
Abstract
Background:
We have recently shown that the acute infusion of angiotensin-(1–7) [Ang-(1-7)] or chronic increase in plasma Ang-(1-7) produces important changes in regional blood flow in rats.
Methods:
To further assess whether these changes are related to Mas, in this study hemodynamic measurements were performed in Ang-(1-7) receptor Mas knockout C57BL/6 (Mas-KO) mice and age-matched wild type (WT) control mice, using fluorescent microspheres.
Results:
Mean arterial pressure in urethane-anesthetized Mas-KO mice (12–16 weeks old) did not differ from that in WT mice (79 ± 2 and 80 ± 2 mmHg respectively). However, pronounced differences were observed in other hemodynamic measurements. Mas-KO mice exhibited a significant decrease in stroke volume (0.03 ± 0.01 versus 0.05 ± 0.01 ml/beat in WT) and decreased cardiac index (0.81 ± 0.08 versus 1.24 ± 0.24 ml/min/g in WT). Strikingly, Mas-KO mice exhibited a marked increase in vascular resistance and a decrease in blood flow in the kidney, lung, adrenal gland, mesentery, spleen and brown fat tissue. The decrease in blood flow ranged from 34% (spleen) to 55% (brown fat tissue).
Conclusion:
These results suggest that the Ang-(1-7)/Mas axis plays an important role in regional and systemic hemodynamic adjustments in mice.
Keywords
Introduction
The renin–angiotensin system (RAS) plays an important role in the pathogenesis of cardiovascular disease [Probstfield and O’Brien, 2010; Singh et al. 2010; Unger et al. 2011; Werner et al. 2008]. Most physiological and physiopathological effects of angiotensin (Ang) II are mediated via Ang II type 1 (AT1) receptors. The recent discovery of the Ang-converting enzyme (ACE) homologue, ACE 2 [Donoghue et al. 2000; Tipnis et al. 2000] as a major Ang-(1-7)-forming enzyme and the characterization of the G-protein-coupled receptor Mas as an Ang-(1-7) receptor [Santos et al. 2003] lends strong support to the concept that Ang-(1-7) is a biologically active component of the renin–angiotensin system [Buczko and Kucharewicz, 2000; Ferrario,1998, 2010; Loot et al. 2002; Santos et al. 2000]. Moreover, growing evidence indicates that Ang-(1-7) appears to counterbalance most of the effects of Ang II [Ferrario et al. 2005; Iusuf et al. 2008; Reudelhuber, 2005; Sampaio et al. 2007a; Santos et al. 2005; Santos and Ferreira, 2007].
Blood vessels are an important site for the opposing actions of Ang-(1-7) and Ang II. Vascular responses to Ang-(1-7) significantly differ from those induced by Ang II. Contrasting with Ang II, Ang-(1-7) in blood vessels acts mainly as a vasodilator and antiproliferative hormone [Freeman et al. 1996; Sampaio et al. 2003; Santos et al. 2000; Santos and Ferreira, 2007; Varagic et al. 2008]. Ang-(1-7) has been shown to produce relaxation of aortic rings in Sprague-Dawley (SD) [Silva et al. 2007] and mRen-2 transgenic rats [Lemos et al. 2002], canine [Brosnihan et al. 1996] and porcine coronary arteries [Porsti et al.1994], canine middle cerebral artery [Feterik et al. 2000], piglet pial arterioles [Meng and Busija, 1993], feline systemic vasculature [Osei et al. 1993], rabbit renal afferent arterioles [Ren et al. 2002] and mesenteric micro vessels of normotensive and hypertensive rats [Fernandes et al. 2001; Oliveira et al. 1999].
Endothelial cells possess Ang-(1-7) receptors [Sampaio et al. 2007b; Tallant et al. 1997]. Ang-(1-7) stimulates the production of nitric oxide (NO) in these cells and regulates endothelial cell function through functionally active Mas [Peiró et al. 2007; Sampaio et al. 2007b; Santos et al. 2005]. It has been reported that Mas deficiency leads to impaired endothelial function [Rabelo et al. 2008; Xu et al. 2008]. However, the relative contribution of Mas to blood flow in different organs is not known.
The aim of the present study was to test whether Mas deficiency is associated with regional changes in vascular resistance. We also aimed to evaluate whether the influence of Mas in the circulation is age dependent. To achieve this, fluorescent microspheres were used in mice with the genetic deletion of Mas. The data demonstrate that Mas-deficient mice exhibited marked changes in regional and systemic hemodynamics.
Materials and methods
Ethical approval
All experimental protocols were performed in compliance with the guidelines for the humane use of laboratory animals established at this institution (Ethics Committee of Animal Experiments/Federal University of Minas Gerais (CETEA/UFMG)).
Mas-knockout mice
Wild type (WT) and Mas-knockout (Mas-KO) C57BL/6 mice were obtained from the transgenic animal facilities of the Laboratory of Hypertension, Federal University of Minas Gerais (Brazil).
Surgical preparation
The mice were anesthetized with urethane (1.2 g/kg intraperitoneal injection; Sigma, Taufkirchen, Germany). Body temperature was continually monitored with a rectal probe and maintained at 36–37°C by a heating pad. The left femoral artery was cannulated with polyethylene tubing (PE-10) for the recording of blood pressure and heart rate (Biopac System Inc., Califórnia, USA.). For the administration of fluorescent microspheres, the right carotid artery was exposed and a PE-10 cannula was guided through the common carotid artery into the left ventricle. The positioning of the cannula in the left ventricle was confirmed by the blood pressure recording. The right femoral artery was cannulated and connected to a pump (Minipuls 3; Gilson, Villiers le Bel, France) for blood withdrawal.
Determination of systemic and regional hemodynamics
Systemic hemodynamics and regional blood flow were determined using 15 µm fluorescent polystyrene microspheres (FluoSpheres Blood Flow Determination; Molecular Probes, Eugene, OR, USA), as described elsewhere [Barbee et al. 1992; Richer et al. 2000; Sarin et al. 1990]. To prevent aggregation, the microsphere suspension was agitated for 2 min, followed by sonication for an additional 60 s. After the mixing was completed, 60,000 fluorescent microspheres were infused into the left ventricle over a 5 s period. To calculate blood flow, arterial blood was withdrawn at a rate of 0.25 ml/min through the right femoral artery. Blood for the reference sample was withdrawn for 55 s starting 5 s before the microsphere injection. At the end of the experiments, the animals were euthanized with an overdose of anesthesia and the tissues (kidneys, brain, mesentery, adrenal glands, spleen, abdominal skin, gastrocnemius muscle, lungs, testes, left ventricle, brown fat and white fat) were dissected, weighed and stored in individual vials. Both tissues and reference blood flow samples were digested using a solution of 4 M ethanolic KOH and 2% Tween 80 (Sigma, Taufkirchen, Germany) in a hot bath (50°C) overnight. At the end of the digestion period, the microspheres were recovered by the sedimentation method (Sampaio et al. 2003) and the dye was extracted in 4 ml of the organic solvent ethyl acetate. The fluorescence intensity of the dye was measured using a spectrophotometer (Cary Eclipse Fluorescence Spectrophotometer; Varian, Victoria, Australia). The following parameters were calculated: cardiac output (ml/min), cardiac index (ml/min/100 g), stroke volume (ml), total peripheral resistance (TPR, mmHg/ml/min/100 g), regional blood flow (ml/min/g), and regional vascular resistance (mmHg/ml/min/g), as described elsewhere [Gervais et al. 1999; Glenny et al. 1993].
Data analysis
Data are expressed as mean ± standard error. Statistical comparisons of mean values were performed using the Student’s unpaired t test. All p values less than 0.05 were considered significant.
Results
Systemic hemodynamics of WT and Mas-KO mice
Figure 1 displays the mean values of stroke volume, arterial pressure, cardiac index and TPR in 12- to 16-week-old urethane-anesthetized WT (n = 11) and Mas-KO (n = 10) mice. The Mas-KO mice had a significantly lower cardiac index (0.81 ± 0.08 ml/min/100 g) and stroke volume (0.03 ± 0.01 ml) than observed in WT mice (1.24 ± 0.23 ml/min/100 g and 0.05 ± 0.01 ml respectively; p < 0.05). Consequently, significantly higher TPR was observed in Mas-KO (103.6 ± 10.95 mmHg/ml/min/100 g) in comparison to WT mice (57.50 ± 13.05 mmHg/ml/min/100 g; p < 0.05). Mean arterial pressure (Mas-KO = 79 ± 2.0 mmHg and WT = 80 ± 2.0 mmHg) and heart rate (Mas-KO = 569 ± 16 beats/min and WT = 541 ± 16 beats/min) were similar between the two groups of mice. Similar results were observed in 36- to 44-week-old WT (n = 8) and Mas-KO (n = 8) mice (Figure 2). The weight of the younger animals was 24.4 ± 0.5 g for the WT mice and 25.5 ± 0.5 g for the Mas-KO mice. The weight of the older animals was 24.4 ± 0.7 g for the WT mice and 24.4 ± 0.5 g for the Mas-KO mice.

Stroke volume, mean arterial pressure, cardiac index and total peripheral resistance measured in 12-to-16 week-old urethane-anesthetized wild type (WT, n = 11) and Mas knockout (Mas-KO, n = 10) mice; *p < 0.05, Student’s t-test.
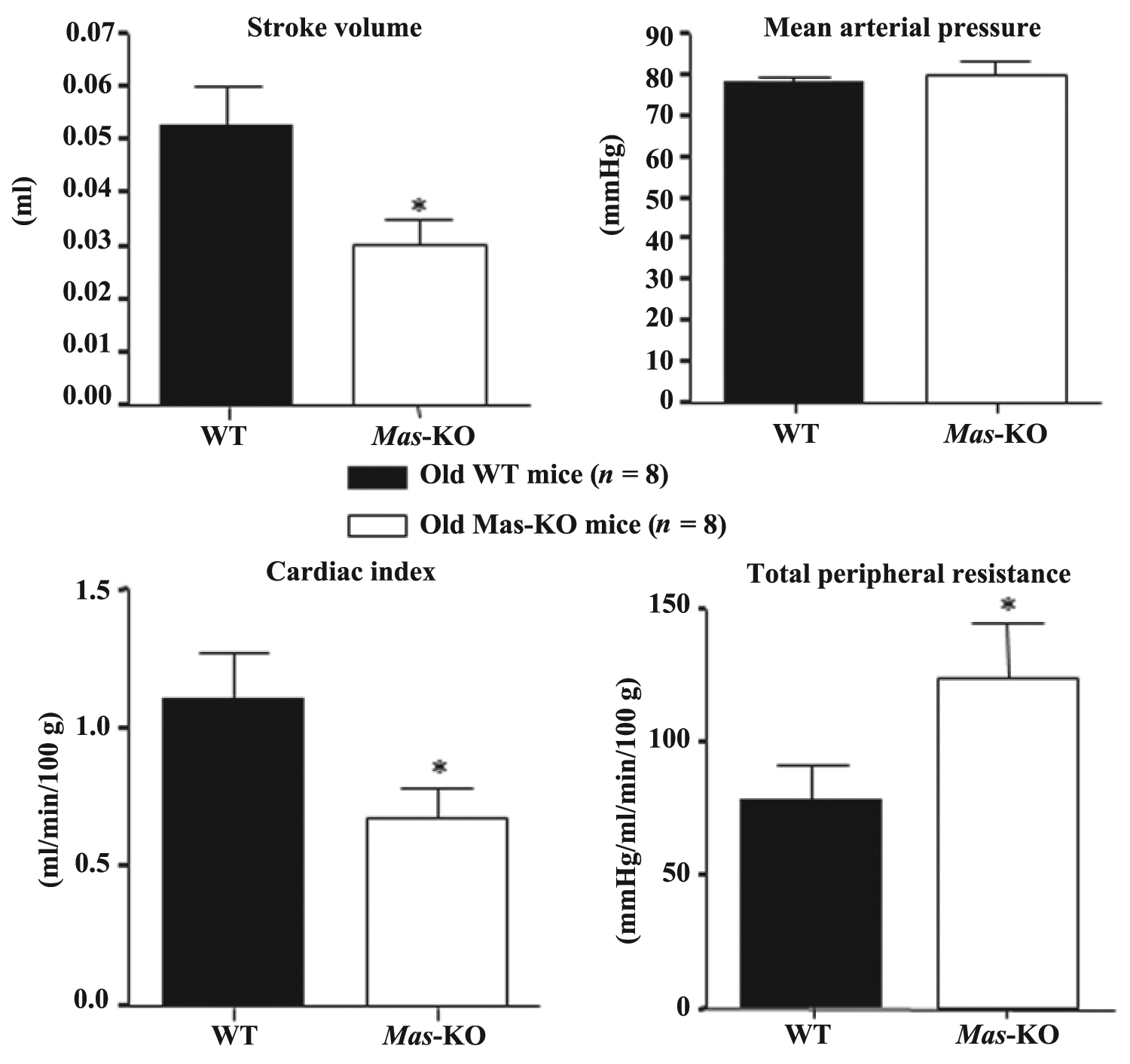
Stroke volume, mean arterial pressure, cardiac index and total peripheral resistance measured in 36-to-44 week-old urethane-anesthetized wild type (WT, n = 8) and Mas knockout (Mas-KO, n = 8) mice; *p < 0.05, Student’s t-test.
Regional blood flow and vascular resistance in WT and Mas-KO mice
Figure 3 and Table 1 display the values of vascular resistance and blood flow respectively in the different regions studied in 12- to 16-week-old and in 36- to 44-week-old WT and Mas-KO mice. Vascular resistance in the kidney, adrenal gland, lung, mesentery, spleen and brown fat was substantially greater in the 12- to 16-week-old Mas-KO mice in comparison to their age-matched controls. There were no statistically significant differences in vascular resistance in the left ventricle, skin, gastrocnemius muscle, brain, testis or white fat tissue (Figure 3). The changes in vascular resistance were paralleled by a decrease in blood flow in the kidney, lung, adrenal gland, mesentery, spleen and brown fat in the 12- to 16-week-old Mas-KO mice in comparison to control mice (Table 1). In 36- to 44-week-old Mas-KO mice, vascular resistance in the adrenal gland, brain, spleen, lung, testis, brown fat, mesentery and skin was significantly greater than that of the same organs in WT mice (Figure 3). The changes in vascular resistance were paralleled by a decrease in blood flow (Table 1). There were no significant differences in vascular resistance in other organs between the WT and Mas-KO mice. Comparison between young and older Mas-KO animals showed that only in brain, skin and in the testis territories the vascular resistance was higher in the older group. No differences were observed for central hemodynamic variables between the two groups.

Vascular resistance in kidney, adrenal gland, left ventricle, lung, skin, gastrocnemius muscle, mesentery, brain, spleen, testis, brown fat tissue and white fat tissue in 12-to-16 week-old urethane-anesthetized wild type (WT, n = 11) and Mas knockout (Mas-KO, n = 10) mice; *p < 0.05, Student’s t-test.
Regional blood flow in 12- to-16-week-old urethane-anesthetized WT and Mas-KO mice and in 36- to-44-week-old urethane-anesthetized WT and Mas-KO mice.

Values expressed as mean ± stand error. WT: young wild type (n = 11); young Mas-KO: Mas knockout (n = 10); old wild-type (n =8); old Mas-KO: Mas knockout (n = 8).
p < 0.05, Student’s t-test.
Discussion
A number of studies have demonstrated the participation of Ang-(1-7) in the regulation of cardiac function [Ferrario et al. 2005; Ferreira et al. 2002; Loot et al. 2002; Santos et al. 2000, 2005; Schmaier, 2003; Wang et al. 2010]. The results of the present study further indicate that Ang-(1–7) plays an important and effective role in the control of cardiac index and stroke volume. Mas-deficient mice exhibited significant lower cardiac index and stroke volume than WT mice. These alterations were present in both young and aged mice. This is in keeping with previous studies in which Ang-(1-7) has been reported to increase cardiac index and stroke volume in Wistar rats [Sampaio et al. 2003] and transgenic rats TGR(A1-7)3292, which exhibit a chronic increase in plasma Ang-(1-7) levels [Botelho-Santos et al. 2007]. Furthermore, the opposite changes in total peripheral resistance and cardiac index could explain the absence of substantial changes in blood pressure in the C57Bl/6 Mas-KO mice. Previous studies have reported that acute and chronic infusion of Ang-(1-7) in normotensive rats did not alter mean arterial pressure [Braga et al. 2002; Sampaio et al. 2003]. Moreover, Santos and colleagues also reported normal blood pressure in black C57Bl/6 Mas-KO mice [Santos et al. 2006].
The results of the present study reveal that Ang-(1-7) plays an important role in the control of blood flow distribution. Mas-KO mice exhibited significantly greater regional vascular resistance in the kidney, adrenal gland, lung, mesentery, spleen, testicle and brown fat. This is in keeping with the pronounced endothelial dysfunction described in Mas-KO mice [Rabelo et al. 2008; Xu et al. 2008] and with a large number of studies showing that Ang-(1-7) is a vasodilator peptide [Botelho-Santos et al. 2007; Bürgelová et al. 2009; Lemos et al. 2002; Osei et al. 1993; Porsti et al. 1994; Ren et al. 2002; Sampaio et al. 2003; Silva et al. 2007]. Although we did not investigate in the current study the mechanism of increased vascular resistance, it is probably related to the decreased NO production and increased reactive oxygen species production observed in these animals [Rabelo et al. 2008; Xu et al. 2008]. Accordingly, it is well documented that upon binding to endothelial cells, Ang-(1–7) can stimulate the production of NO, prostaglandins or endothelium-derived relaxing factor [Santos and Ferreira, 2007]. However, the relative contribution of each of these endothelium-derived factors to vasorelaxation induced by this peptide seems to vary with species and vascular territory [Botelho-Santos et al. 2007; Sampaio et al. 2003]. Another explanation for the increased peripheral vascular resistance observed in Mas-deficient animals could be an increase in Ang II or an increased expression or activity of AT1R due to the lack of Mas. However, in a recent study we could not detect any significant change in Ang II plasma levels in Mas-KO compared with WT mice [Guimarães et al. 2012]. Likewise, we found no alteration of Ang II binding in kidney slices of Mas-KO mice [Santos et al. 2003] or AT1R expression in the heart [Guimarães et al. 2012] or adipose tissue [Santos et al. 2008]. In addition AT1R are rather activated by Mas overexpression [Canals et al. 2006] although the AT1R signaling is antagonized by Mas [Kostenis et al. 2005]. The fact that the changes observed in Mas-deficient mice are essentially opposite to those produced by Ang-(1-7) infusion in rats [Sampaio et al. 2003] or in transgenic rats overexpressing an Ang-(1-7)-producing fusion protein [Botelho-Santos et al. 2007] further support a direct role of Ang-(1-7) in vascular resistance control.
Accumulating evidence suggests that Ang-(1-7) plays a significant role in renal function. The acute infusion of Ang-(1-7) and the chronic increase in Ang-(1-7) has been found to increase renal blood flow through the Mas receptor in anesthetized rats [Botelho-Santos et al. 2007; Sampaio et al. 2003] and to produce afferent arteriolar relaxation through specific receptor-mediated NO release in isolated rabbit kidneys [Ren et al. 2002]. However, van der Wouden et al. (2006) found no effect of Ang-(1-7) on renal blood flow in freely moving rats. Factors such as anesthesia may account for the differences observed. Furthermore, differences were not found in vascular resistance in the renal territory of older Mas-KO and WT mice in our study, suggesting a variation in the renal vascular territory with the ageing process, such as the stiffening of renal arterioles or decreased responsiveness to vasodilator agents.
Alenina and colleagues demonstrated that the testis is an important site for the expression of Mas receptor mRNA [Alenina et al. 2002]. Moreover, TGR rats which express an Ang-(1-7)-producing fusion protein in the testis have been found to exhibit a lower vascular resistance and greater testicular blood flow than SD rats [Botelho-Santos et al. 2007]. We observed in the present study that the testis vascular resistance of young Mas-deficient mice was comparable to that of WT mice of the same age, whereas vascular resistance in this organ in older Mas-KO mice was greater than that found in controls. This is particularly important considering that blood flow through the testicular microvasculature is considered to be important for normal testis function as it regulates interstitial fluid dynamics which is as important intratesticular transport medium [Welsh et al. 2010]. A decrease in testicular function is already present in 10-week-old Mas-KO mice [Leal et al. 2009]. Whether the deterioration of the testis blood flow in older Mas-KO mice would further impact testicular functions remains to be clarified.
A number of studies have reported the vasodilator effect of Ang-(1-7) in the adrenal gland [Botelho-Santos et al. 2007; Ehrhart-Bornstein et al. 1998; Mulrow, 1998; Sampaio et al. 2003]. Ang-(1-7)-induced relaxation was found to be eliminated by the removal of endothelial cells and inhibited by the NO synthase inhibitor N-nitro-L-arginine methyl ester (L-NAME) in bovine adrenal cortical arteries, suggesting that Ang-(1-7) relaxation occurs through the activation of Ang-(1-7) receptors and is mediated by endothelial cell NO [Gauthier et al. 2008]. In the present study we observed that Mas-KO mice exhibited greater adrenal vascular resistance than control mice, thereby confirming the participation of the Mas receptor in the regulation of adrenal vascular tone.
The existence of a local renin–angiotensin system in the lungs has recently been demonstrated [Kuba et al. 2006; Orte et al. 2000; Uhal et al. 2011]. Pulmonary arterial hypertension is reported to be caused by functional and structural changes in the pulmonary vasculature, leading to increased pulmonary vascular resistance [Orte et al. 2000]. Moreover, Ang II is involved in this process of vascular remodeling through AT1 receptors [Morrell et al. 1999]. Here we show that Mas-KO mice exhibited greater vascular resistance than control mice in the pulmonary vascular territory. This is in keeping with an early report showing that chronic increase in plasma Ang-(1-7) levels reduced the vascular resistance for the lung [Botelho-Santos et al. 2007]. Although further studies are needed to assess the participation of Ang-(1-7) in the regulation of pulmonary vascular tone, our data and the recent observations that treatment with Ang-(1-7) [Shenoy et al. 2011; Watts et al. 2011] or ACE2 activation [Shenoy et al. 2011] can reduce pulmonary hypertension illustrate the importance of this heptapeptide in the lung vascular territory.
The older Mas-KO mice exhibited greater vascular resistance in the brain than control mice of the same age. This observation is in agreement with previous studies that report a vasodilator effect of Ang-(1-7) in the cerebral vascular territory [Botelho-Santos et al. 2007; Sampaio et al. 2003], in piglet pial arterioles [Meng and Busija, 1993] and in canine middle cerebral arteries [Feterik et al. 2000]. However, in this study, we did not observe changes in cerebral blood flow of young Mas-KO mice. Whether the lack of alteration in brain vascular resistance in young Mas-KO mice compared with old Mas-KO mice may reflect an increased dependency of vascular brain Mas expression with the ageing process remains to be clarified.
The discovery of a local adipose tissue RAS has allowed the detection of a pathophysiological role for RAS in obesity-associated disturbances and insulin resistance, as well as a function in the secretion of adipocyte-derived products [Engeli et al. 2003]. Moreover, Ang I, Ang II and Ang-(2-8) have been detected in rat brown adipose tissue (BAT) [Engeli et al. 2003; Shenoy and Cassis, 1997], suggesting endogenous production, especially of Ang II. We recently found a marked difference in BAT blood flow between TGR and SD rats, suggesting an effect of Ang-(1-7) on the metabolic rate of BAT. In agreement with this result, the BAT of Mas-KO mice exhibited greater vascular resistance and lesser blood flow in comparison to WT mice, suggesting the participation of the Mas receptor in the regulation of vascular tone in this tissue.
The absence of the Mas receptor caused an increase in vascular resistance in the spleen. In a previous study we demonstrated that a chronic increase in Ang-(1-7) causes a reduction in vascular resistance in the same organ [Botelho-Santos et al. 2007]. However, no alteration was found in vascular resistance in the spleen when Ang-(1-7) was administrated in the acute form [Sampaio et al. 2003]. This result suggests that the participation of the Mas receptor in this vascular bed is mediated by mechanisms dependent upon a long-term change in Ang-(1-7)/Mas, modulating endothelial NO synthase or similar vasodilator mechanisms. However, further studies are needed to clarify this point.
Ang-(1–7) increased mesentery blood flow. Oliveira and colleagues demonstrated that Ang-(1–7) causes both vasodilatation and bradykinin potentiation in mesenteric arterioles which was blocked by A-779, N-nitro-L-arginine methyl ester and indomethacin, suggesting important participation of local prostanoids and NO in the action of Ang-(1–7) [Oliveira et al. 1999]. The same authors demonstrated that Ang-(1–7) potentiates the bradykinin vasodilator effect in mesenteric arterioles in spontaneously hypertensive rats through the release of prostanoids [Fernandes et al. 2001]. Accordingly, we observed greater vascular resistance in the Mas-KO in this territory, suggesting that endogenous Ang-(1-7)/Mas-related mechanism plays a role in the modulation of blood flow in this territory.
In contrast with most territories, the vascular resistance in the heart and skeletal muscle was not statistically different from that of WT mice, although the tendency was a reduction, especially in the heart. This is in keeping with the absence of changes in the coronary flow in TG-L3292 rats which present a chronic increase in plasma Ang-(1-7) levels [Botelho-Santos et al. 2007]. The observation in the heart can be at least partially related to the complex interaction of Mas with AT1R and AT2R in the mouse heart circulation [Castro et al. 2005]. Only in the presence of AT1R blockade with losartan, Ang-(1-7) produced a decrease in the perfusion pressure which was blocked by the Mas antagonist, A-779. However, in the presence of an AT2R antagonist, PD123319, Ang-(1-7) produced an increase in perfusion pressure which was diminished but not abolished by losartan and not affected by A-779 [Castro et al. 2005]. These observations are in keeping with several studies describing vasodilation [Brosnihan et al. 1996; Porsti et al. 1994], vasoconstriction [Kumagai et al. 1993; Neves et al. 1997], or even absence of effect [Almeida et al. 2000; Gorelik et al. 1998] depending on the dose of Ang-(1-7), in the coronary circulation of rodents. It is important to note that the relatively normal coronary blood flow in Mas-KO mice indicates that the changes in ventricular contractility observed in these animals are mainly a result of the absence of Mas in cardiomyocytes in which Ang-(1-7)/Mas appears to play a major role [Gomes et al. 2012; Santos et al. 2008]. Whether similar mechanisms are present in skeletal muscle myocytes or blood vessels is not known.
In this and former studies [Botelho-Santos et al. 2007; Sampaio et al. 2003] we have used anesthetized animals, mainly because of technical limitations. This could interfere with the variables analyzed. However, although future studies using awake animals would be important to perform, our data indicate an important role for Ang-(1-7)/Mas in regional blood flow distribution, at least in anesthetized animals. Of note is the fact that with few exceptions (brain, skin and testis) the increase in vascular resistance was similar in young and older mice, suggesting that the consequences of Mas deficiency were not dependent on age-related changes in vascular function.
In summary, the results of the present study indicate that Mas is importantly involved in the regulation of regional and systemic hemodynamics in anesthetized mice. More importantly, the results indicate that the hemodynamic changes producing Mas deficiency are sustained, suggesting a formerly unsuspected important physiological role of Ang-(1-7)/Mas in the tonic control of blood flow. Furthermore our current results are in keeping with regional variations in the role of Ang-(1-7)/Mas in vascular tonus modulation, which may have important clinical implications for cardiovascular therapeutics.
Footnotes
Funding
This work was supported by FAPEMIG and CNPq. Programa de Pós-Graduação em Fisiologia e Biofísica, UFMG.
Conflict of interest statement
The authors declare no conflicts of interest in preparing this article.