
Abstract
Experimental in vitro and in vivo studies support the hypothesis that heat-treated, lyophilized Lactobacillus acidophilus LB cells and concentrated, neutralized spent culture medium conserve the variety of pharmacological, antimicrobial activities of the live probiotic strain against several infectious agents involved in well-established acute and persistent watery diarrhoea and gastritis. Heat-treated cells and heat-stable secreted molecules trigger multiple strain-specific activities explaining the therapeutic efficacy of L. acidophilus LB. This review discusses the current body of knowledge on the antimicrobial mechanisms of action exerted by L. acidophilus LB demonstrated in in vitro and in vivo experimental studies, and the evidence for the therapeutic efficacy of this anti-infectious biotherapeutic agent proved in randomized clinical trials for the treatment of acute and persistent watery diarrhoea associated with several intestinal infectious diseases in humans.
Introduction
The patented Lactobacillus acidophilus strain LB (L. acidophilus LB) [Servin et al. 1999; Chauviere et al. 2005; Servin et al. 2006] is the major component of a gastrointestinal anti-infectious biotherapeutic pharmaceutical product consisting of a combination of lyophilized, heat-treated Lactobacillus LB cells and concentrated, neutralized spent culture medium. The heat-treated L. acidophilus LB cannot be considered to be probiotic [Sanders et al. 2007], but is of interest since the pharmacological activities of the parental probiotic live strain are conserved.
Anti-infectious activities of human intestinal microbiota Lactobacillus
At the beginning of the 20th century, Elie Metchnikoff, the Russian-born Nobel Prize winner who worked at the Pasteur Institute in Paris, reported the first observation of the positive role played by some selected bacteria from the human intestinal flora and suggested that: ‘The dependence of the intestinal microbes on food makes it possible to adopt measures to modify the flora in our bodies and to replace the harmful microbes by useful microbes’ [Metchnikoff, 1907].
In 2001, an expert consultation of international scientists working on behalf of the Food and Agriculture Organization of the United Nations (FAO) and the World Health Organization (WHO) defined probiotics as follows: ‘live microorganisms which, when administered in adequate amounts, confer a health benefit on the host’ [FAO/WHO, 2001, 2002]. This definition has become the most widely adopted and accepted version worldwide, and a consensus meeting of the International Scientific Association for Probiotics and Prebiotics (ISAPP) recently refined the appropriate use and scope of the term ‘probiotic’ [Hill et al. 2014].
A given effect of a probiotic strain is strain-specific and cannot be extrapolated to other strains of the same genus or even to other strains belonging to the same species and sub-species [FAO/WHO, 2001; WGO, 2012; Hill et al. 2014; Liévin-Le Moal and Servin, 2014]. The main well-described Lactobacillus strains of human intestinal microbiota origin used to treat human gastrointestinal infectious disorders are L. rhamnosus strain [Segers and Lebeer, 2014], L. casei strain Shirota YIT9029, L. acidophilus strain LB, L. johnsonii NCC 533, L. casei DN-114 001 and L. reuteri DSM 17938. Antimicrobial activities of these strains have been demonstrated in a comprehensive set of in vitro and in vivo experiments revealing the involvement of their cell-associated components and secreted molecules [Lebeer et al. 2008, 2010; Gareau et al. 2010; Kleerebezem et al. 2010; van Baarlen et al. 2013; Liévin-Le Moal and Servin, 2014]. In addition, well-designed and well-conducted randomized controlled trials (RCTs) have demonstrated their therapeutic efficacies against infectious acute watery diarrhoea and antibiotic-associated diarrhoea [Verna and Lucak, 2010; Yan and Polk, 2012; Liévin-Le Moal and Servin, 2014; Vandenplas et al. 2015].
Several lactic acid producing and non-lactic acid producing probiotic strains have been classified as ‘live biotherapeutics’ with a therapeutic effect and these are intended for clinical use according to the US Food and Drug Administration (FDA) [Hoffman, 2008; Sutton, 2008] and the European Society for Paediatric Gastroenterology, Haepatology and Nutrition (ESPGHAN) [Szajewska et al. 2014a]. Recommendations graded from A to B for the clinical use of probiotics in necrotizing enterocolitis, childhood diarrhoea, inflammatory bowel disease, irritable bowel syndrome, antibiotic-associated diarrhoea and C. difficile-induced diarrhoea were published following a Yale University Workshop in 2005 [Floch et al. 2006]; an ‘A’ recommendation was made for strong positive, well-conducted RCTs; a ‘B’ recommendation for RCTs that did not reach ‘A’ level because of some negative studies or a limited number of studies; and a ‘C’ recommendation for RCTs that were significant but failed to receive stronger ratings because of the size of the reported patient studies, and also the factors that limited categories to ‘B’. The probiotic L. rhamnosus GG [
Historical perspectives
L. acidophilus strain LB (Lactobacillus Boucard) was isolated from the stool of a healthy human by Dr Pierre Boucard in 1907. A pharmaceutical company (Laboratoire du Lactéol, France) was founded to manufacture L. acidophilus strain LB for the development of the antidiarrheal drug called Lactéol®, first used in France and then worldwide. Originally, two pharmaceutical forms existed: a tablet form containing heat-treated (110°C, 1 hour) L. acidophilus LB cells alone (1907) and a glass ampoule form containing the complete heat-treated L. acidophilus LB culture (110°C, 1 hour) (1928). The L. acidophilus LB culture has now been molecularly identified as a culture containing L. fermentum (L. fermentum LB-f) (CNCM I-2998) [Chauviere et al. 2005; Servin et al. 2006] and L. delbrueki (L. delbrueki LB-d) (CNCM I-4831) strains with a ratio of 95:5, respectively, when cultured in both laboratory conditions and industrial batches. The adult capsule (1978) and sachet (1988) pharmaceutical forms consist of a combination of 10 billion lyophilised, heat-treated Lactobacillus LB cells (L. fermentum LB-f and L. delbrueki LB-d) and 160 mg of concentrated, neutralized spent culture medium. Three AMMs (Autorisations de mise sur le marché) (1978, 1988, 2000) for the antidiarrheal heat-treated and lyophilized L. acidophilus LB cells and concentrated neutralized spent culture medium have been given by the French National Agency for the Safety of Medicine and Health Products (Agence nationale de sécurité du médicament et des produits de santé) with the therapeutic indication for the symptomatic treatment of diarrhoea in addition to rehydration by oral rehydration solution (ORS) or intravenously in infants and children under 6 years of age, and in addition to rehydration and/or dietary measures in adults and children over 6 years.
Live L. acidophilus LB (L. fermentum LB-f and L. delbrueckii LB-d) meet several criteria defined for probiotic strains including the history, phenotypic and genotypic characterization of the strains, and the depositing of the two constitutive L. fermentum LB-f and L. delbrueki LB-d strains into a qualified cell-bank system [FAO/WHO, 2001, 2002; Hill et al. 2014]. The heat-treated and lyophilized L. acidophilus LB cells and culture medium meet the criteria defined for the therapeutic use of a biotherapeutic agent, i.e. the step-by-step description of the manufacturing process which is carried out under aseptic conditions, and the quality control process including the process input parameters and the expected output results which ensure that the product remains stable and that the biotherapeutic agent remains pharmacologically active over the indicated conservation time.
In vitro and animal studies have provided useful background information on the pharmacological antibacterial effects of the live or heat-treated L. acidophilus LB culture. Several mechanisms of action have been identified as directly directed against human gastroenterovirulent bacteria or directed against the mechanisms of pathogenesis of these bacterial pathogens that promote dramatic structural injuries at the gastrointestinal epithelial barrier accompanied by dysfunctions that caused the acute and persistent diarrhoeal diseases in children and adults.
The goal of this review is to analyse the reported experimental in vitro and in vivo data that show the antagonistic activities against gastroenterovirulent bacterial pathogens exerted by the live or heat-treated L. acidophilus LB culture, the L. fermentum strain LB-f (the major strain of the L. acidophilus LB culture) and the rehydrated powder of the pharmaceutical capsule form. Moreover, the review describes and analyses the results obtained in non-RCTs and RCTs that demonstrate the therapeutic efficacy and safety of heat-treated and lyophilized L. acidophilus LB cells and culture medium in the treatment of some gastrointestinal infectious diseases.
Antibiotic-like activity
Secreted molecules present in Lactobacillus LB culture exert a time-dependent killing activity against major enterovirulent bacteria such as Salmonella enterica serovar typhimurium (S. typhimurium), Listeria monocytogenes, Shigella flexneri, Yersinia enterocolitica, enterotoxigenic Escherichia coli, enteropathogenic E. coli and diarrhoea-associated diffusely adhering E. coli (DAEC) expressing Afa/Dr adhesins [Coconnier et al. 1997, 2000; Liévin-Le Moal et al. 2002; Coconnier-Polter et al. 2005] and gastritis-associated Helicobacter pylori [Coconnier et al. 1998] (Table 1). A bactericidal level of activity has been reached against S. flexneri, L. monocytogenes, S. typhimurium and H. pylori after 2–3 hours of direct contact. Enterovirulent E. coli is less sensitive to the killing activity of secreted molecules of Lactobacillus LB. As shown in Figure 1a, a bactericidal level of activity is exerted against the S. typhimurium strain SL1344 by untreated and heat-treated secreted molecules of Lactobacillus LB [Coconnier et al. 1997] and L. fermentum strain LB-f, and by heat-stable secreted molecules present in the supernatant of the rehydrated powder of the pharmaceutical capsule form. After treatment observation of S. typhimurium by transmission electron microscopy shows an altered cell surface without affecting the expression of flagella [Coconnier et al. 1997; Liévin-Le Moal et al. 2011]. This induced S. typhimurium SL1344 cell death was preceded by a loss of intracellular adenosine triphosphate (ATP), a release of lipopolysaccharide, and an increase in membrane permeability [Coconnier-Polter et al. 2005].
Overview of experimental in vitro and in vivo antibacterial effects of Lactobacillus acidophilus LB (L. fermentum LB-f + L. delbrueki LB-d) against specific gastroenterovirulent bacterial pathogens.
ATP, adenosine triphosphate; CFCS, cell-free culture supernatant; LPS, lipopolysaccharide; Sat, secreted auto transporter.
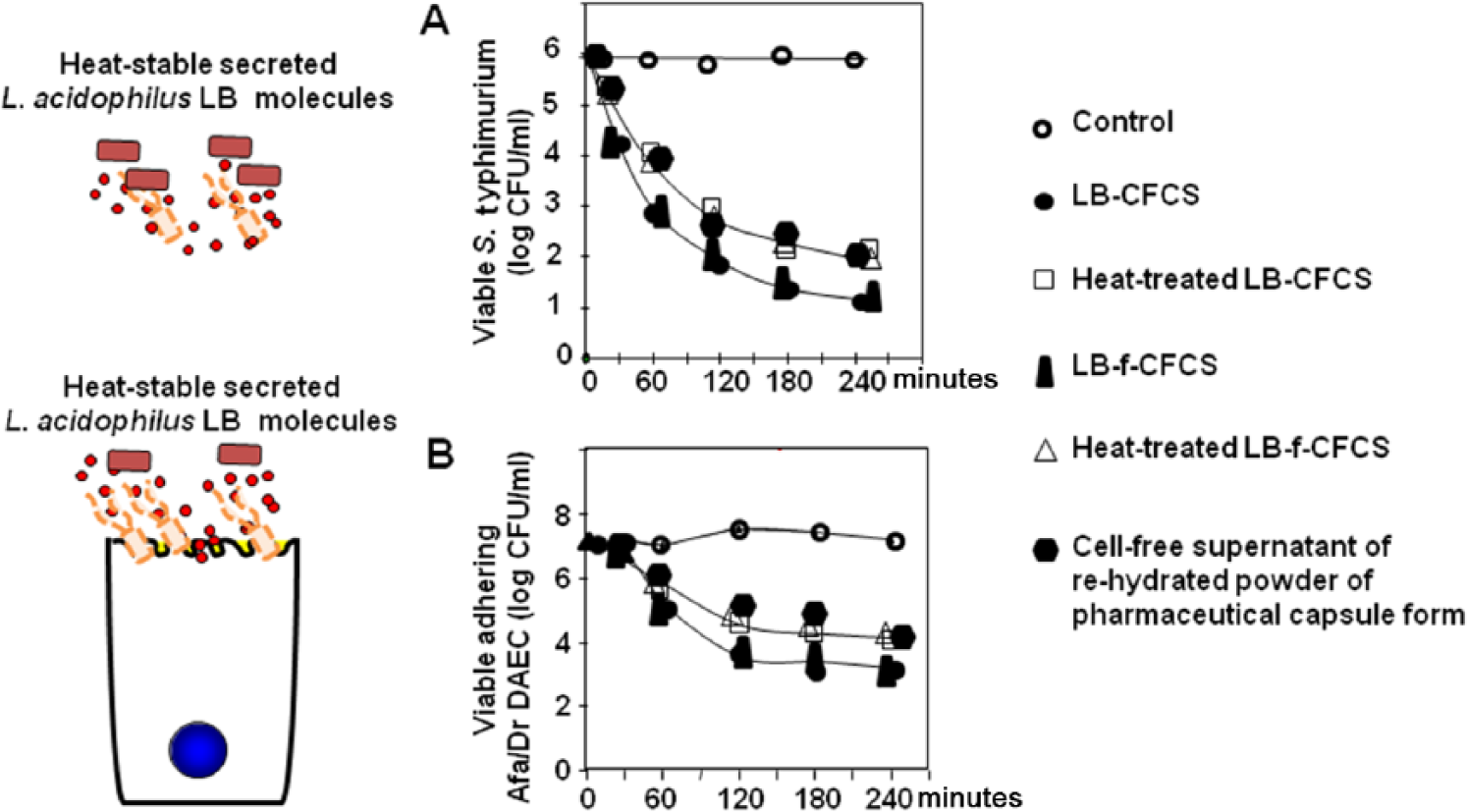
Bactericidal effect of Lactobacillus acidophilus LB (L. fermentum LB-f + L. delbrueki LB-d) on enterovirulent pathogens. (A) Time course of the bactericidal effect by direct contact against S. typhimurium strain SL1344. (B), Time course of the bactericidal effect against a diarrhoea-associated diffusely adhering E. coli strain C1845 adhering onto the brush border of cultured human enterocyte-like Caco-2/TC7 cells.
The bactericidal activity of Lactobacillus LB was sensitive for a small part to trypsin treatment and disappears after dialysis (molecular mass cutoff, 1000 Da), indicating the involvement of heat-stable and small nonproteinaceous secreted molecules [Coconnier et al. 1997]. In addition, bactericidal activity against H. pylori, the numerically dominant gastric microorganism associated with gastritis, and an increased risk of noncardia gastric adenocarcinoma, gastric lymphoma, and peptic ulcer [Cover and Blaser, 2009] has been observed exerted by secreted molecules of Lactobacillus LB, resulting in a dramatic alteration of the normal helical morphology of the pathogen and the appearance of a U-shaped form considered as a pre-coccoid death form [Coconnier et al. 1998]. This bactericidal activity resembles the morphological changes observed after the treatment of H. pylori with antibiotics used in the classical anti H. pylori triple therapy.
Antagonistic effects against activities of bacterial virulence factors
Enterotoxigenic E. coli (ETEC) are a main cause of diarrhoea in young children under 5 years of age in developing countries and also in adult travellers visiting endemic areas. By way of their heat-stable and heat-sensitive toxins, ETEC produce a cholera-like, watery diarrhoeal disease [Isidean et al. 2011]. In suckling mice infected with the enterotoxigenic E coli strain B41 which induces a toxin-triggered mouse death, administration of the rehydrated powder of the pharmaceutical form results in a significant increase in mouse survival [Fourniat et al. 1986] (Table 1).
Secreted molecules of Lactobacillus LB at a sub-bactericidal concentration promote a decrease in the attachment of diarrhoea-associated Afa/Dr DAEC strain C1845 at the brush border of enterocyte-like Caco-2 cells [Liévin-Le Moal et al. 2002], indicating an activity at the level of the interaction between the F1848 bacterial adhesin and the brush border membrane-associated receptors of this adhesin, i.e. the decay-accelerating factor and members of the human
The S. typhimurium cells swim onto the intestinal content to cross the mucus layer and the swimming motility facilitates the interaction of the pathogen with the brush border of enterocytes lining the intestinal epithelial barrier [Fabrega and Vila, 2013]. Secreted molecules of Lactobacillus LB at a sub-bactericidal concentration transiently impair the functionality of S. typhimurium SL1344 flagella without affecting the flagella expression [Liévin-Le Moal et al. 2011] (Table 1). This effect is the result of heat-stable secreted molecules with a molecular mass of less than 700 Da, which triggers a destabilization of the S. typhimurium membrane accompanied by a membrane depolarization affecting the functionality of the flagellar motor [Liévin-Le Moal et al. 2011]. Importantly, the supernatant of the rehydrated powder of the pharmaceutical capsule form exerted this blockade of S. typhimurium swimming motility [Liévin-Le Moal et al. 2011].
Salmonella attaches onto the brush border of host intestinal cells for cell entry and triggers a dramatic membrane remodelling [Fabrega and Vila, 2013]. This phenomenon follows the localized reorganization of the cell actin cytoskeleton that is promoted by the bacterial delivery onto the host cell via the type 3 secretion system (T3SS) dependent molecular syringe apparatus of a complex set of Salmonella pathogenicity island-1 (SPI-1) and island-2 effectors that hijack host cell signalling pathways. When S. typhimurium SL1344 is subjected to a sub-bactericidal concentration of secreted molecules of Lactobacillus LB prior to infection, there was an absence of F-actin dense region surrounding attached bacteria at the brush border of enterocyte-like Caco-2/TC7 cells [Coconnier et al. 2000; Liévin-Le Moal et al. 2011] (Table 1). This effect develops without affecting the presence of the T3SS needle, and modifies the expression levels of the invH gene coding for a component of the T3SS complex, and SPI-1 genes coding for SipA, SipB, SipD and SopD effectors [Liévin-Le Moal et al. 2011]. In consequence, it is tempting to hypothesize that the secreted molecules altered the functionality of the Salmonella T3SS syringe.
The secreted auto transporter toxin (Sat) of enterovirulent Afa/Dr DAEC promoted functional lesions at the brush border by increasing the transcellular passage of fluid across the intestinal barrier and inducing an accumulation of fluid within the intestinal lumen [Servin, 2014]. Both the untreated and heat-treated secreted molecule(s) of Lactobacillus LB at sub-bactericidal concentrations, and the supernatant of the rehydrated powder of the pharmaceutical capsule form concentration-dependent, antagonized Sat toxin activation of transcellular passage of fluid [Liévin-Le Moal et al. 2007] (Table 1).
Antagonist effects at mucosa
Secreted molecules of Lactobacillus LB exert a bactericidal activity against the diarrhoea-associated Afa/Dr DAEC C1845 bacteria attached at the brush border of enterocyte-like Caco-2/TC7 cells [Liévin-Le Moal et al. 2002]. As shown in Figure 1b, this bactericidal activity is also exerted by the secreted molecules of L. fermentum strain LB-f and the supernatant of the rehydrated powder of the pharmaceutical capsule form. Similarly, when the cultured human HT29-MTX cells secreting mucins of gastric immunoreactivity were infected with H. pylori, there was a dose-dependent decrease in the viability of the adhering pathogen after treatment with the secreted molecules of Lactobacillus LB [Coconnier et al. 1998] (Table 1). Moreover, the remaining adhering H. pylori showed a decreased urease activity and displayed an altered morphology.
Both live and heat-treated Lactobacillus LB cells displayed a high adhesiveness capacity to adhere onto enterocyte-like Caco-2 cells and mucus-secreting HT29-MTX cells [Chauviere et al. 1992b; Coconnier et al. 1993a]. As a result of this high capacity of adhesion, Lactobacillus LB cells and L. fermentum LB-f cells, and the cells present in the rehydrated powder of the pharmaceutical capsule form, exert a competitive inhibition activity to prevent the attachment at the brush border of different classes of enterovirulent E. coli and S. typhimurium (Table 1). Adhesion of E. coli strain B41 onto cultured epithelial cervix HeLa cells was inhibited competitively by heat-treated Lactobacillus LB cells and inhibitory activity was lost after lysis of the Lactobacillus cells, suggesting a steric hindrance of E coli adhesion sites at the epithelial cell membrane rather than competition for a common binding site [Fourniat et al. 1992]. Examined in competition-binding conditions in which bacterial pathogens and Lactobacillus cultures co-incubate with the cultured enterocyte-like Caco-2 cells, the attachment at the brush border of human ETEC H10407 and Pb176 strains expressing colonization factors CFA/I and CFA-II, respectively, was concentration-dependently inhibited by both live and heat-treated Lactobacillus LB cultures [Chauviere et al. 1992a] (Table 1).
As shown in Figure 2a, untreated and heat-treated Lactobacillus LB and L. fermentum LB-f cells, and cells isolated from the rehydrated powder of the pharmaceutical capsule form, exert a similar competitive inhibition activity against the attachment of ETEC H10407 at the brush border of enterocyte-like Caco-2/TC7 cells. Identically, when enterocyte-like Caco-2 cells were infected with enteropathogenic E. coli (EPEC) strain JPN15, L. monocytogenes strain EGD, Y. pseudotuberculosis [Coconnier et al. 1993a, 1993b] or a diarrhoea-associated Afa/Dr DAEC strain C1845 [Liévin-Le Moal et al. 2002], the attachment of pathogens at the brush border was concentration-dependently inhibited by untreated and heat-treated Lactobacillus LB cultures (Table 1). Similarly, live and heat-treated Lactobacillus LB cultures [Coconnier et al. 1993a, 1993b], and the rehydrated powder of the pharmaceutical capsule form [Liévin-Le Moal et al. 2011], promote the inhibition of cell association of S. typhimurium SL1344 in enterocyte-like Caco-2/TC7 cells (Table 1).
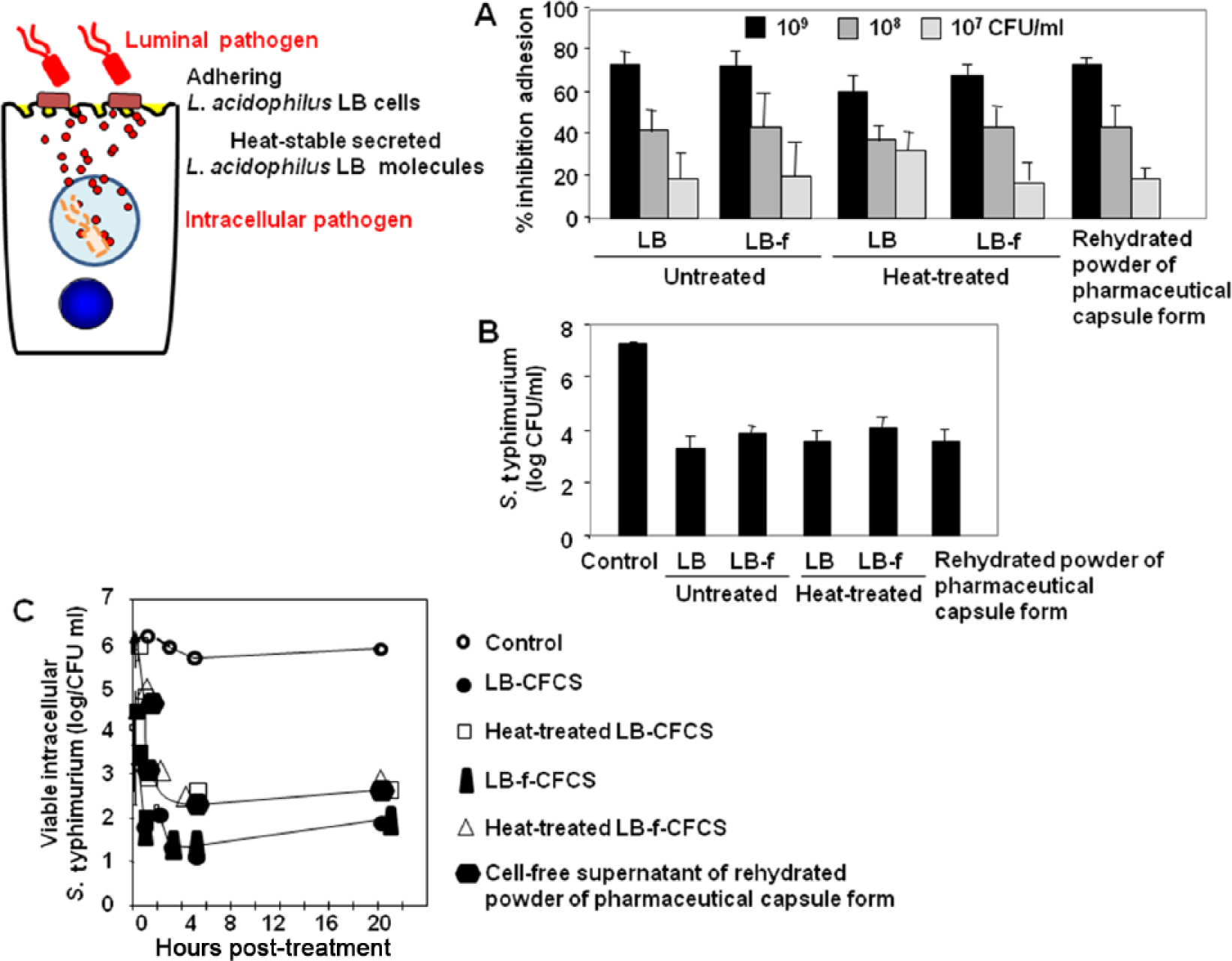
Antagonistic effects of Lactobacillus acidophilus LB (L. fermentum LB-f + L. delbrueki LB-d) against adhesion and/or invasion of enterovirulent pathogens in cultured human enterocyte-like cells. (A) Concentration-dependent competitive inhibition of adhesion of diarrhoea-associated enterotoxigenic Escherichia coli strain H10407 expressing CFA-I adhesive fimbriae onto the brush border of Caco-2/TC7 cells. (B) Inhibition of the internalization of Salmonella typhimurium SL1344 into Caco-2/TC7 cells. (C) Time course of the bactericidal effect against S. typhimurium SL1344 residing in intracellular vacuoles after cell entry of the pathogen within Caco-2/TC7 cells.
S. typhimurium after attachment onto the brush border of host enterocytes penetrates the cells [Fabrega and Vila, 2013]. Untreated and heat-treated Lactobacillus LB cultures, and untreated and heat-treated cell-free culture supernatants of Lactobacillus LB, inhibit the cell-entry of S. typhimurium in enterocyte-like Caco-2/TC7 cells [Coconnier et al. 1997, 2000; Coconnier-Polter et al. 2005; Liévin-Le Moal et al. 2011] (Table 1). As shown in Figure 2b, S. typhimurium SL1344 shows a similar decreased cell entry in Caco-2/TC7 cells when subjected to untreated and heat-treated secreted molecules of Lactobacillus LB or L. fermentum strain LB-f at subinhibitory bactericidal concentrations, or the supernatant of the rehydrated powder of the pharmaceutical capsule form during cell infection.
After cell entry S. typhimurium resides in the intracellular vacuoles of enterocytes and adapts to this situation by exploiting multiple endogenous host nutrients [Fabrega and Vila, 2013]. When S. typhimurium SL1344 resides intracellularly in vacuoles in pre-infected enterocyte-like Caco-2/TC7 cells, the secreted molecules of Lactobacillus LB exert a bactericidal activity against the intracellular bacteria [Coconnier et al. 2000] (Table 1). As shown in Figure 2c, untreated and heat-treated secreted molecules of Lactobacillus LB and L. fermentum strain LB-f, and the supernatant of the rehydrated powder of the pharmaceutical capsule form, exert an identical bactericidal activity against the intracellular S. typhimurium cells. In contrast, none of the antibiotics tested, including cephalothin, tetracycline, amoxicillin, clavulanic acid, gentamycin and pefloxacin, displayed any bactericidal activity against S. typhimurium located in the intracellular vacuoles [Coconnier et al. 2000]. Observation by transmission electron microscopy of the remaining present S. typhimurium cells located within the intracellular vacuoles shows that the cells displayed a death morphology [Coconnier et al. 2000] (Table 1). This effect at the intracellular level is of interest since it is well-established that several intracellular enterovirulent bacteria constitute a population of persistent and dormant pathogenic bacteria that play an important role in recurrent infections [Lewis, 2007].
The brush border of enterocytes is constituted by a regular array of microvilli, the membrane of which is endowed with a number of hydrolytic enzymes that are involved in the digestion of dietary carbohydrates and peptides, and transporters such as sucrase–isomaltase (SI), lactase, amino-peptidase N, dipeptidylpeptidase IV (DPP IV), alkaline phosphatase (APL) and hexose transporters (SGLT1, GLUT1, GLUT2, GLUT3 and GLUT5) [Turner, 2009]. Following diarrhoea-associated Afa/Dr DAEC cell infection, the disappearance of the brush border is accompanied by a dramatic loss of intestinal functions [Servin, 2014]. In Afa/Dr DAEC C1845-infected enterocyte-like Caco-2/TC7 cells, the treatment with secreted molecule(s) of Lactobacillus LB at sub-bactericidal concentrations results in the maintenance of a normal F-actin brush border cytoskeleton and by normal expression of microvillus-associated functional proteins SI, DPP IV, AP and GLUT5 [Liévin-Le Moal et al. 2002] (Table 1).
In enterocyte-like Caco-2/TC7 cells, untreated and heat-treated secreted molecules of Lactobacillus LB, and the supernatant of the rehydrated powder of the pharmaceutical capsule form, exerted an antagonistic activity against the Sat toxin-secretory activity since there was a concentration-dependent inhibition of the toxin-induced increase of fluid domes formed by an increased apical to basal transcellular passage of water [Liévin-Le Moal et al. 2007] (Table 1).
Closure of the intestinal barrier is maintained by intercellular junctional complexes composed of tight junctions (TJs), adherent junctions and desmosomes. In particular, the TJs are highly developed structures that function as a ‘gate’ between the mucosal and serous environments controlling the paracellular vectorial transports occurring across the intestinal barrier [Turner, 2009]. The rehydrated powder of the pharmaceutical capsule form protects the TJs of cultured human intestinal HT-29 cells by counteracting the aspirin-induced delocalization and loss of expression of structural TJs-associated structural ZO-1 protein [Montalto et al. 2004].
Anti-infectious effects in infected animal models
Antibacterial activity of L. acidophilus LB against diarrhoea-associated bacterial pathogens has also been investigated in different infectious mice models (Table 1). In newborn mice infected with the bovine enterotoxigenic ETEC strain B41, all the infected mice were dead 72 hours after infection; however, there was a high significant increase of the survival rate when the infected animals received the rehydrated powder of the pharmaceutical form orally every day [Fourniat et al. 1986]. In comparison with erythromycin treatment, oral administration of the rehydrated powder of the pharmaceutical capsule form in germ-free mice infected with the invasive Campylobacter jejuni strain 79K results in an accelerated decrease of pathogen colonization of ileum and colon mucosa without modifying the level of the pathogen within the intestinal content and a high significant decrease of translocation of the pathogen into mesenteric lymph nodes [Moyen et al. 1986]. Oral administration of secreted molecules of Lactobacillus LB in S. typhimurium strain C5 infected conventional C3H/He/oujco mice promotes a high significant decrease of the level of live pathogen in the faeces compared with untreated mice [Coconnier et al. 1997]. Oral treatment of H. felis infected conventional BALB/c mice with secreted molecules of Lactobacillus LB decreases the level of gastric mucosa-associated H. felis, reduces the urease activity of the remaining mucosa-attached H. felis, and lowers the level of H. felis induced gastric inflammation characterized by a diminished number of infiltrated inflammatory lymphocytes, plasma cells, and polymorphonuclear cells in the antrum and corpus mucosa [Coconnier et al. 1998] (Table 1).
As analysed above, the untreated or heat-treated Lactobacillus LB (L. fermentum LB-f and L. delbrueckii LB-d) and L. fermentum strain LB-f cells, and their heat-stable secreted molecules are experimentally active in vitro and in vivo against a large set of gastroenterovirulent bacteria triggering deleterious structural and functional lesions in the stomach or intestinal barrier leading to acute watery diarrhoea or gastritis. The identified mechanisms underlying the anti-infectious effects of heat-treated, lyophilized Lactobacillus LB cells and concentrated, neutralized spent culture medium composing the pharmaceutical forms appear to be multifaceted (summarized in Figure 3). The heat-treated whole cells inhibited the cell attachment and internalization of gastrointestinal bacterial pathogens with cultured human intestinal epithelial cells. Heat-stable secreted molecules have a direct bactericidal effect after direct contact with gastrointestinal bacterial pathogens at the luminal level and against the brush border associated or internalized gastrointestinal bacterial pathogens. In addition, heat-stable secreted molecules inhibited the functionality of bacterial virulence factors.

Summary of mechanisms of action of Lactobacillus acidophilus LB (L. fermentum LB-f + L. delbrueki LB-d) against human diarrhoea-associated bacterial pathogens.
These anti-bacterial effects indicate that, in the infectious situation, there is an absence of bacterial pathogens-induced, signalling-dependent structural and functional lesions at the intestinal mucosa and junctional levels, and gastritis-associated inflammatory lesions at the gastric mucosa. Moreover, the observation that heat-stable secreted molecules are active at a sub-bactericidal concentration suggests the presence of molecules exerting antagonistic or regulatory activities against the pathogen-activated cell signalling pathways at the intestinal epithelial barrier. In addition, effects observed in non-infectious situations, i.e. in the presence of aspirin-induced TJs lesions [Montalto et al. 2004] and Sat toxin-induced activation of transcellular passage of fluids [Liévin-Le Moal et al. 2007], and in patients with diarrhoea-predominant irritable bowel syndrome [Halpern et al. 1996; Tarrerias et al. 2011; Andresen et al. 2012] suggest the production by L. acidophilus LB of compounds having regulatory activities onto intestinal cellular functions.
Safety
In a recent analysis conducted to analyse the safety of organisms from six different genera used as probiotic agents including Lactobacillus alone or in combination, and including clinical trials conducted with the heat-treated and lyophilized L. acidophilus LB cells and concentrated, neutralized spent culture medium, Hempel and colleagues concluded that there was an absence of reported adverse events in intervention studies conducted to reduce the risk, prevent or treat gastrointestinal infectious diseases [Hempel et al. 2011]. The safety of heat-treated and lyophilized L. acidophilus LB cells and culture medium has been indicated in two RCTs [Liévin-Le Moal et al. 2007; Salazar-Lindo et al. 2007] and there was an absence of reported adverse effects in others RCTs [Boulloche et al. 1994; Simakachorn et al. 2000].
Clinical efficacy against enterovirulent bacteria and rotavirus induced acute watery diarrhoea
The World Health Organization (WHO) has defined diarrhoea as the passage of 3 or more loose or watery stools within a 24-hour period. Diarrhoea is described as being acute if the illness started less than 14 days previously, and persistent if the episode lasts 14 days or more. Worldwide, rotavirus is a major common cause of severe diarrhoea and diarrhoea-related mortality in children [Hagbom et al. 2012]. The major bacterial pathogens involved in diarrhoea are ETEC, Salmonella, Shigella, Yersinia, Campylobacter and Vibrio cholera [Croxen et al. 2013]. Moreover, ETEC-induced acute diarrhoea is common among travellers and tourists. To prevent dehydration and nutritional damage in children and infants with infectious acute watery diarrhoea, the therapeutic regimen recommended by the WHO, the European Society for Paediatric Infectious Diseases, and ESPGHAN is to provide an ORS and to continue feeding [Guarino et al. 2008; Piescik-Lech et al. 2013]. Although oral rehydration effectively mitigates dehydration, it has no effect on the infection causes and the duration, severity or frequency of diarrhoeal episodes. A variety of luminally acting agents are used to treat diarrhoea by altering intestinal and/or colonic motility, or fluid secretion, but some of which have adverse effects [Menees et al. 2012]. Innovative therapeutics in the pipeline target intestinal ion channels and transporters, cell surface receptors, and cell signaling regulatory proteins [Thiagarajah et al. 2015]. Recommendations have been given for the use of a limited number of probiotic strains as an adjunct to rehydration therapy for the treatment of acute gastroenteritis in previously healthy infants and children [Szajewska et al. 2014a].
Non-RCTs and RCTs have shown the therapeutic efficacy of heat-treated and lyophilized L. acidophilus LB cells and culture medium (Drug) in conjunction with ORS for the treatment of rotavirus-induced, well-established acute watery diarrhoea in children (Table 2). In infants and young children with well-established acute watery diarrhoea of undetermined infectious origin, Boulloche and colleagues [Boulloche et al. 1994] reported that treatment with the heat-treated and lyophilized L. acidophilus LB cells and culture medium had resulted in a shortening of the duration of diarrhoea and an acceleration of the reappearance of the first normal consistent stool from 64.7 hours in placebo + ORS group versus 49.7 hours in Drug + ORS group [mean difference (MD) −15.0 hours, 95% confidence intervals (95% CI) −28.99 to −1.01]. Simakachorn and colleagues [Simakachorn et al. 2000] have shown in children aged 3–24 months with rotavirus-induced, well-established acute watery diarrhoea that Drug + ORS treatment reduced the duration of diarrhoea from 57.0 hours to 43.4 hours (MD −13.6 hours, 95% CI −28.10 to 0.90), accompanied by an accelerated reappearance of formed stools compared with placebo + ORS treatment. Interestingly, in children with identified rotavirus-associated watery diarrhoea, only 16% of the children had watery stools after the first day of Drug + ORS treatment whereas 56% had watery stools in the group treated with placebo + ORS. Additionally, a non-RCT conducted in infants and young children mainly with rotavirus-associated diarrhoea has shown that the treatment with heat-treated and lyophilized L. acidophilus LB cells and culture medium leads to a high significant reduction of the number of stools per day [Li Xie, 1995].
Overviews of clinical therapeutic anti-infectious effects of lyophilized, heat-treated Lactobacillus LB cells and concentrated, neutralized spent culture medium.

Drug: sachet or capsule pharmaceutical forms (Lactéol
ORS, oral rehydration solution.
The therapeutic efficacy of probiotic Lactobacillus strains against established enterovirulent bacteria-induced acute watery diarrhoea in children has rarely evidenced in RCTs [Liévin-Le Moal and Servin, 2014]. Consistent with the in vitro and in vivo antibacterial activities of L. acidophilus LB against the major diarrhoea-associated bacterial pathogens described above, two RCTs have shown that heat-treated and lyophilized L. acidophilus LB cells and culture medium treatment was therapeutically effective in children with established enterovirulent bacteria-induced diarrhoea. Salazar-Lindo and colleagues [Salazar-Lindo et al. 2007] have shown in an RCT an age-dependent shortening of the duration of diarrhoea in children aged 2–4 after treatment with the heat-treated and lyophilized L. acidophilus LB cells and culture medium. While there is an absence of therapeutic effect with Drug + ORS versus placebo + ORS in overall enrolled children, a subclassification of children in two groups as a function of the duration of diarrhoea at the inclusion showed that Drug + ORS versus placebo + ORS is ineffective in the reduction of the duration of diarrhoea in children with mild acute watery diarrhoea (<24 hours) and is well effective in children with well-established acute watery diarrhoea (>24 hours), with a highly significant decrease in the duration of diarrhoea (8.2 hours versus 30.4 hours) (Table 2). Moreover, a comparison of children by age showed a better reduction of the duration of diarrhoea in children between the ages of 2 and 4 years (29.0 hours in placebo + ORS group versus 8.55 hours in Drug + ORS group) versus children aged 1 month to 2 years (12.3 hours in placebo + ORS group versus 10.3 hours in Drug + ORS group). Liévin-Le Moal and colleagues [Liévin-Le Moal et al. 2011] reported that in children between the ages of 1 and 34 months with infectious well-established acute watery diarrhoea of non-rotavirus origin, treatment with heat-treated and lyophilized L. acidophilus LB cells and culture medium in association with ORS reduced the duration of diarrhoea (63.4 hours in placebo + ORS group versus 39.5 hours in Drug + ORS group (MD −23.9 hours, 95% CI −29.6 to −18.2). Moreover, the diarrhoea was resolved in a significantly higher proportion of children in the Drug + ORS group (90%: 36 of 42 children) than in the placebo + ORS group (53%: 20 of 38 children), and solid stools were formed in children of the Drug + ORS group, whereas those of the children of the placebo + ORS group tended to remain fluid or semi-fluid. Xiao and colleagues [Xiao et al. 2003] showed in a multicentre RCT conducted in adults of ages 16 years and over with bacteria- and parasitic-associated chronic diarrhoea that heat-treated and lyophilized L. acidophilus LB cells and culture medium promotes a rapid recovery of consistent stools in 81% of treated patients.
Szajewska and colleagues [Szajewska et al. 2014a, 2014b] conducted a meta-analysis pooling the results of the 224 children in three RCTs conducted with heat-treated and lyophilized L. acidophilus LB cells and culture medium + ORS in comparison with placebo + ORS [Boulloche et al. 1994; Simakachorn et al. 2000; Liévin-Le Moal et al. 2007]. Results showed a significant reduction in the duration of diarrhoea for Drug + ORS compared with placebo + ORS (MD −21.6 hours, 95% CI −26.5 to −16.6). Moreover, the pooled results of 2 RCTs [Simakachorn et al. 2000; Liévin-Le Moal et al. 2007] involving 153 children showed that the Drug + ORS treatment increased the chance of cure to cure on day 4 compared with placebo + ORS [risk ratio (RR) 95% CI: 1.44].
The therapeutic efficacy of probiotic Lactobacillus strains in the treatment of the antibiotic-associated diarrhoea has been clinically proven [Hickson, 2011; Johnston et al. 2011, 2012; Hempel et al. 2012]. In a RCT conducted in 200 adult patients who received an antibiotic treatment, the heat-treated and lyophilized L. acidophilus LB cells and culture medium lead to a reduction of the duration of diarrhoea compared with patients who were receiving the antibiotics treatment alone, but did not reduce the risk of developing bloating, flatus and abdominal pain [Yap Soo Kor et al. 2010] (Table 2).
Enterovirulent bacteria-induced acute diarrhoea is common amongst travellers and tourists. Briand and colleagues [Briand et al. 2006] evaluated the efficacy of heat-treated and lyophilized L. acidophilus LB cells and culture medium in the prevention of travellers’ diarrhoea in a RCT conducted in adults travellers visiting West, East, Central and North Africa, Oceania, South America, Asia, Central America/the Caribbean and Middle East between 2001 and 2004. They reported that treatment before departure twice daily with heat-treated and lyophilized L. acidophilus LB cells and culture medium had no effect on the incidence of diarrhoea in the treated group compared with the placebo group during the interval of 27 days of travel (Table 2). This observation is consistent with previous RCTs showing the lack of therapeutic efficacy for the prevention of intestinal infectious episodes in travellers by the administration of probiotic Lactobacillus strains (Ritchie and Romanuk, 2012; Liévin-Le Moal and Servin, 2014].
Comparison with different marketed probiotic Lactobacillus strains
The level of therapeutic efficacy of heat-treated and lyophilized L. acidophilus LB cells and culture medium against established infectious acute watery diarrhoea (MD −21.6 hours, 95% CI −26.5 to −16.6) [Szajewska et al. 2014a] is close to the level found by Allen and colleagues [Allen et al. 2010] in their Cochrane analysis of 63 RCTs and quasi-randomized controlled trials monitored with different Lactobacillus probiotic strains for the treatment of infectious acute watery diarrhoea (various aetiology origins; MD −24.76 hours, 95% CI −15.9 to −33.6).
L. rhamnosus GG, L. casei Shirota YIT9029, L. johnsonii NCC 533, L. casei DN-114 001 and L. reuteri DSM 17938 are the main experimentally and clinically well-described Lactobacillus strains of human intestinal microbiota origin [Liévin-Le Moal and Servin, 2014].
L. rhamnosus GG displays a similar therapeutic efficacy against infectious acute watery diarrhoea (11 RCTs; aetiology: mainly rotavirus-induced and unknown causes; MD −25.2 hours, 95% CI −40.8 to −9.6) [Szajewska et al. 2007b, 2013b, 2014a; Guarino et al. 2008; Liévin-Le Moal and Servin, 2014] compared with heat-treated and lyophilized L. acidophilus LB cells and culture medium (3 RCTs; aetiology: 80–100% rotavirus- and enterovirulent bacteria-induced diarrhoea; MD −21.6 hours, 95% CI −26.5 to −16.6). In contrast to heat-treated and lyophilized L. acidophilus LB cells and culture medium, L. rhamnosus GG displays no therapeutic efficacy against enterovirulent bacteria-induced acute watery diarrhoea [Liévin-Le Moal and Servin, 2014]. The probiotic L. reuteri strain 17938 shows a similar therapeutic efficacy against infectious acute watery diarrhoea (5 RCTs; aetiology: 60% rotavirus-induced diarrhoea and unknown causes; MD −32 hours, 95% CI −41.0 to −24.0) [Guarino et al. 2008; Szajewska et al. 2013a, 2014a; Liévin-Le Moal and Servin, 2014] compared with heat-treated and lyophilized L. acidophilus LB cells and culture medium (MD −21.6 hours, 95% CI −26.5 to −16.6).
There is an absence of RCTs conducted with probiotic L. casei Shirota YIT9029 and L. johnsonii NCC 533 strains against infectious acute watery diarrhoea [Liévin-Le Moal and Servin, 2014]. Other probiotic Lactobacillus strains isolated from the human intestinal microbiota are present in products [WGO, 2012] such as L. acidophilus strain NCFM (also designated RL8K/NCK45/NCK56/N2) (ATCC 700396) [Sanders and Klaenhammer, 2001], L. plantarum 299v (DSM 6595) and L. fermentum ME-3 (DSM 14241) [Mikelsaar and Zilmer, 2009]. These probiotic strains have been tested in vitro against gastrointestinal bacterial pathogens or in animal infection models, but there was an absence of RCTs conducted in human infection gastrointestinal situations. For the L. rhamnosus R0011 (CNCM I-1720) and L. helveticus R0052 (CNCM I-1722) strains isolated from dairy [Foster et al. 2011], there was only one RCT (unknown aetiology; MD −23.8 hours, 95% CI −60.0 to −9.6) [Szajewska et al. 2014a].
It is noteworthy that the probiotic, non-lactic acid producing yeast strain Saccharomyces boulardii displays a therapeutic efficacy against acute watery diarrhoea of diverse infectious origins (11 RCTs; MD −23.8 hours, 95% CI −33.6 to −14.4) [Szajewska and Skorka, 2009; Szajewska et al. 2014a]. Moreover, S. boulardii exerts a strong therapeutic efficacy against antibiotic or Clostidium difficile associated diarrhoea [Szajewska et al. 2007a; Guarino et al. 2008; McFarland, 2010; Johnston et al. 2011, 2012].
While stating that the heat-treated and lyophilized L. acidophilus LB cells and culture medium cannot be considered as a probiotic, the ESPGHAN Working Group for Probiotics and Prebiotics included the results of RCTs conducted with heat-treated and lyophilized L. acidophilus LB cells and culture medium in a recent comparison of RCTs conducted with probiotic products for the treatment of acute gastroenteritis [Szajewska et al. 2014a]. Two groups of products have been proposed, the first containing products with a positive recommendation and the second including products with insufficient evidence to make a recommendation. The heat-treated and lyophilized L. acidophilus LB cells and culture medium has been classified in the first group together with the L. rhamnosus GG and L. reuteri DSM 17938 strains, and the yeast strain S. boulardii. However, an additional mention indicates limited or weak evidences for L. reuteri DSM 17938 and heat-treated and lyophilized L. acidophilus LB cells and culture medium on the basis that the quality of the clinical efficacy was downgraded by the methodological limitations of the trials and by the low number of published RCTs.
The most common forms containing probiotics Lactobacillus strains are dairy products, although tablets, capsules and sachets containing freeze-dried probiotic bacteria are also available [WGO, 2012]. Stability dependence on various factors is critical to guarantee the efficacy of the pharmacological effects ascribed to a probiotic strain present in the final product formulation [Sanders et al. 2014]. As decay of the viable cells count is a phenomenon typical of every formulation containing viable probiotic bacteria [Del Piano et al. 2006], the strict observance of the storage conditions is needed to guarantee that the product remains stable and pharmacologically effective over the indicated conservation time. For lyophilized probiotic bacteria, the conduct of the recommended resuscitation process is necessary considering that this step is critical for to obtain viable, active probiotic bacteria [Muller et al. 2010]. The pharmaceutical forms containing heat-treated and lyophilized L. acidophilus LB cells and concentrated, neutralized spent culture medium have the advantage of providing a controlled and stable quantity of heat-treated whole-cells and secreted, heat-stable anti-bacterial molecules displaying the experimentally observed pharmacological antibacterial activities exerted by the live L. acidophilus LB.
Conclusion
Experimental studies and non-RCTs and RCTs demonstrated the role of L. acidophilus LB cells and concentrated, neutralized spent culture medium as a good biotherapeutic agent to treat several gastrointestinal infectious diseases. These pharmacological anti-infectious properties fit well with the term ‘pharmabiotics’ proposed by Shanahan and colleagues [Shanahan et al. 2009]. However, some of the RCTs performed several years ago not meet the new criteria/requirements for the demonstration of the therapeutic efficacy of drugs in clinical infectious conditions. Considering the above mentioned opinion by the ESPGHAN Working Group for Probiotics and Prebiotics [Szajewska et al. 2014a] underlining the necessity to improve the evidence of the therapeutic efficacy of heat-treated L. acidophilus LB cells and concentrated, neutralized spent culture medium to treat gastrointestinal infectious disorders, additional RCTs are needed particularly in outpatients and inpatients for the treatment of established acute watery diarrhoea.
Footnotes
Acknowledgements
I wish to acknowledge all researchers who have helped to unravel aspects of antimicrobial Lactobacillus LB effects. I am grateful to M.H. Coconnier-Polter for our fruitful collaboration in identifying the mechanisms of the antimicrobial properties of Lactobacillus LB. I would like to express my earnest appreciation to A.L. Servin who has outstandingly headed the research on the antimicrobial properties of lactic acid producing probiotic strains at the Inserm and University Paris-Sud Research Units at the Faculty of Pharmacy Châtenay-Malabry. Finally I would like to thank R. Boisseau for his constant support for academic and therapeutic research.
Funding
This academic review received no specific grant from any research agency, commercial, or non-profit associations.
Conflict of interest statement
The author holds US and European patents for Lactobacillus LB and LB-f strains without holding shares or equity. The author declares no conflicts of interest in preparing this article.