
Abstract
Innate immunity natural Abs (NAbs) execute a number of functions, including protection and surveillance. Despite active research, the stimuli that induce the formation of NAbs are still described only hypothetically. Here, we compared repertoires of anti-glycan Abs in the peripheral blood of mice that received per os various bacteria. The repertoires of Abs of mice primed in this way were compared using a microarray that included about 350 glycans, as well as 150 bacterial polysaccharides. Sterile mice did not possess anti-glycan Abs. Oral inoculation of a single strain or combination of two to four strains of bacteria, as well as putting the animals on short-term nutrition with non-sterile food, did not contribute significantly to the formation of Abs, whereas a single gavage of digested food of non-sterile mice induced the formation of a repertoire close to the natural ones. Interestingly, the priming with polysaccharide Ags (in a composition of the bacterial cell envelope), that is, dominant Ags of bacteria, led to the induction of Abs against typical glycans of mammalian glycoproteins and glycolipids (e.g. Abs of the ABH blood group system) that do not have a structural similarity to the polysaccharides. The results support the importance of early contact with a naïve immune system with microorganisms of the environment to form a normal NAbs repertoire.
Keywords
Introduction
Innate immunity is of paramount importance at the early stages of mammals' development. The moment of birth becomes a starting point of immune system development because the conditions of the limited Agic stimulation in utero turn into an environment rich with Ags. Natural Abs (NAbs) as a part of innate immunity are present in the blood of mammals at an early age and demonstrate a stable level and repertoire throughout life.1,2
The difference between NAbs and adaptive Abs, induced in response to Ag invasion, was first mentioned when the ABO blood group system was discovered. 3 Since then some of the other NAbs directed to blood group Ags, xeno-Ags and malignant cells have been found and partly characterized.4–8
NAbs mainly belong to the IgM class, have low affinity, and are believed to be polyreactive immunoglobulins1,7; nevertheless, some of them demonstrate precise specificity, as mentioned above for anti-A and anti-B. Similar to adaptive Abs, broadly reactive NAbs protect the organism, but have a number of distinctive features. NAbs are the first line of defense from infection because of their permanent presence in the organism (without canonical immunization) and can respond to foreign agents much faster. In mice, a 2- to 7-d period is required to produce high-specificity adaptive Abs – this is more than enough for infection to spread due to the high speed of viral and bacterial propagation. NAbs also perform the clearance of metabolites, i.e. utilization of cell degradation products (including the excessive or pathogenic proteins) and the elimination of transformed cells.9–11
NAbs formation is poorly understood, despite decades of research. Several key points have been suggested,12–16 particularly that NAbs appear to result from enterobacterial stimulation. Many factors can affect the formation of Abs directly or indirectly, including nutrition. Nutrition components are substrates for many microorganisms that are involved in the digestion of food and the synthesis of vital compounds. A normal microbiota is required for defense against pathogens by competing for limited resources. In the presence of normal microflora, which is relatively stable during the lifetime and contains more than 400 species, pathogens remain within the transient microflora.17,18
NAbs play an important role in childbirth, that is during the transition from relatively sterile intrauterine conditions to an environment rich in Ags. The first days and weeks of life represent a crucial window of opportunity for shaping the development of the gastrointestinal tract and immune system, as well as for the adult microbiota. The study of the immune system at the level of cells and immunoglobulins in humans, especially young children, is difficult not only because of the risk of harm to health, but also because of the complexity of immunity. A useful model, apparently, is that of sterile animals whose living conditions can be controlled and varied. Adjusted for obvious differences between the human and mouse organisms, the results can be informative and useful. Little information about NAbs in mice is available, despite the fact that NAbs' producers (B1 cells) were first discovered and characterized in mice, 19 particularly data on Abs directed to auto-Ags (DNA, proteins, membrane oxidation-caused epitopes).20–22 To our knowledge, a systematic study of the anti-glycan Abs repertoire in mice has not been carried out yet. Presumably, as in humans, the murine repertoire of anti-glycan Abs is broad, because the environmental conditions (microbiota and nutrition) are similar to those of humans.
In this study, we challenged sterile mice with bacteria per os, and examined the specificity and comprehensiveness of the raised repertoires of Abs that are believed to be NAbs.
Materials and methods
Serum samples
The repertoire of anti-glycan Abs in sera of male Swiss Webster mice at the age of 3–4 mo was studied. Animals were obtained by Cesarean section and housed in special isolators as described previously. 23 Each group (see Supplement 1) of animals included four mice. Group #1: sterile conditions were kept; group #2: contact to Escherichia coli K-12 strain W 3110; group #3: contact to Bifidobacterium longum strain NCC2705 (Nestec Ltd., Nestlé Research Center Lausanne, P.O. Box 44, CH-1000 Lausanne 26); group #4: contact to Bacteroides thetaiotaomicron strain VPI-5482 (ATCC 29148); group #5: contact to Lactobacillus reuteri strain SD2112 (ATCC 55730); group #6: contact to four strains of bacteria (E. coli strain W3110, B. longum strain NCC2705, B. thetaiotaomicron strain VPI-5482, L. reuteri strain SD2112); group #7: transferred to normal diet (standard granulated food); group #8: gavaged orally with a sample of non-sterile mice feces; group #9: conventional controls, CD-1 male mice fed with standard granulated food.
All mice from groups #1–6 were fed with sterilized food (autoclaved chow diet, LabDiet).
Mice from groups #2–5 were given a single 200-µl dose (108 CFU/mouse) of an overnight bacterial culture. Conventional controls were born normally and reared with their mothers until 9–14 wk of age. Blood samples were obtained 28 d after the exposure in all experiments. Animals were handled in strict accordance with good animal practice as defined by the relevant local animal welfare bodies; all animal procedures were supervised and approved by Food for Health Science Centre, Lund University ethics committee for animal experimentation.
Environmental endotoxins were not tested in the material used to prime Abs.
Microarray chips
Microarrays were prepared from 341 different synthetic amine-functionalized glycans and 146 bacterial polysaccharides, using N-hydroxysuccinimide-derivatized glass slides (slide H, Schott-Nexterion, Mainz, Germany), as described previously. 24 The glycan library included blood group Ags and some of the most frequently occurring terminal oligosaccharides, as well as core motifs of mammalian N- and O-linked glycoproteins and glycolipids, tumor-associated carbohydrate Ags and polysaccharides from pathogenic bacteria. Synthetic glycan structures (> 95% purity) are structurally the same as natural ones. Their structures, NMR data of polysaccharides and related references are available at http://csdb.glycoscience.ru/bacterial (Zelinsky Institute of Organic chemistry, Moscow, Russia). The glycans (50 mM solutions) and polysaccharides (10 µg/ml) were printed in 6–12 replicates. A complete list of printed glycans can be found in the supplementary material (see Supplement S2). Two chips from each batch were analyzed using standard Complex Immunoglobulin Preparation (CIP, Immunogem, Russia) at a concentration of 1 mg/ml with biotinylated goat-anti-human Abs (Thermo Fisher Scientific, USA) at a concentration of 10 µg/ml followed by Streptavidin Alexa Fluor 555 Conjugate (Thermo Fisher Scientific, USA) at a concentration of 1 µg/ml. Batches of printed microarrays with intra- and inter-chip correlation more than 0.9 were used.
After printing, microarrays were blocked for 90 min at 25℃ with blocking buffer (100 mM boric acid, 25 mM ethanolamine plus 0.2% (v/v) Tween 20 (Sigma-Aldrich, St. Louis, MO, USA), pH 8.5), which was then washed out with milli-Q grade water and dried by the air. Blocked microarrays were stored at –20℃ for further analysis.
Before serum analysis, microarrays were incubated in an incubation chamber for 15 min at 25℃ with PBS plus 0.1% (v/v) Tween-20 (buffer 1) (Sigma-Aldrich, St. Louis, MO, USA), and the buffer was then carefully removed from the microchip surface using Whatman® filter paper (Sigma-Aldrich, St. Louis, MO, USA). Mouse sera were diluted (1:30) in PBS plus 1% (w/v) BSA (Sigma-Aldrich, St. Louis, MO, USA) and 0.1% (v/v) Tween-20 (buffer 2). Diluted serum was spread over the slide surface and incubated with agitation (32–36 rpm) at 37℃ for 1.5 h. After a round of washing steps with buffer 1, buffer 3 (PBS with 0.001% v/v Tween-20) microarrays were then incubated for 1 h at 37℃ (32–36 rpm) with goat anti-mouse IgG + IgM (H + L) conjugated to biotin (Thermo Fisher Scientific, Waltham, MA, USA) at a concentration of 10 µg/ml in PBS plus 1% BSA and 0.1% Tween-20 (buffer 2). The unbound fraction was removed by repeating the same round of washing steps. Microarrays were incubated in darkness at 25℃ for 45 min (32–36 rpm) with streptavidin labeled with Alexa555 dye (Thermo Fisher Scientific, Waltham, MA, USA) at a concentration of 1 µg/ml. After another round of washing (plus using milli-Q grade water at the final stage), the microarrays were dried by airflow. The microarrays were scanned using ScanArrayGx scanner (PerkinElmer, USA) using the excitation wavelength 543 nm. The obtained data were processed using ScanArray Express 4.0 software and the fixed 70 µm-diameter circle method as well as Microsoft Excel software. Six to twelve spot replicates represent each oligosaccharide or polysaccharide on the array, and data are reported as median relative fluorescence units (RFU) of replicates. Median deviation was measured as interquartile range. A signal (the titer of Abs as the result of their affinity and avidity) where the fluorescence intensity exceeded the background value by a factor of five was considered as significant.
The choice of reagents was due to our desire to catch as many diverse Abs as possible, rather than whether they belonged to a particular class. Considering the literature and our own experience of working with small rodents, we did not expect to see a wide repertoire of anti-glycan Abs.25,26 For the same reasons, data on the repertoire of each mouse were not subjected to additional statistical processing and were presented in their original and most detailed form (see Supplementary Table 3).
Results
Abs in the group of sterile mice unexposed to bacteria (group #1)
No anti-glycan Abs was found in a group of mice born and housed under sterile conditions and not exposed to any bacteria, except for one mouse in which serum significant titers of Abs directed to glycans containing Fucα1-2Gal (blood group H motif) were observed, namely Fucα1-2Galβ1-4(Fucα1-3)GlcNAcβ (LeY), Fucα1-2Galβ1-4(Fucα1-3)GlcNAcβ1-3Galβ1-4Glcβ (LeYhexa), GalNAcα1-3(Fucα1-2)Galβ1-3GalNAcα (A type 4), fucosylated polysaccharide from E. coli O127 and also
Abs in groups of mice exposed to a single bacterial strain
Specificity of carbohydrates targeting natural Abs in the group of mice exposed to B. longum, L. reuteri, B. thetaiotaomicron, a mixture of four strains of bacteria (B. longum, L. reuteri, B. thetaiotaomicron and E. coli) and mice put to normal food. The list of glycans with binding signals above 1500 relative fluorescence units (RFU) is presented.
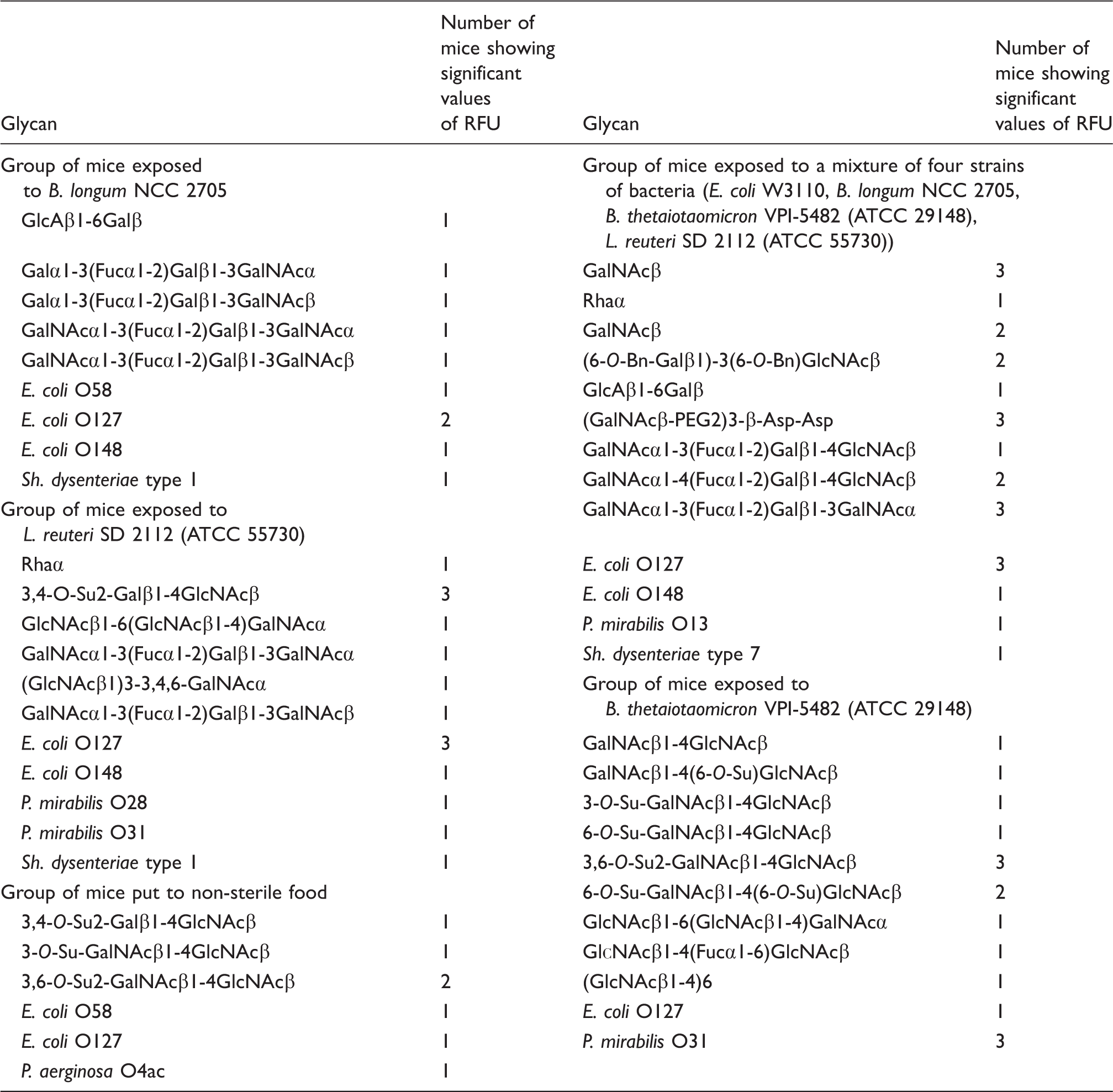
In the group orally inoculated with B. thetaiotaomicron VPI-5482 (ATCC 29148) (group #4), Abs were observed in all animals and were mainly directed to LacdiNAc disaccharide and its O-sulfates. There was a high titer of Abs to P. mirabilis O31 (Table 1).
In the group orally inoculated with L. reuteri SD2112 (ATCC 55730) (group #5), Abs were observed to the blood group ABH Ags,
In the group of animals orally inoculated with a combination of four strains of bacteria (group #6), Abs were detected to the blood group A Ags,
In the group of animals fed with non-sterile food (group #7), Abs were observed to O-sulfated LN, O-sulfated LacdiNAc and to polysaccharides E. coli O58, E. coli O127 and P. aeruginosa O4ac (Table 1). The repertoire of the rise in Abs appears to be poor, similar to previous cases, where only one or a limited number of bacteria were gavaged.
Specificity of carbohydrates targeting natural Abs in the group of mice orally gavaged with a sample of non-sterile mice feces (left columns, ligands positive for at minimum two of four mice are shown; total amount of glycans to which Abs were detected is 54, see Supplementary Table 1) and non-sterile, conventional mice (right columns, only ligands positive for at minimum three of four mice are shown; total amount of glycans to which Abs were detected is 93, see Supplementary Table 1). The list of glycans with binding signals above 1500 relative fluorescence units (RFU) is presented.

In the group of non-sterile mice without any additional challenging (group #9), anti-glycan Abs directed to 67 oligosaccharides and 26 polysaccharides were detected, that is, the number of primed Abs was not significantly higher than in the previous group, although the repertoires were dramatically different (see Supplement 3 and 4). Table 2 demonstrates glycans to which Abs were detected in at least three animals.
Discussion
Glycans that bound Abs in the sera of non-sterile mice as well as mice orally gavaged with feces of non-sterile, conventional mice.

The result of exposure to a combination of four bacteria was not the “sum” of four single-bacteria repertoires (groups #2–6). Notably, specificity of the Abs does not correspond to the chemical structure of Ags in the composition of the bacteria; indeed, Abs directed to ABH blood group Ags are observed, although there is no bacterial blood group related glycoproteins or glycolipids, and their polysaccharides do not contain closely or even remotely related structures.
25
And vice versa: LOS of E. coli W3110, used for the oral inoculation, has the (complete) structure GlcNAcβ1-7Hep1-6Glcα1-2Glcα1-3(Galα1-6)Glcβ-inner core (in which Hep stands for
The priming of Abs formation in animals having standard nutrition (group #7) leads to the appearance of a poor repertoire (about 10 glycans) of Abs; furthermore, these are not in the top-rank Abs found in non-sterile animals (group #9) (Table 1). This observation shows the insufficiency of that set of antigenic stimuli that is present in the nutrition components as such, as well as those belonging to the killed (autoclaved) bacteria – if they were present in the food. Unexpectedly, no identical Ab repertoires were observed for animals orally inoculated with identical bacteria in the same conditions (groups #2–6); this is true for all studied groups, including mice gavaged with feces (group #8), that is, when we observed the highest number of NAbs. This is an astonishing phenomenon. Apparently, apart from the dictates of both identical genetics and the same conditions, the repertoire of the Abs partially has a stochastic component. Notably, this is consistent with previous data demonstrating non-identical NAb repertoires in α-galactosyltransferase-knock-out BALB/c mice housed together,27,28 but is contradictory to the publication in which three Ags were reported to ostensibly have primed their cognate NAbs. 29
However, the immature repertoire can be different in certain animals (individuals), and here the principle ‘first come first served’ as regards to the encounter of B1 cells with bacteria works. The lack of compliance between specificity of challenged Abs to the structure of administered Ags is easily explained in terms of the key role of bacteria in the formation of B1 cell immunity. Antigen mimicry is well known for B2 cell immunity,30–32 i.e. for the production of specific high affinity Abs. B1 cells generally produce a polyreactive low affinity Abs, therefore the phenomenon of mimicry is more “easily allowed” and thus expected to be more common. In particular, human NAbs against blood group A and B antigen bind to bacterial polysaccharides having no any similarity to these antigens, a large number of other examples can be found in the literature. 33
Why are B1 cells primed with bacterial but not primed with self Ags at an early stage of development? The role of TLRs (of B1 lymphocytes) in establishment of natural Abs is well documented; 34 we believe that the trigger function of bacteria in the genesis of NAbs is stipulated by co-localization of Agic determinant with TLR ligands – this promotes the formation of an immuno-synapse 35 followed by activation of the B1 cell. Before birth, the organism's ability to prime B1 cells with bacteria is very limited, so it has virtually no NAbs (or there are none at all, as is the case with the model of sterile animals). Immediately after birth, active occupation of intestine by microorganisms favors the formation of the NAbs repertoire, including Abs to bacteria, self-proteins, allo- and xeno-Ags, tumor-associated epitopes, anionic polymers (DNA, GAGs) and other Ags. In other words, we believe that the total repertoire of NAbs is provided exclusively by microorganisms from a normal microbiota. Although it is not known how many orally administrated antigens reach the mouse digestive tract and pass the mucin/endothelial cells barriers to reach immunocompetent cells, there are assumptions according to which we could suggest that M cells (in Payer's patches) evaluate the intestinal lumen environment and transport bacteria through the epithelial barrier to sub-mucosal macrophages and dendritic cells, which then act as antigen-presenting cells. This process, called ‘bacterial translocation', is well described in sterile animals, as well as prematurely born mice and children, characterized by underdevelopment or imperfection of the mucin layer.35–41
In our work, we can assume (but not affirm) that all or a major part of the anti-glycan Abs we identified are related to NAbs, because of two main reasons that we consider most important. First, Abs emerge as a response to oral contact with a source of a potential Ag in a relatively short time, which is unlikely with an adaptive immune response. Second, we see a structural discrepancy between the specificity of the observed Abs and Ags, whereas in the case of adaptive Abs this structural similarity underlies the specific immune response.
The following conclusions can be drawn.
Sterile animals, reliably deprived of the possibility of contact with microorganisms, have little or no anti-glycan Abs. Contact with a small number of bacterial strains belonging to the normal microflora stimulates (primes) in mice production of anti-glycan Abs; their level and variety are dramatically inferior to Abs of mice living under standard non-sterile conditions. An almost comprehensive repertoire of anti-glycan NAbs is formed as a result of single gavage of digested food of non-sterile animals. The observed priming with bacteria leads to the appearance of Abs directed not only to bacterial polysaccharides, but also to typical glycans of mammal glycoproteins/glycolipids having no distinct structural similarity with bacterial polysaccharides. Specifying the hypothesis of Springer et al.
12
(i.e. the appearance of NAbs as a result of the structural similarity of bacterial polysaccharides with mammalian glycans), we explain this phenomenon not by the direct structural similarity of ABH oligosaccharides and bacterial polysaccharides, but by molecular mimicry when the epitope is a spatial glycotope, a protein epitope or a molecular pattern.
Supplemental Material
Supplementary table1 -Supplemental material for Repertoire of Abs primed by bacteria in gnotobiotic mice
Supplementary tables
Supplemental Material
Supplementary table2 -Supplemental material for Repertoire of Abs primed by bacteria in gnotobiotic mice
Supplementary tables
Supplemental Material
Supplementary table3 -Supplemental material for Repertoire of Abs primed by bacteria in gnotobiotic mice
Supplementary tables
Supplemental Material
Supplementary table4 -Supplemental material for Repertoire of Abs primed by bacteria in gnotobiotic mice
Supplementary tables
Footnotes
Acknowledgments
The work of N Khasbiullina, N Shilova and N Bovin was supported by grant #14-14-00131 of the Russian Science Foundation. The work of Yu Knirel on isolation and characterization of bacterial polysaccharides was supported by grant #14-14-01042 of the Russian Science Foundation.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.