
Abstract
The aim of this study was to evaluate the effect of the miR-301a/PTEN pathway in cervical cancer. miR-301a and PTEN expression were measured by quantitative real-time PCR (qRT-PCR) in tissues samples and HeLa cells. PTEN protein level was determined by Western blotting. Dual reporter luciferase assay was performed to validate PTEN as a direct target of miR-301a. The gain- and loss-of function assay was performed by miR-301a overexpression and silencing. Cell proliferation was monitored by cell counting Kit-8 (CCK-8). Cell apoptosis was quantitated by flow cytometry. SPSS was used to analyze the significant difference in the treatments. miR-301a demonstrated a significantly higher expression in cervical carcinoma tissues compared with the paired non-carcinoma tissues (n = 12), while PTEN expression was found to be significantly lower in cervical carcinoma tissues than their paired non-carcinoma tissues (n = 12). In addition, PTEN was identified as the direct target of miR-301a. Moreover, overexpression of miR-301a significantly promoted HeLa cells proliferation and anti-apoptosis which had a reverse pattern after PTEN overexpression. Our results confirm PTEN as a direct target of miR-301a in HeLa cells and suggest that miR-301a/PTEN pathway contributes to the development and progression of cervical cancer.
Keywords
Introduction
Among females worldwide, cervical cancer remains one of the leading causes of cancer death. It represents an infectious disease that is accompanied with gene-involved multiple steps and more complex biological processes. 1 Patients identified during the early stages of cervical cancer can be successfully treated; but there is a deficit in therapeutic efficacy for patients that are diagnosed at the late or metastatic stages. Although tremendous advances have been made in the diagnosis and treatment of cervical cancer over the past years, there is still an urgent need for biomarkers and targets available for predicting the prognosis of this cancer. Therefore, a better understanding of the molecular mechanisms underlying the development and progression of cervical cancer is crucial for treatment strategies of this disease.
MicroRNAs (miRNAs) are short non-coding RNAs (ncRNAs) of ∼22 nucleotides in length that mediate gene expression by binding the 3′ untranslated region (3′-UTR) of targeted mRNAs. 2 They have been revealed to target a wide spectrum of genes and de-regulate their functions which play important roles in cancer progression. 3 Currently, the research of miRNA regulation of gene expression has become a hot spot in cancer research. Thus, investigation on the expression, biological roles and underlying mechanisms of miRNAs in cervical cancer might provide valuable therapeutic targets in the treatment of patients with this disease. However, the effect of miR-301a/PTEN pathway on the proliferation and apoptosis of cervical cancer still has not yet been elucidated.
In this study, we observed that miR-301a was significantly up-regulated, while the phosphatase and tensin homolog (PTEN) was significantly down-regulated in the cervical carcinoma tissues. In addition, we also identified PTEN as a direct target of miR-301a. Next, we showed that overexpression of miR-301a leads to increased cell proliferation and an-apoptosis in HeLa cells. Furthermore, PTEN overexpression showed a reverse effect on cell proliferation and apoptosis compared with miR-301a. Our results provide the first evidence of regulatory mechanisms of miR-301a/PTEN pathway in cervical cancer. Hence, our data indicate that miR‐301a could be a valuable novel candidate therapeutic target for cervical cancer.
Materials and methods
Ethical approval
The study was approved by the Institutional Ethics Committee of our Liaocheng People’s Hospital, and written informed consent was obtained from all participants.
Cell line
HeLa cells were purchased from
qRT-PCR
Total RNAs were extracted from cells and tissues using TRI Reagent (Molecular Research Center, Cincinnati, OH). Residual DNA was removed using DNA-free DNase (Ambion, Austin, TX). RNA measurements were quantitatively performed using nanodrop spectrophotometry (Thermo Fisher, Waltham, MA). Reverse transcription was performed using Moloney murine leukemia virus reverse transcriptase (Invitrogen, Carlsbad, CA). miR-301a and gene quantitation was determined by TaqMan™ analysis run on a QuantStudio 6 Flex PCR system (Thermo Fisher). 4 qRT-PCR primers for gene and miR-301a expression were commercially available from Applied Biosystems (Foster City, CA). GAPDH and U6 small nuclear RNA (snRNA) were used as internal controls for gene and miRNA quantitation, respectively. Gene or miRNA quantitation was determined as previously described.5,6 All independent PCR-based reactions were performed in triplicate.
Cell transfection
Transfections of miRNA mimics miR-301a alongside the mirVana™ miRNA mimic Negative Control #1 (miR-Ctl) utilizing the RNAiMAX Reagent (Invitrogen) were performed according to manufacturer protocols (Carlsbad, CA). The PTEN expression plasmid (SC119965) was obtained from OriGene Technologies Inc. (Rockville, MD). The HeLa cells were plated in 6-well plates (1 × 105 cells per well), and were transfected with or without PTEN plasmid and Mock plasmid vector using Lipofectamine 2000 (Invitrogen) as previously described, 1 as was miR-301a silencing. 7 HeLa cells were transfected with LNA-anti-miR-301a or LNA-anti-control (Exiqon, Woburn, MA) by using Lipofectamine RNAiMAX reagent. After 48 h, the cells were collected, and qRT-PCR was performed to ensure silencing was achieved. The LNA-anti-control (anti-miR-ctl) was the miRCURY LNA miRNA inhibitor control Negative Control A (Exiqon, Woburn, MA) which has not more than 70% homology to any sequence in the mouse genome. 7
Western blotting
Whole cells were lysed in lysis buffer (Thermo Fisher), and protein concentrations were determined using a DC Protein Assay kit (Bio-Rad, Hercules, CA). Total protein lysates were fractionated on 4–15% polyacrylamide gels and transferred onto nitrocellulose (Bio-Rad). As primary Abs were used: anti-PTEN at 1:500 dilution (1:500; bsm-52192R, Bioss, Woburn, MA) and anti-GAPDH (1:500; G9545 Sigma-Aldrich). After being washed with TBST three times, the membranes were incubated with HRP-conjugated secondary Abs (Bio-Rad) for 1 h. All protein experiments were performed in triplicate. Separate blots were used for each independent experiment to avoid problems related to incomplete membrane stripping. The blotting bands were quantified by ImageJ.
Cell proliferation
CCK-8 allows sensitive colorimetric assays for the determination of cell viability in cell proliferation, which is based on WST-8 which is reduced by dehydrogenases in cells to give an orange colored product (formazan). The amount of the formazan in cells is directly proportional to the number of living cells. HeLa cells were seeded at a density of 5000 cells per well in the 96-well plates. CCK‐8 assay (Dojindo Molecular Technologies, Gaithersburg, MD) was performed to measure cell proliferation at the indicated time according to manufacturer protocol.
3′-UTR luciferase assay
HeLa cells grown in a 48-well plate were co-transfected with PTEN 3′-UTR luciferase reporter construct and with miR-301a mimic or mirVana™ miRNA mimic Negative Control # 1 using Lipofectamine 2000 (Invitrogen) with Opti-MEM (Thermo Fisher) as previously described. 8 After 48 h post-transfection, the cells were lysed for measurement of luciferase activities using the Duo-Luciferase kit 2.0 (GeneCopoeia, Rockville, MD) according to the manufacturer's instructions. Relative luciferase activity was calculated by normalizing Firefly to Renilla.
Flow cytometry
The cells were collected, and the apoptotic or dead cells were detected by staining with annexin V-FITC and propidium iodide by an annexin V-FITC apoptosis detection kit (BioVision, Mountain View, CA) according to the manufacturer's instructions. Each experiment was performed in triplicate.
Statistics
The statistical analyses were executed by statistical software (SPSS17.0 for Windows; SPSS, Inc., Chicago, IL). Two-tailed, unpaired Student’s t-test assessed significance between two conditions. The one-way ANOVA was used to compare the significant difference in more than two conditions with Tukey’s post-hoc test. The correlation between relative miR-301a expression (normalized to U6) and the relative expression of PTEN mRNA (normalized to GAPDH) was analyzed using Pearson correlation coefficient analysis with r and P values. Significance was accepted at P < 0.05. Data were expressed as mean ± SE from at least three independent experiments.
Results
Up-regulation of miR-301a expression in human cervical carcinoma tissues
Previous studies reported that miR-301a functions as an oncogene in many human cancers, however, there are limited data regarding miR-301a and cervical cancer. 9 In this study, we first evaluated the expression level of miR-301a in randomly selected 12 paired cervical carcinoma tissues and their matched non-carcinoma tissues by qRT-PCR. We observed that miR-301a was significantly up-regulated by 2.5-fold in the human cervical carcinoma tissues compared with the non-carcinoma tissues (Figure 1). These results suggest that up-regulation of miR-301a may be involved in the tumor development and progression.
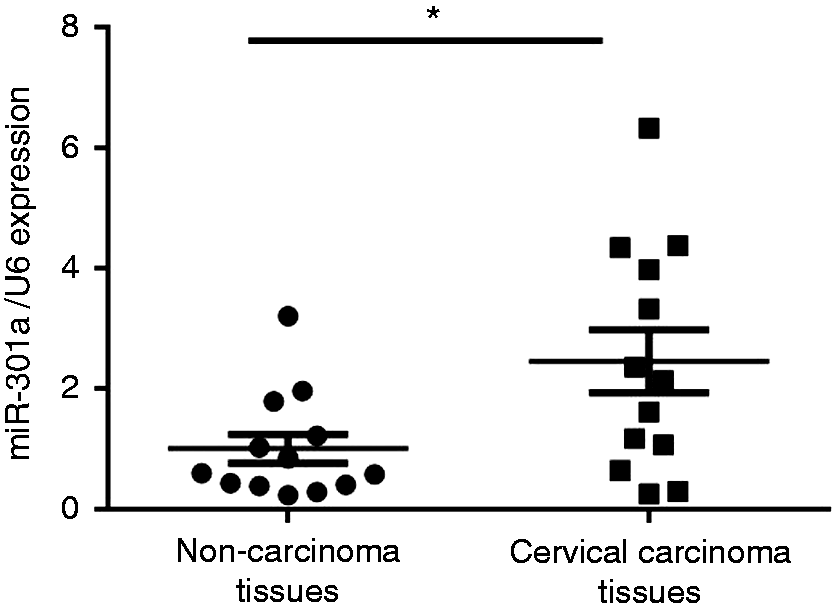
miR-301a is overexpressed in cervical cancer. qRT-PCR verified the miR-301a level in a separate random cohort of cervical carcinoma tissues compared to paired non-carcinoma tissues (n = 12). Bars represent the mean ± SE, *P < 0.05.
PTEN as a direct miR-301a target in HeLa cells
In the previous study, miR-301a had been revealed to directly target PTEN in Ewing's sarcoma cell lines.9,10 To evaluate the possible mechanistic involvement of miR-301a and to reveal any correlation between miR-301a and PTEN in cervical carcinoma, we subsequently detected the expression of PTEN in cervical carcinoma tissues and their matched non-carcinoma tissues (n = 12) by qRT-PCR. PTEN down-regulation was observed in the cervical carcinoma tissues compared with the non-carcinoma tissues (Figure 2a). Moreover, PTEN expression levels were reversely correlated with miR-301a (Figure 2b). To further determine whether miR-301a expression affected endogenous PTEN expression, we transfected the miR-301a mimic as well as the negative miR-Ctl into HeLa cells. We observed an increased miR-301a expression by 7.2-fold compared with miR-Ctl (P < 0.05) (Figure 2c), and decreased PTEN expression at the mRNA and protein levels (Figure 2d and e). In addition, the 3′-UTR luciferase assay confirmed that miR-301a directly binds PTEN mRNA in HeLa cells (Figure 2f).
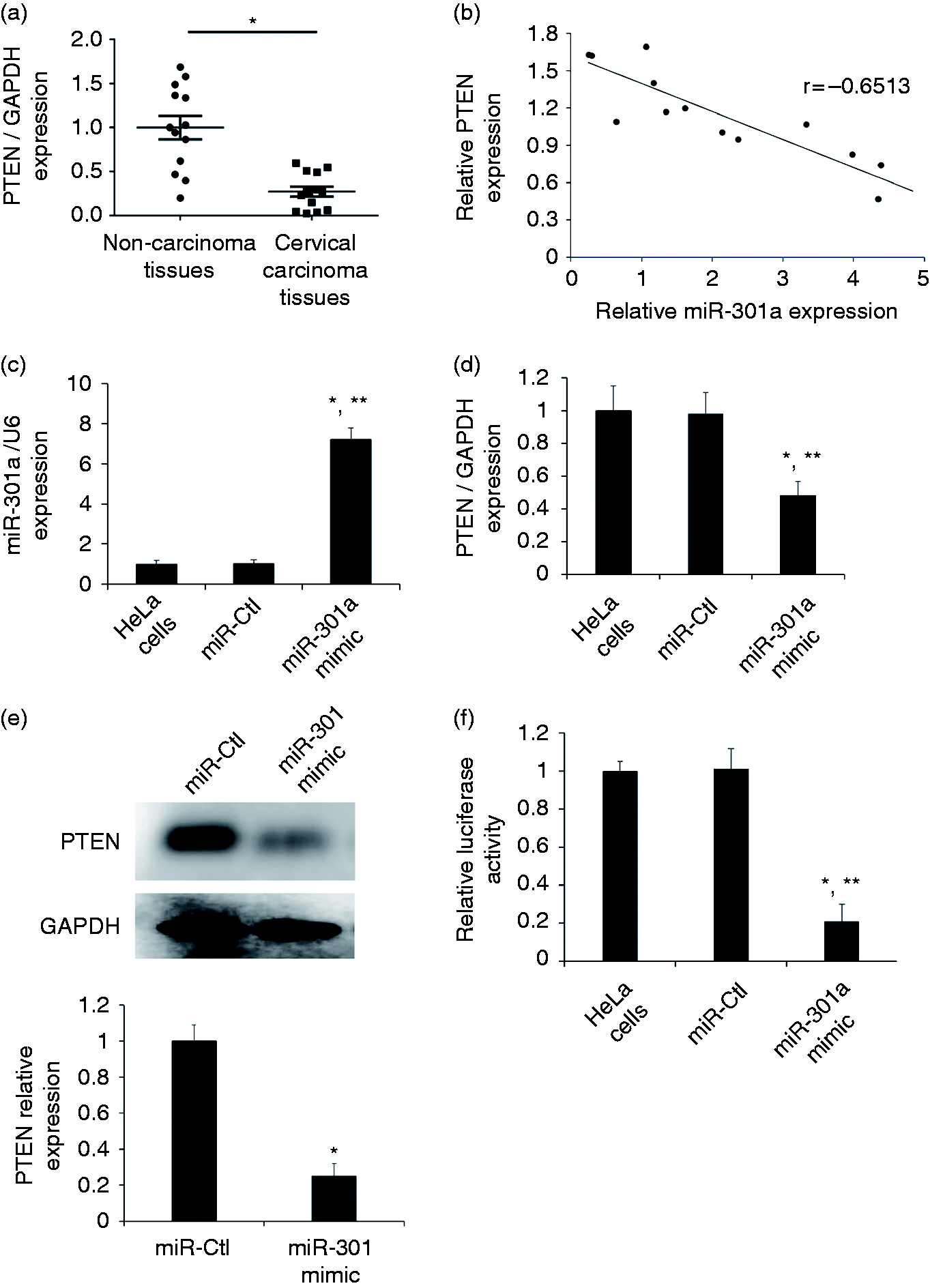
PTEN is directly suppressed by miR-301a. (a) qRT-PCR verified the PTEN expression in a separate random cohort of cervical carcinoma tissues compared to paired non-carcinoma tissues (n = 12). Bars represent the mean ± SE, *P < 0.05. (b) Correlation analysis of the relative expression of miR-301a and PTEN in cervical carcinoma tissues (Pearson correlation coefficient = −0.6513). (c, d) miR-301a and PTEN expression levels were determined by qRT-PCR in HeLa cells transfected with miR-Ctl or miR-301a mimic. Bars represent the mean ± SE, *P < 0.05 vs HeLa cells; *P < 0.05 vs miR-Ctl. (e) Western blotting showed PTEN protein level after miR-301a or miR-Ctl overexpression in HeLa cells. The blotting bands were quantified by ImageJ. The relative PTEN expression levels were normalized to GAPDH. Results are mean ± SE (n = 3). *P < 0.05 vs miR-Ctl. (f) 3′-UTR luciferase reporter assay was performed to confirm that miR-301a bound to specific regions in the PTEN 3′-UTR. Bars represent the mean ± SE, *P < 0.05 vs HeLa cells; *P < 0.05 vs miR-Ctl.
Effect of miR-301a overexpression/silencing and PTEN expression on the proliferation and anti-apoptosis of HeLa cells
PTEN is well known for playing important roles in the regulation of proliferation and apoptosis.11–13 Since transfection of miR-301a resulted in the reduction of PTEN expression, we next evaluated the effects of miR-301a/PTEN on the proliferation and apoptosis of HeLa cells. The cell growth of HeLa was promoted by the transfection of miR-301a overexpression as determined by cell counting in comparison with negative miR-Ctl transfected cells (Figure 3a). For those cells overexpressed miR-301a, the amount of apoptosis was significantly decreased compared to negative miR-Ctl transfected cells (Figure 3b). In support, mRNA levels of anti-apoptotic factors (BCL2 and MCL1) increased in HeLa cells overexpressing miR-301a while levels of pro-apoptotic factors (BAD and BAX) decreased (Figure 3c). We also observed opposite patterns of decreased cell proliferation and increased apoptosis after miR-301a silencing (Figure 3d to g). Furthermore, PTEN overexpression gave also decreased cell proliferation and promoted apoptosis in HeLa cells, which is a similar pattern as in miR-301 silencing (Figure 3h to j).
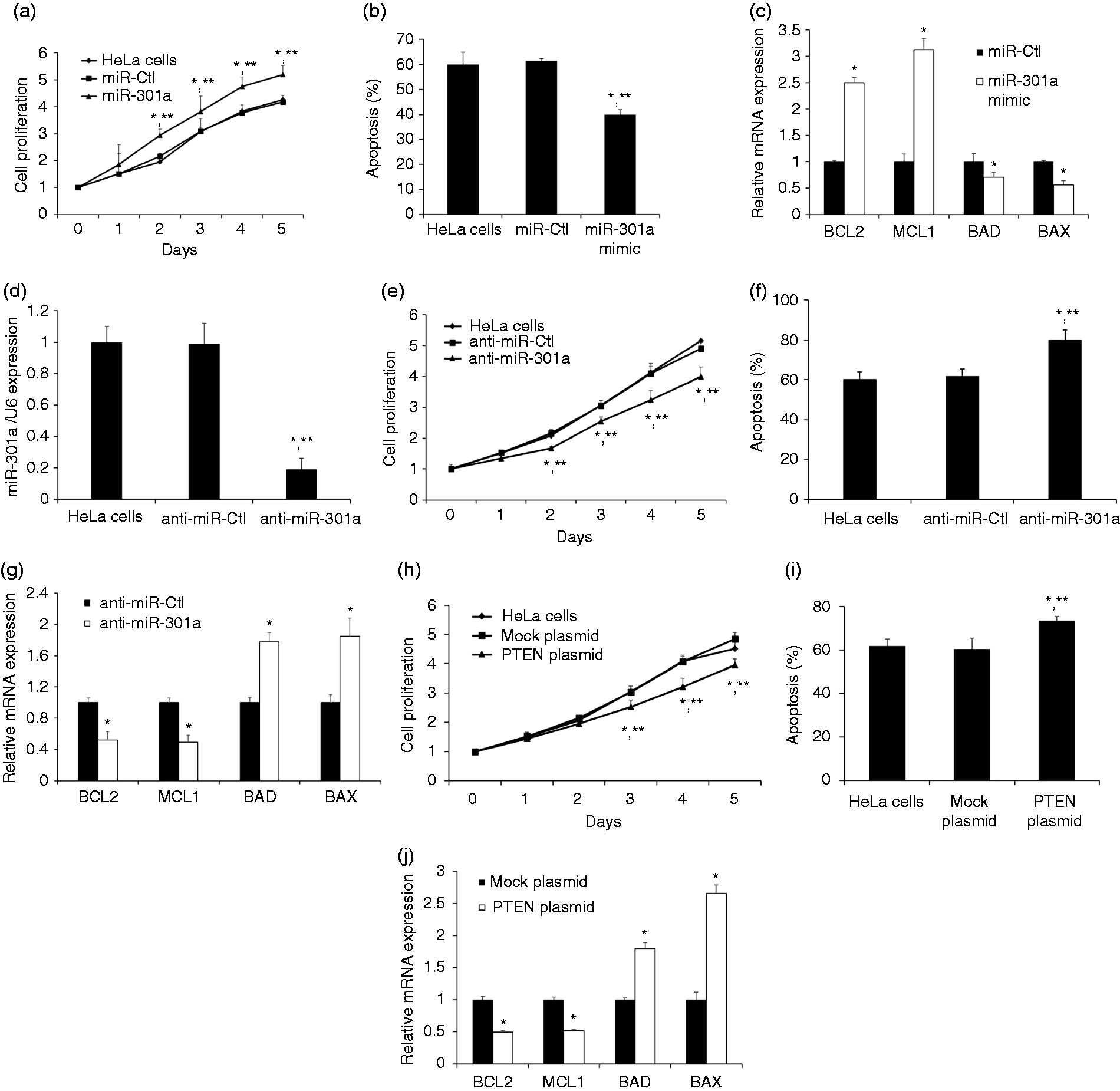
Biologic effects of miR-301a or PTEN overexpression in HeLa cells. (a) miR-301a overexpression promoted proliferation in HeLa cells. Data are expressed as relative fold change compared with d 0. (b) Annexin V flow cytometry measured levels of apoptosis after miR-301a overexpression. (c) Transcript analysis showed that levels of anti-apoptotic factors (BCL2, MCL1) increased while pro-apoptotic factors (BAD, BAX) decreased after miR-301a overexpression. (d) HeLa cells were transfected with anti-miR-ctl and anti-miR-301a. After 48h post transfection, miR-301a was silenced by anti-miR-301a. (e) miR-301a silencing suppressed proliferation in HeLa cells. Data are expressed as relative fold change compared with d 0. (f) Annexin V flow cytometry measured levels of apoptosis after miR-301a silencing. (g) The expression level of anti-apoptotic factors (BCL2, MCL1) decreased while pro-apoptotic factors (BAD, BAX) increased after miR-301a silencing. (h) PTEN overexpression inhibited proliferation in HeLa cells. Data are expressed as relative fold change compared with d 0. (i) Annexin V flow cytometry measured levels of apoptosis after PEN overexpression. (j) Transcript analysis showed that levels of anti-apoptotic factors (BCL2, MCL1) decreased while pro-apoptotic factors (BAD, BAX) increased after miR-301a overexpression. Data represents the mean ± SE, *P < 0.05 vs HeLa cells; *P < 0.05 vs miR-Ctl, anti-miR-ctl or mock plasmid.
Discussion
miRNAs are the most studied class of small ncRNAs with approximately 22 nucleotides. By binding the 3′-UTR of specific coding transcripts, miRNAs cause degradation or translation inhibition of their target genes and affect a multitude of biological processes, such as proliferation and apoptosis. The expression of miRNAs is tightly controlled, and the expression dysregulation of miRNAs has been revealed to be linked to cancer. 14 It has been reported that miR-301a is a negative prognostic marker in many human cancers related to high metastatic potential. 15 miR-301a has been observed to promote colorectal cancer cell growth and invasion by targeting SOCS6. 16 In addition, transfection of miR-301a inhibitor significantly inhibited growth and enhanced apoptosis in malignant melanoma cells. 10 However, the role of miR-301a in the cervical cancer has not been completely elucidated yet. In the present study, miR-301a was found to be up-regulated in cervical carcinoma tissues compared to the paired non-cervical carcinoma tissues. Moreover, the overexpression of miR-301a could promote the proliferation and anti-apoptosis of cervical carcinoma cells. The miR-301a silencing reversed the effect of miR-301a overexpression on cell proliferation and apoptosis.
The PTEN gene has been revealed to participate in a myriad of physiological and pathological processes. It is also one of the most frequently mutated tumor suppressors in human cancer. Functions for PTEN have been identified in the regulation of many cell processes, including growth, adhesion, migration, invasion and apoptosis.11,13,17 PTEN has been found to be involved in the regulation network of proliferation and apoptosis of colorectal cancer cells. 11 In addition, it has been observed that miR-21 promoted cell proliferation and inhibited apoptosis by PTEN signaling pathway in esophageal cancer. 18 Moreover, miR-301a has been found to promote cell proliferation via PTEN targeting in Ewing’s sarcoma cells. 1 Since PTEN has been revealed to be directly regulated with miR-301a in Ewing’s sarcoma cells and miR-301a was observed to be up-regulated in cervical carcinoma tissues, it might be rational to propose that miR-301a could potentially play a role in human cervical carcinoma through PTEN. In this study, PTEN was found to be downregulated in cervical carcinoma tissues. In addition, PTEN was also identified as a direct target of miR-301a by 3′-UTR luciferase assay and Western blotting. Furthermore, PTEN overexpression exhibited opposite patterns of decreased cell proliferation and increased apoptosis compared with miR-301a overexpression, which is similar with the miR-301a silencing. Future efforts will be directed towards developing an in vivo model to verify the in vitro results.
In summary, our study provides functional evidence that miR-301a/PTEN pathway is able to regulate the cell proliferation and apoptosis in the HeLa cells in vitro. This newly identified miR-301a/PTEN pathway suggests the potential to be a prognostic marker and benefits targeted therapies for patients with cervical carcinoma in the future.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.