful for shy species or dense habitats, like the study area. During the surveys, the orientation and the distance of the birds from the observer were measured using a Silva Sight compass and a 10x25 laser range finder (Bushnell). The geographic location of each sampling point was registered using Google Maps and we inferred the location of each bird by triangulation using the coordinates of the sampling point, the measured bird orientation and distance.
In other studies, it has been observed that in adjacent territories males of the genus seem to sing mainly at the border of their territory close to the territory of the neighbouring male. 37 Thus, in the data analysis, all detected songs and calls (i.e. territorial and non-territorial vocalizations) were classified by species and, if in different samples two vocalizations were located within a radius of 50 meters (the maximum distance between nesting areas of the species in natural habitats), they were assumed as the same individual. 16
Vegetation characterization
Description of the sampled vegetation types.

Data analyses
We determined the effect of vegetation characteristics on the presence of species fitting Generalized Linear Models (GLM) with binomial error distribution and logit link function. 39 We considered the presence (1) and absence (0) of each species in each of the 40 sampling points during the entire duration of the study as a dependent variable. As independent variables we used the site (Alcoy or Carcaixent), the dominant vegetation type at the sampling point (pine, scrub or crops), the cover (above or below 50%), height of the vegetation (above or below 1.5 m) and the presence of the other two species. To analyse which vegetation type differs significantly, we carried out a pot-hoc Tukey test using the “multcomp” R package. 40
Afterwards, we drew an area of 0.5 ha around each detected individual using QGIS v2.1 38 in order to study the habitat selection of each species. We chose this size because it is half of the average maximum territory (1 ha) reported for these species during the breeding season.7,16,41 Our study is similar to those of the household range, where the same individual is recorded several times in many samples, but not in the same one.7,41 Following previous studies, 42 where the authors applied a similar methodology, the data from both years were combined for the analysis since the effect of lack of independence should be insignificant. Especially considering that only approximately half of the adult birds survive between successive years, which also decreases the effect of dependence.42,43 To analyse if warblers exhibit differences in habitat selectivity at the territorial level, we used the Ivlev's selectivity index: 44 Ivlev = (UI-DI) / (UI + DI), where UI = ui / u+*100 and DI = di / d+*100, being ui the number of contacts of the species that used the vegetation type i and u+ the total number of contacts of the species observed in the vegetation type studied, and di, the availability in the environment of vegetation type i, and d+, the total availability of vegetations types. This index oscillates between -1 and +1; according to Atienza, 45 values < -0.5 indicate a negative selection, values > 0.5 indicate a positive selection, while values between -0.5 and 0.5 indicate no selection at all.
We determined if the use made by the species of the vegetation type is dependent on its availability with a compositional analysis of habitat use, using the adehabitat package. 46 The number of repetitions in the random assigning tests was 500. We replaced the values of 0 (which represented the absence of individuals in the vegetation types) by 0.01, following Aebischer et al. 47 We built the usage matrix with the proportion of contacts of each species recorded in each habitat type during the surveys; and the vegetation matrix, with the proportion of the area covered by each type of vegetation. We determined the existence of significant differences in the use of the vegetation performing a Wilks Lambda test and built a use frequency rank matrix for each study area.
All statistical analyses were carried out using the statistics software R. 48
Results
Mean ± SD of the number of detected birds for each sampling site per species; and the percentage of sampling points where the species were detected, grouped by the dominant vegetation type, vegetation height, cover and site.
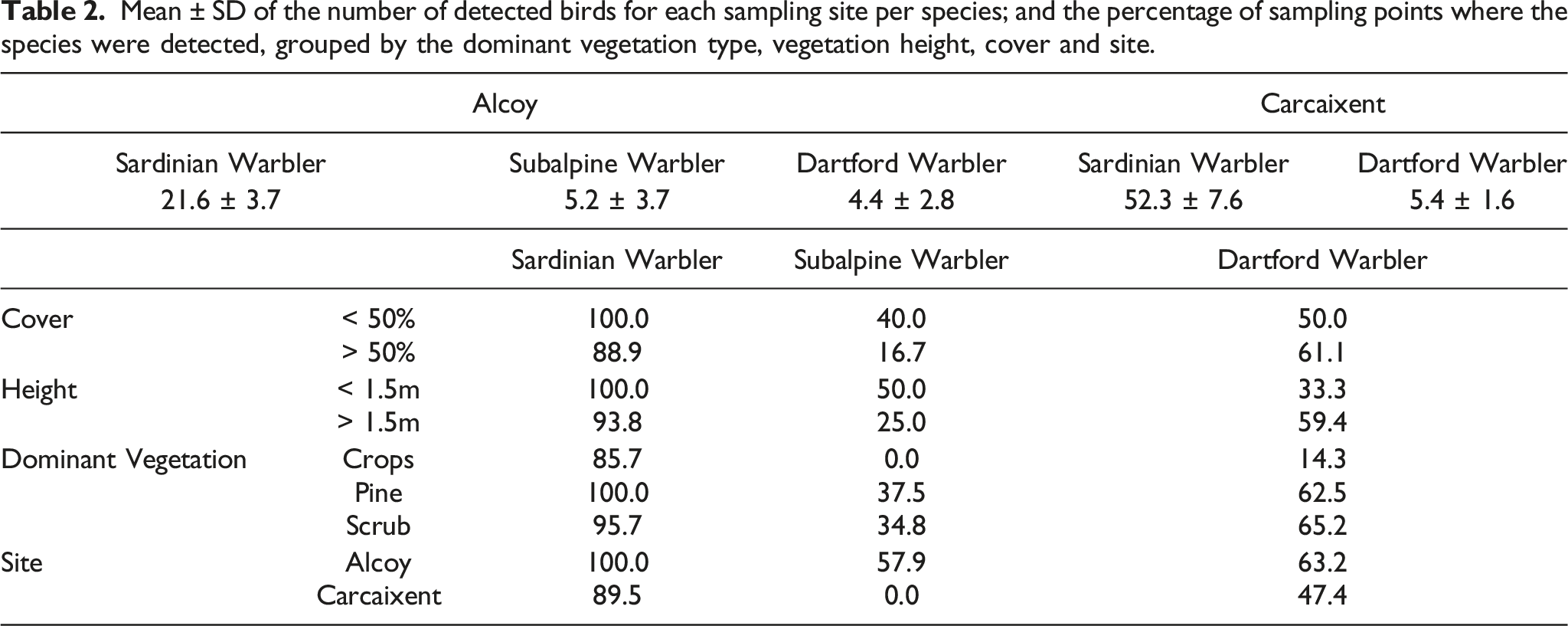
Results of the GLM for each predictor in each species, together with the result of the post hoc Tukey test for the Dominant Vegetation variable: Estimate (Est), Standard Error (SE) and t (GLM), z (Tukey) and P statistics. Significant (P < 0.05) results are shown in bold.

Ivlev's selectivity index showed that the Sardinian Warbler did not select or avoid any of the different vegetation types present in the studied sites (Figure 2). The Dartford Warbler positively selected oS (0.64), and avoided dC (-0.89) in Carcaixent, while in Alcoy the habitat selection of the species was negative. (Figure 2). Finally, the Subalpine Warbler was only registered in Alcoy and negatively selected the oPS (-0.62) (Figure 2). Ivlev’s index of selection for different vegetation types by three species of warblers in two study sites (Carcaixent and Alcoy): Sardinian Warbler (black), Subalpine Warbler (grey) and Dartford Warbler (white).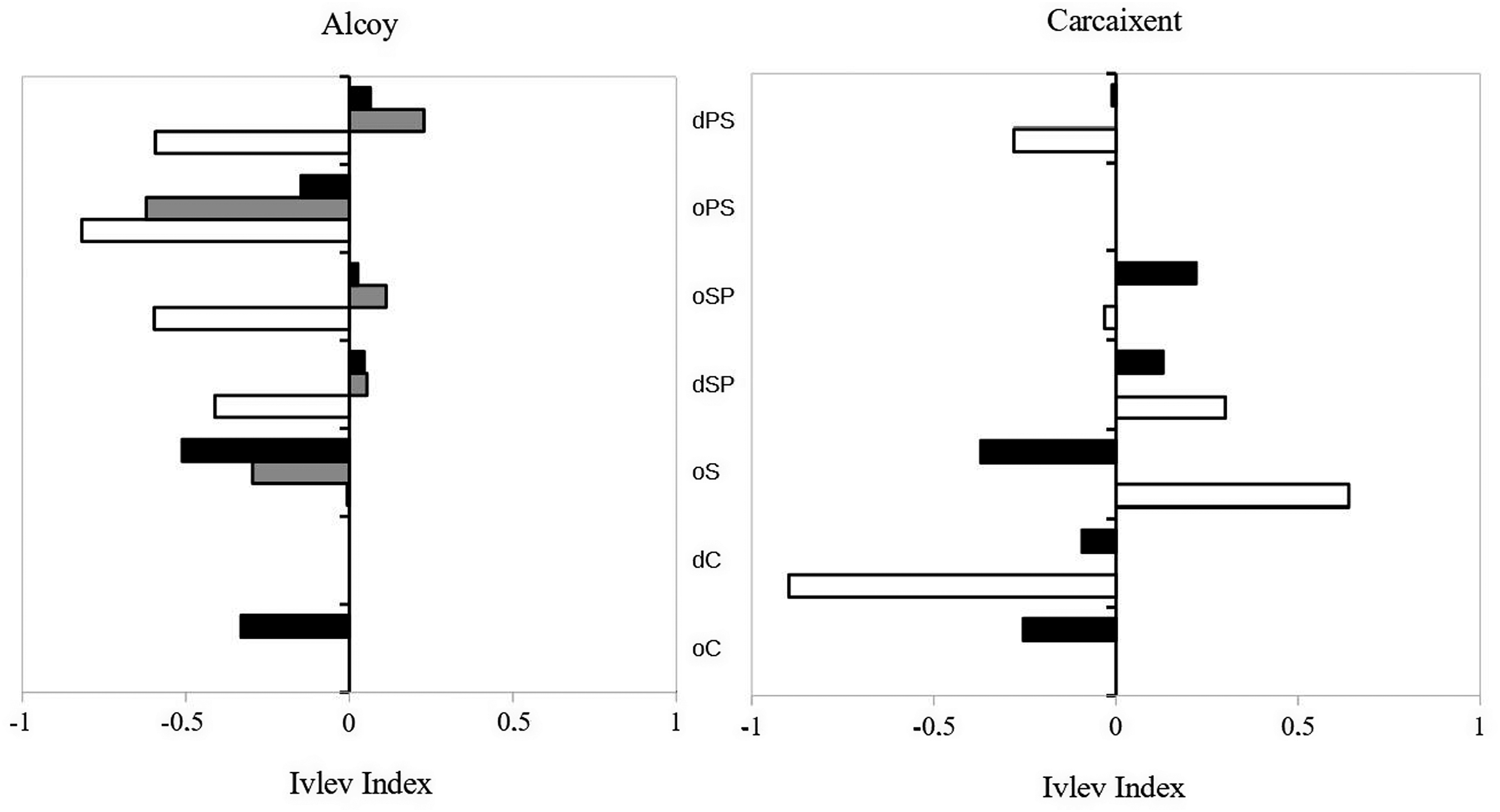
Simplified classification matrix for each species, comparing the proportion of observed individuals in each type of vegetation, with the proportion of the sampled vegetation in Alcoy and Carcaixent. Symbols indicate preference (+) or avoidance (−) of the row habitat in relation to the column habitat, the triple symbol indicating significance (P < 0.05). Rank is equal to the sum of the positive values in each row. Higher ranks indicate a more preferred habitat.

Discussion
We did not observe a general vegetation selection pattern between species on a territorial scale. Possibly, as proposed by Coreau & Martin, 22 the spatial distribution of the studied warblers can be explained by the interaction between two or more habitat variables. Other studies suggest differences in habitat use between ecologically similar migratory tanagers in the same habitat. 21 On an ecological scale, species can differ in the chosen vegetation level and feeding strategies.21,49 So that their coexistence in reproductive territories can be favoured by the spatial partition on a microhabitat scale. It is also important to consider that reproductive birds can use strategies in opposition to the habitat selection as a response mechanism to the spatio-temporal variation of selective pressures.50–53 In some bird species, different life history traits and environmental factors are involved in setting the moment of reproduction, determining the reproductive habitat use of those species. 54
Territorial exclusion is typical of non- or semi-colonial species, in areas where defending the resources is very important. 41 As demonstrated by other authors,9,55 the maximum density of the studied warbler species was observed in a specific vegetation within the gradient of available local and territorial habitats, a result supporting the hypothesis of ecological segregation which was already described for this group.4,6,16,18,20,22,56 In the particular case of the studied species and the considered scale, we assume that the most used sites present optimal conditions for the reproduction, i.e. being occupied first, evidencing the selection priority.57–59 In our study, we did not include food availability variables to explain the selection, however, Bas et al. 41 showed that, in the case of warblers, the reason for grouping in specific areas could be the abundance of resources in these areas in a period when resources in general are scarce. This behaviour could explain that the habitat selection slightly differed among the species, something also described by Ieronymidou et al. 9
The Sardinian Warbler showed a higher relation to a scrub dominated vegetation, what can be related to the higher availability of this vegetation structure within the studied areas. In accordance with the distribution of the species observed in other studies,22,55 our results corroborate the extended distribution of the species. During the reproductive period, the constant search for feeding grounds and the risk of predation41,60,61 may lead the individuals to select more natural habitats, 20 where the productivity is higher and hiding opportunities are more abundant. In general, the scrub type habitat is preferred by the warbler species for reproduction. 53 This type of cover may be necessary for their reproductive success, suggesting a tight relation of locally available resources and the body condition of females before the reproduction. 62
The considered habitat variables did not show any relation to the habitat selection in the case of the Dartford Warbler. In other studies it was observed that parcel composition and landscape variables, like the number of rocky parches and the area of scrub dominated parches, describe the distribution of the species better at a multiscale level. 22 Several studies focus on describing the vegetation type preferred by the species,17,20,30,63 showing its clear preference for a certain vegetation structure (scrub). The dynamic behind the formation of this type of vegetation structure is interesting, as it includes climatic factors and landscape dynamics (fires), which, in consequence, determine warbler colonization and habitat selection. 64 Our results suggest that the Dartford Warbler can select different areas, depending on their availability, however, this species shows a higher use in habitats with scrub presence, being it disperse or dense. We also have to remark that the data collection in our case was not focused on determining the effect of habitat selection on the reproductive success of the species, an important aspect to infer the future of the population in this region. It would be necessary to consider other microhabitat related variables, which are correlated on the landscape level and which allow to explain with more clarity the observed selection patterns of the Dartford Warbler in our study.
The Subalpine Warbler is a migratory species which also related to areas dominated by scrub. In some individual migratory birds it was observed a certain disregard in the selection of habitat characteristics during the migratory periods, which suggests a higher plasticity during the migration. 65 The habitat selection of the Subalpine Warbler in our study was consistent with its preference for heterogeneous scrub formations which include small trees or large scrubs which the birds use for feeding or singing, and small open patches embedded in a medium scrub formation, used mainly as feeding grounds.16,17,66 Coreau & Martin 22 indicate that the distribution of the Subalpine Warbler on a landscape level is related to the local vegetation composition, mainly the presence of numerous small patches of grass. In the case of migrant birds, it was also evidenced that individuals consider social information offered by other individuals when selecting their habitat.21,67–69 Our results do not support the hypothesis of congener incidence on the selection, as we did not observe a significant effect of the presence of other warbler species on the presence of a certain warbler species. This may be in discord with Bensusan et al., 69 where the authors observed a change in the habitat use migrant populations of the Sardinian Warbler in Gibraltar.
In conclusion, our results indicate that the behaviour of habitat selection in warblers is dependent on the ecological characteristics of the species and the environment and their interaction on different spatial levels. Furthermore, regional changes in habitat selection suggest a relation to regional differences in landscape composition.
Footnotes
Authors contribution
Eyda A. Moreno-Mosquera: Study conception, Methodology, Data collection. Robby M. Drechsler: Writing and manuscript preparation. Silvia Rivera: Data collection. Juan S. Monrós: Supervision
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The main author thanks the Administrative Department of Science, Technology and Innovation (Departamento Administrativo de Ciencia, Tecnología e Innovación – COLCIENCIAS) of Colombia for the predoctoral grant which allowed us to carry out this study.