
Abstract
It is generally regarded that Sirtuin 1 (SIRT1), a longevity factor in mammals, acts as a negative regulator of inflammation. However, recent studies also found that SIRT1 might be a detrimental factor under certain inflammatory circumstance. In this study, the potential pathophysiological roles and the underlying mechanisms of SIRT1 in a mouse model with endotoxemia-associated acute lung injury were investigated. The results indicated that treatment with the selective SIRT1 inhibitor EX-527 suppressed LPS-induced elevation of TNF-α and IL-6 in plasma. Treatment with EX-527 attenuated LPS-induced histological abnormalities in lung tissue, which was accompanied with decreased myeloperoxidase level and suppressed induction of tissue factor and plasminogen activator inhibitor-1. Treatment with EX-527 also suppressed LPS-induced phosphorylation of eukaryotic translation initiation factor-binding protein 1 (4E-BP1). Co-administration of a mammalian target of rapamycin (mTOR) activator 3-benzyl-5-[(2-nitrophenoxy) methyl]-dihydrofuran-2 (3H)-one (3BDO) abolished the inhibitory effects of EX-527 on 4E-BP1 phosphorylation. Meanwhile, the inhibitory effects of EX-527 on IL-6 induction and the beneficial effects of EX-527 on lung injury were partially reversed by 3BDO. This study suggests that selective inhibition of SIRT1 by EX-527 might alleviate endotoxemia-associated acute lung injury partially via suppression of mTOR, which implies that SIRT1 selective inhibitors might have potential value for the pharmacological intervention of inflammatory lung injury.
Introduction
Acute lung injury (ALI), triggered by systemic or pulmonary infection, trauma and other harmful factors, is a common cause of acute respiratory failure with a high morbidity and mortality rate. 1 ALI is characterized as extensive destruction of the alveolar-capillary membrane and severe hypoxia. 2 In principle, the excessive activation of inflammatory cells and the overproduction of pro-inflammatory mediators in ALI play pivotal roles in the development of respiratory membrane damage. 3 In addition, the uncontrolled inflammation abnormally activates the coagulation cascade, which further impairs pulmonary gas exchange and has a synergistic association with higher mortality in patients with ALI. 4
Sirtuin 1 (SIRT1) is an NAD+-dependent class III histone deacetylase (HDAC), which attracts extensive research interests because it is suggested to function as a longevity factor in mammals. 5 In addition to its crucial roles in the control of senescence, recent studies have found that SIRT1 exhibits pronounced anti-inflammatory properties via deacetylation of several inflammation-associated transcription factors.6,7 Therefore, SIRT1 is regarded as a novel target for the negative regulation of inflammation and several SIRT1 activators have shown beneficial effects in animal models with inflammatory disorders.8–10
However, several studies have challenged the beneficial roles of SIRT1 in inflammatory injury. In a murine model of ovalbumin-induced bronchial asthma, inhibition of SIRT1 resulted in reduced inflammatory cells infiltration and suppressed induction of pro-inflammatory cytokines.
11
In mice with LPS/
To explore the pathophysiological significance of SIRT1 in the progression of ALI induced by systemic LPS exposure, a selective SIRT1 inhibitor EX-527 was administered in mice with endotoxemia. EX-527 is widely used as a SIRT1 inhibitor in experimental studies both in vitro and in vivo with high potency and significant isoform selectivity.14,15 Crystal structure analysis suggested that EX-527 interacts with SIRT1 at the nicotinamide site, which provides the molecular basis of SIRT1 inhibition by EX-527.16,17 In the present study, the degree of lung injury, inflammatory response and coagulation activation were determined to evaluate the potential effects of EX-527 on endotoxemia-associated ALI.
Materials and methods
Animals
Male Balb/C mice, 6–8 wk of age (weighing 18–22 g) were obtained from the Experimental Animal Center of Chongqing Medical University. Mice were fed with a standard laboratory diet and water ad libitum. All animals were maintained in steel cages in a room with a controlled temperature of 20–25℃ and a 12-h light/12-h dark cycle. All animal protocols were approved by the Institutional Animal Care and Use Committee of Chongqing Medical University.
Reagents
LPS (Escherichia coli 055:B5), the SIRT1 inhibitor EX-527 and the mTOR activator 3-benzyl-5-((2-nitrophenoxy) methyl)-dihydrofuran-2 (3H)-one (3BDO) were purchased from Sigma-Aldrich (St. Louis, MO, USA). Myeloperoxidase (MPO) detection kit was produced by Nanjing Jiancheng Bioengineering Institute (Nanjing, China). ELISA kits for detecting mouse TNF-α and IL-6 were purchased from NeoBioscience Technology Company (Shenzhen, China). The rabbit anti-mouse β-actin, tissue factor (TF), plasminogen activator inhibitor-1 (PAI-1), eukaryotic translation initiation factor 4E (eIF4E)-binding protein 1 (4E-BP1), phosphorylated 4E-BP1 (p-4E-BP1) Abs were purchased from Abcam (Cambridge, UK). The BCA protein assay kit, HRP-conjugated goat anti-rabbit Ab and enhanced chemiluminescence (ECL) reagents were purchased from Pierce Biotechnology (Rockford, IL, USA).
Experimental protocol
After 1 wk of adaptation, endotoxemia in mice was induced by i.p. injection of LPS (20 mg/kg, dissolved in 0.9% physiological saline). To investigate the potential roles of SIRT1, the animals were randomly divided into the following groups (n = 8, each group): (A) the control (CON) group—mice received vehicle only; (B) the EX-527 group—mice injected i.p. with SIRT1 inhibitor EX-527 (10 mg/kg, dissolved in DMSO); (C) the LPS group—mice with LPS-induced endotoxemia; (D) the LPS + EX-527 group—mice received EX-527 (10 mg/kg) administration 0.5 h prior to LPS challenge.
To explore the underlying mechanisms, the animals were randomly divided into the following groups (n = 8, each group): (A) the LPS group—mice with LPS-induced endotoxemia; (B) the LPS + EX-527 group—mice received the SIRT1 inhibitor EX-527 (10 mg/kg) administration 0.5 h prior to LPS challenge; (C) the LPS + EX-527 + 3BDO group—mice received the mTORC1 activator 3BDO (100 mg/kg, dissolved in DMSO) 0.5 h prior to EX-527 administration, and LPS was injected at 0.5 h later.
Next, the animals were returned to their cages and provided with food and water ad libitum. The mice were anesthetized and sacrificed at 18 h after LPS challenge. The plasma samples were harvested for determining TNF-α and IL-6. The left lobe of the lung was fixed in formalin for morphological examination and the remaining lung tissues were stored at –80℃ for further experiments.
Histological analysis
The lobe of left lung was fixed in formalin and embedded in paraffin. The sections were stained with hematoxylin and eosin for histopathological evaluation under a light microscope (Olympus, Tokyo, Japan).
Measurement of TNF-α and IL-6
The levels of TNF-α and IL-6 in plasma were assessed according to the manufacturer's instructions by using the ELISA kits. The amount of TNF-α and IL-6 were read off the standard curves.
Determination of MPO activity
The lung tissue samples were thawed and homogenized in 1 ml 50 mmol/l potassium PBS, pH 6.0, containing 0.5% hexadecyltrimethylammonium bromide. The samples were then centrifuged at 10000 g for 15 min at 4℃. The enzyme activity was tested by measuring absorbance changes in the redox reaction of H2O2 by spectrophotometry at 460 nm. The value was normalized by the total protein concentration of each sample and the results expressed as MPO units/g of protein.
Western blot analysis
The tissue samples were homogenized and total proteins were prepared with the protein extract kit according to the manufacturer's instructions (Nanjing Jiancheng Bioengineering Institute, China). The total protein concentration was determined using the BCA protein assay kit (Pierce Biotechnology). Protein extracts were fractionated by SDS-PAGE (10% polyacrylamide) and then transferred to nitrocellulose membrane. The membrane was blocked with 5% (w/v) nonfat milk (in Tris-buffered saline, containing 0.05% Tween-20), and then the membrane was incubated with primary Ab overnight at 4℃, followed by incubation with secondary Ab. Ab binding was visualized with an ECL chemiluminescence system and short exposure of the membrane to X-ray films.
Statistical analysis
Data are expressed as mean ± SD, and differences between means were analyzed by one-way ANOVA with the Turkey's post-hoc test. Results were considered statistically significant when P < 0.05.
Results
SIRT1 inhibitor alleviated LPS-induced systemic inflammation
The effects of the SIRT1 inhibitor EX-527 on LPS-induced systemic inflammation were evaluated by determination of the levels of TNF-α and IL-6 in plasma. As shown in Figure 1, the LPS-challenged mice exhibited a significant increase in TNF-α and IL-6, which was significantly decreased after EX-527 administration. These findings indicated that EX-527 suppressed LPS-induced systemic inflammation.
EX-527 suppressed LPS-induced systemic inflammation. Mice were treated with vehicle or EX-527 (10 mg/kg) in the absence or presence of LPS (20 mg/kg) exposure. The plasma samples were harvested at 18 h after LPS exposure. The levels of (a) TNF-α and (b) IL-6 in plasma were determined by ELISA. Data expressed as mean ± SD, n = 8. *P < 0.05, **P < 0.01.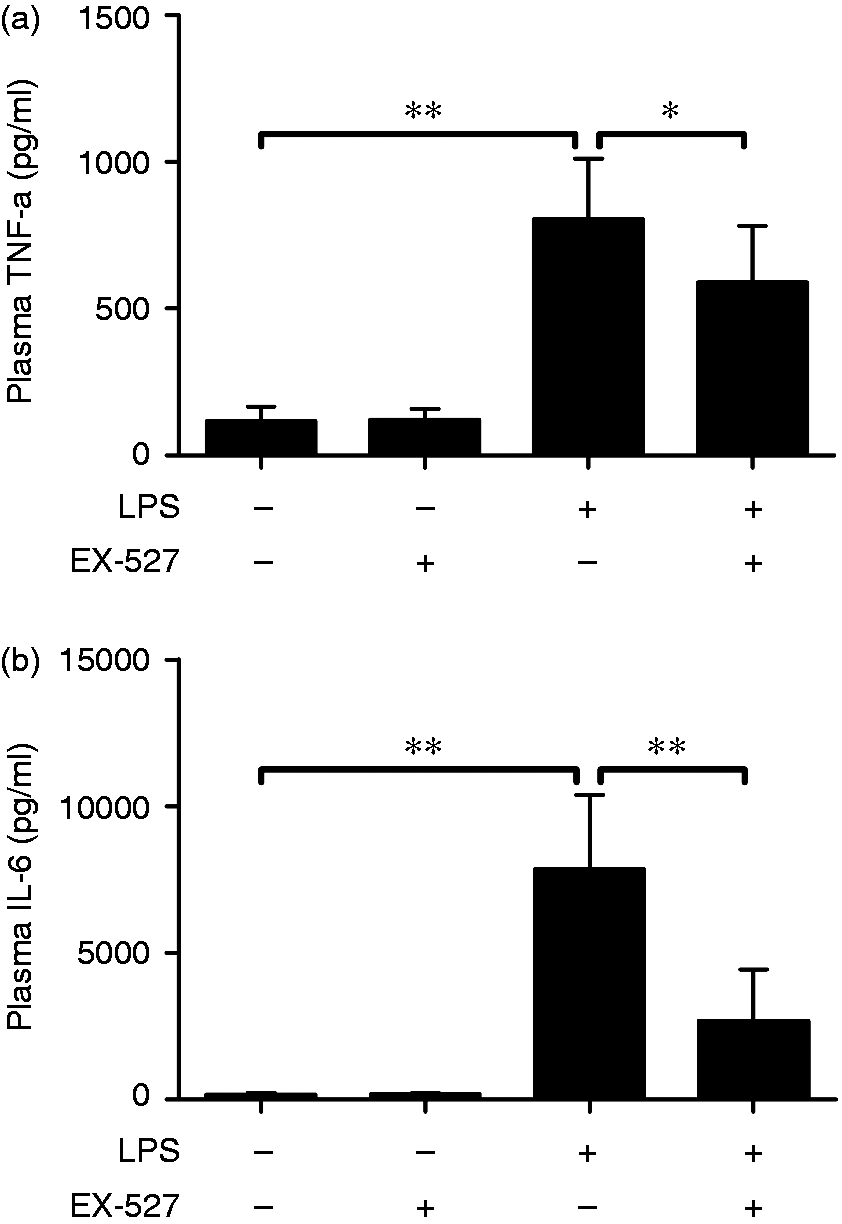
SIRT1 inhibitor attenuated LPS-induced lung injury
The lung is the most sensitive organ be severely damaged under systemic inflammatory circumstance.
18
In the morphological examinations, LPS-induced histopathological abnormalities, including leukocytes infiltration, hemorrhage, bronchial wall thickening and alveolar edema, were significantly alleviated by EX-527 (Figure 2). These results indicated that EX-527 treatment alleviated endotoxemia-associated ALI.
EX-527 alleviated LPS-induced histological abnormalities in lung. Mice were treated with vehicle or EX-527 (10 mg/kg) in the absence or presence of LPS (20 mg/kg) exposure. The lung samples were harvested at 18 h after LPS exposure. The histopathological alterations of lung tissue were evaluated with hematoxylin and eosin staining. The representative lung sections of each group are shown (original magnification: 100× and 400×).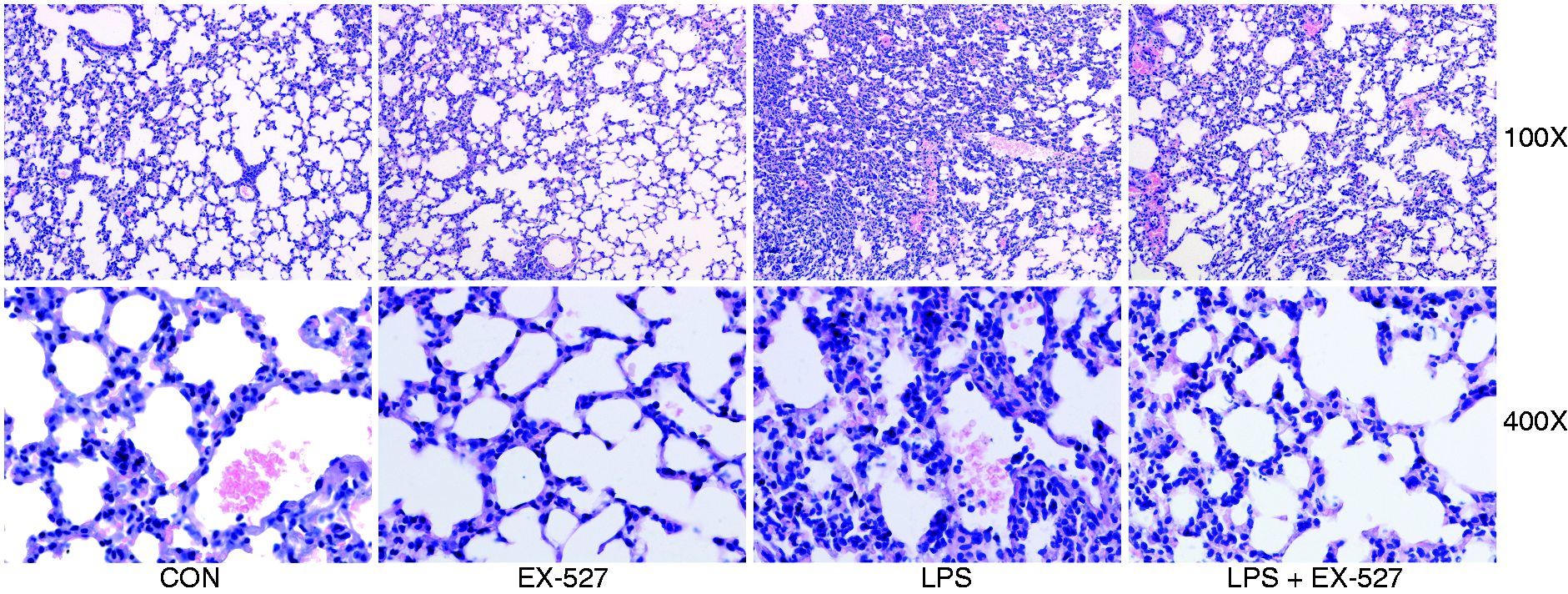
The SIRT1 inhibitor resulted in suppressed intrapulmonary inflammation and coagulation
MPO is a constituent enzyme found principally in neutrophil, which is widely regarded as a molecular marker of neutrophil infiltration.
19
The marked elevation of MPO was detected in the LPS-exposed group, but the elevated level of MPO was suppressed by EX-527 (Figure 3). These data indicated that LPS-induced intrapulmonary inflammation was attenuated by EX-527.
EX-527 suppressed LPS-induced elevation of MPO in lung. Mice were treated with vehicle or EX-527 (10 mg/kg) in the absence or presence of LPS (20 mg/kg) exposure. The lung samples were harvested at 18 h after LPS exposure and the activities of MPO in lung tissue were determined. Data expressed as mean ± SD, n = 8. ** P < 0.01.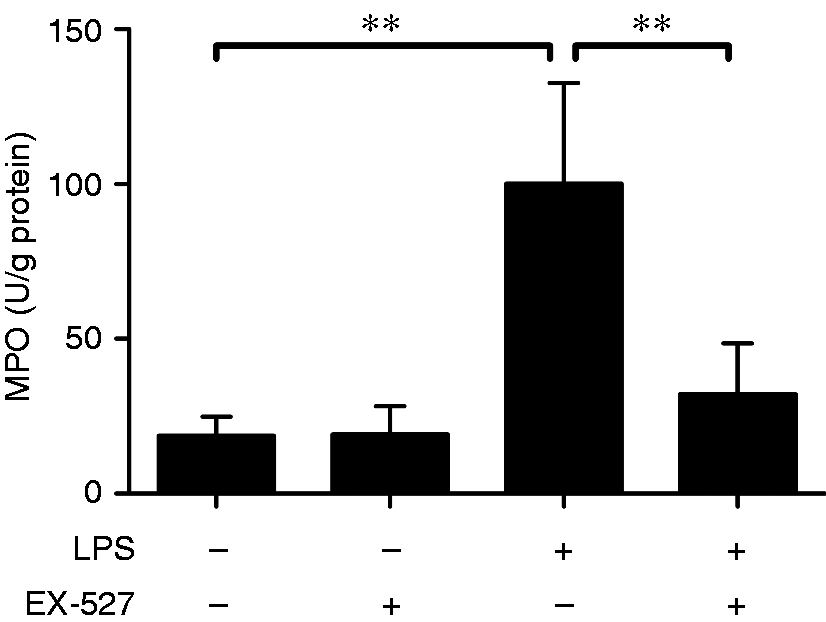
The excessive inflammatory response is usually associated with the up-regulation of TF and PAI-1, which might result in abnormal activation of coagulation and the exacerbated tissue injury.
20
The present study found that the levels of TF and PAI-1 significantly increased in the LPS-treated group, but EX-527 treatment could suppress these alterations (Figure 4). These data indicated that LPS-induced intrapulmonary coagulation was also suppressed by EX-527.
EX-527 suppressed LPS-induced expression of TF and PAI-1. Mice were treated with vehicle or EX-527 (10 mg/kg) in the absence or presence of LPS (20 mg/kg) exposure. The lung samples were harvested at 18 h after LPS exposure. (a) The level of TF and PAI-1 were determined by immunoblotting. The target proteins are indicated by arrows in the blot. The bands of (b) TF and (c) PAI-1 were semi-quantified by gray scale. Data expressed as the mean ± SD, n = 4. *P < 0.05, **P < 0.01.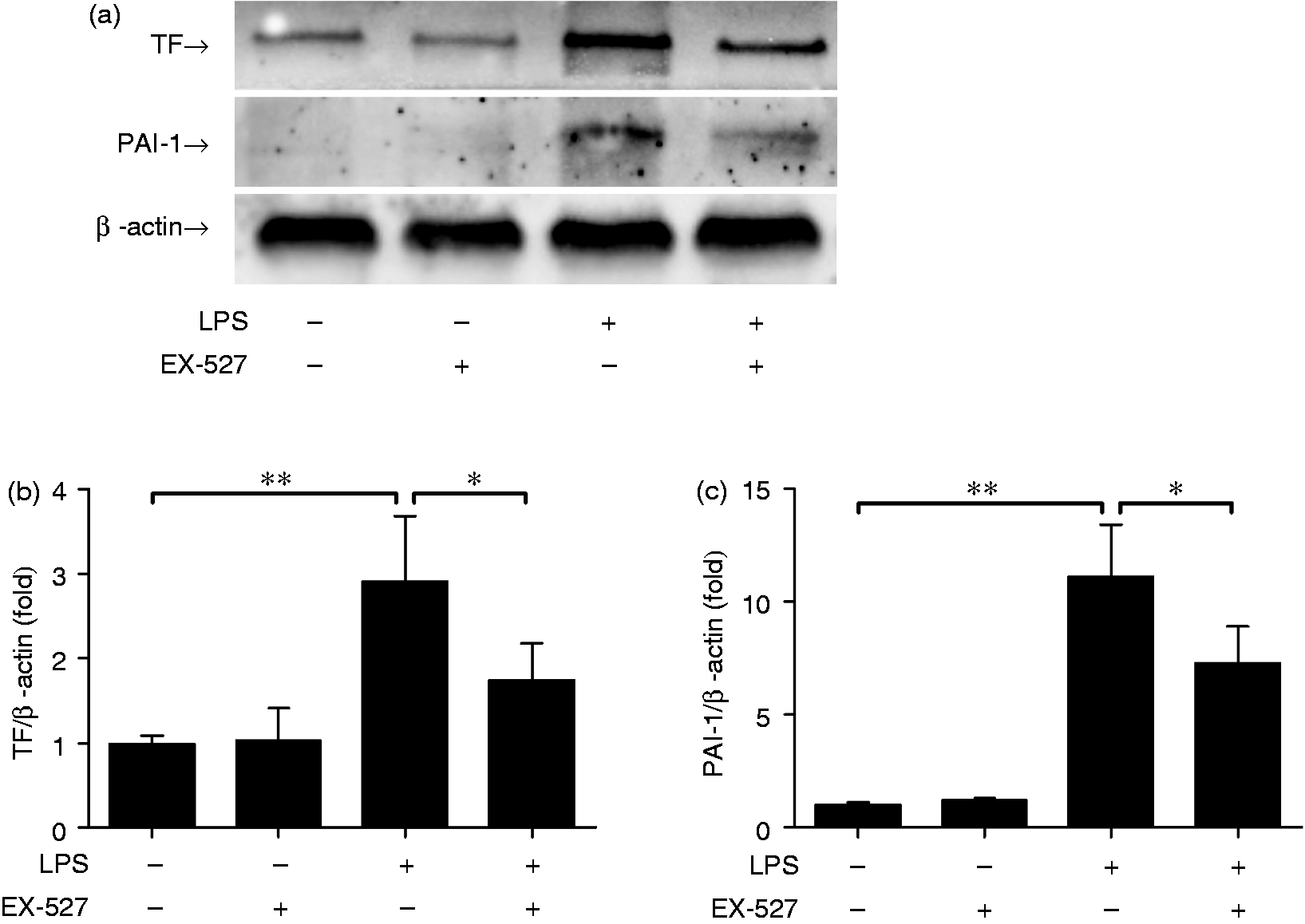
The beneficial effects of EX-527 were associated with suppressed mTOR activation
Previous reports have demonstrated that SIRT1 was involved in the regulation of mTOR.21,22 The present study analyzed the phosphorylation level of 4E-BP1, a representative downstream target of mTOR.
23
As shown in Figure 5, treatment with the SIRT1 inhibitor EX-527 significantly suppressed LPS-induced elevation of 4E-BP1 phosphorylation.
EX-527 suppressed LPS-induced phosphorylation of 4E-BP1. Mice were treated with vehicle or EX-527 (10 mg/kg) in the absence or presence of LPS (20 mg/kg) exposure. The lung samples were harvested at 18 h after LPS exposure. (a) The level of p-4E-BP1 and total 4E-BP1 were determined using immunoblot assay. The target proteins were indicated by arrows in the blot. (b) The bands of p-4E-BP1 and 4E-BP1 were semi-quantified by gray scale. Data expressed as the mean ± SD, n = 4, *P < 0.05, **P < 0.01.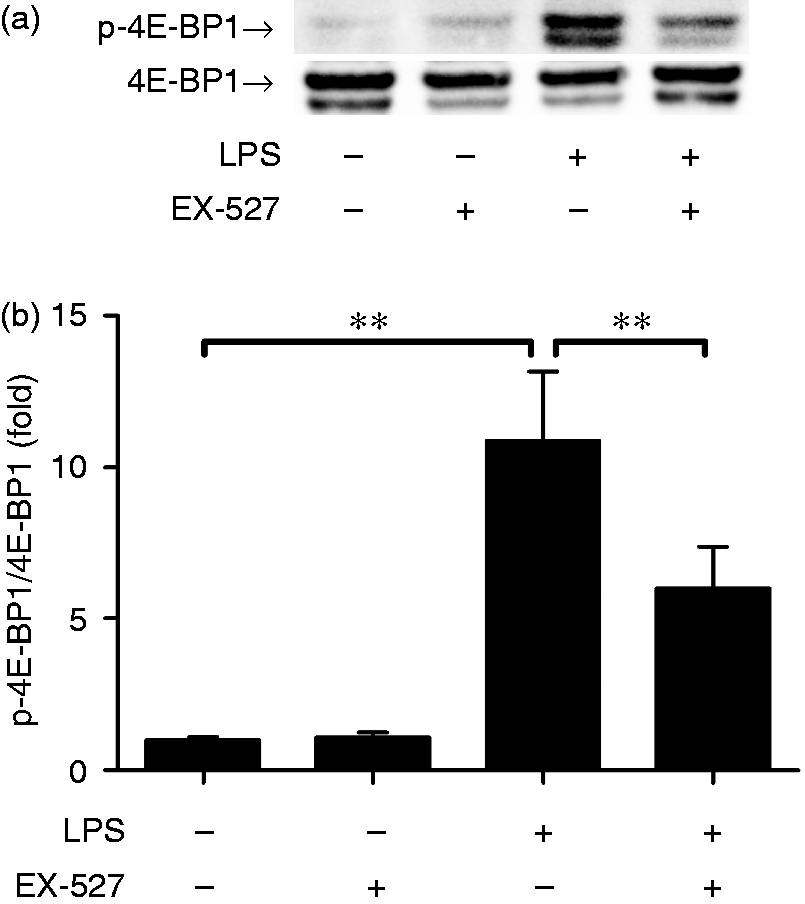
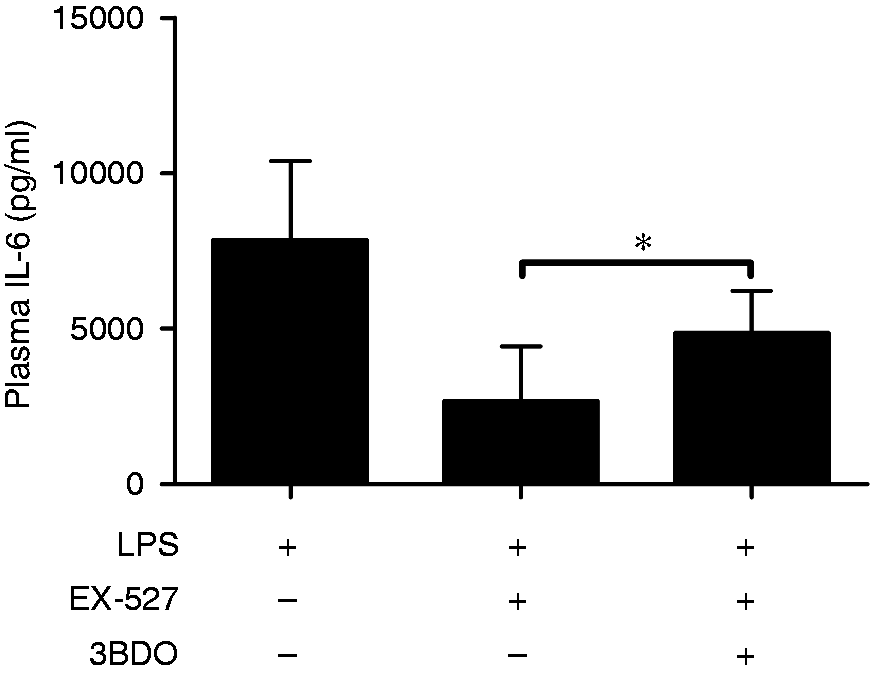
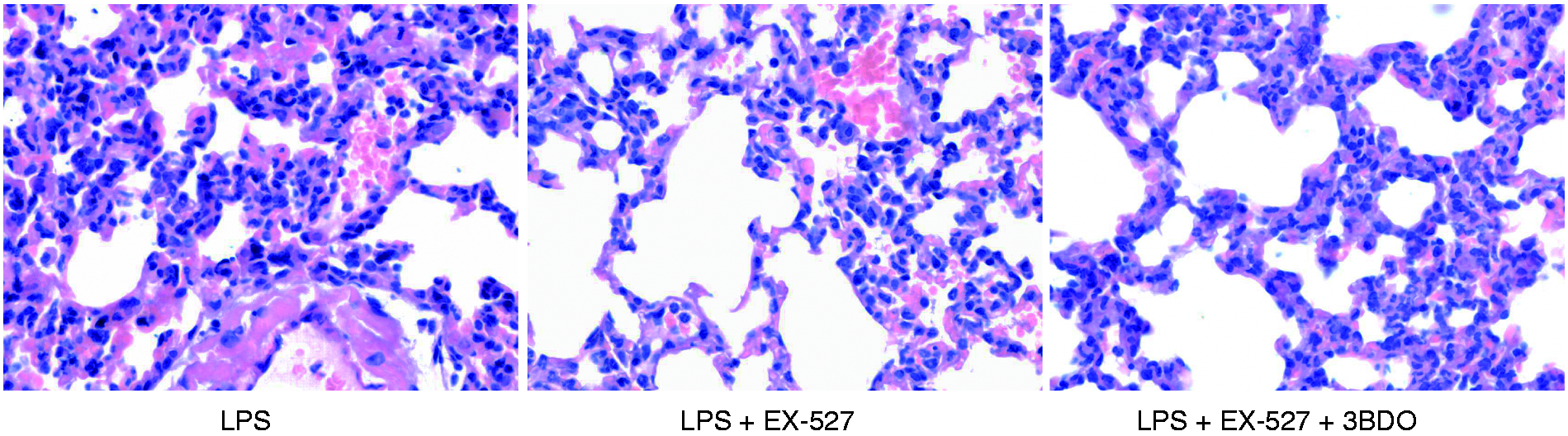
To investigate the potential roles of mTOR in the beneficial effects of EX-527, 3BDO, a mTOR activator,24,25 was co-administrated with EX-527. The results indicated that co-treatment with 3BDO abolished the inhibitory effects of EX-527 on LPS-induced 4E-BP1 phosphorylation (Figure 6). In addition, the suppressive effects of EX-527 on IL-6 induction and lung injury were also partially reversed by 3BDO (Figures 7 and 8). These data suggest that the anti-inflammatory benefits of EX-527 might be associated with suppressed mTOR activation.
3BDO abolished the inhibitory effects of EX-527 on 4E-BP1 phosphorylation. LPS (20 mg/kg)-exposed mice were treated with EX-527 (10 mg/kg) in the absence or presence of 3BDO (100 mg/kg). The lung samples were harvested at 18 h after LPS exposure. (a) The level of p-4E-BP1 and total 4E-BP1 were determined using immunoblot assay. The target proteins were indicated by arrows in the blot. (b) The bands of p-4E-BP1 and 4E-BP1 were semi-quantified by gray scale. Data expressed as the mean ± SD, n = 4. **P < 0.01. mTOR activator 3BDO partially reversed the anti-inflammatory effects of EX-527. LPS (20 mg/kg)-exposed mice were treated with EX-527 (10 mg/kg) in the absence or presence of 3BDO (100 mg/kg). The lung samples were harvested at 18 h after LPS exposure. The plasma samples were harvested at 18 h after LPS exposure and the levels of IL-6 in plasma were determined by ELISA. Data expressed as mean ± SD, n = 8, *P < 0.05. 3BDO partially reversed the beneficial effects of EX-527 on lung injury. Mice were treated with EX-527 (10 mg/kg) or 3BDO (100 mg/kg) + EX-527 in the presence of LPS exposure. The lung samples were harvested at 18 h after LPS exposure. The histopathological alterations of lung tissue were evaluated with hematoxyline eosin staining. The representative lung sections of each group are shown (original magnification: 400×).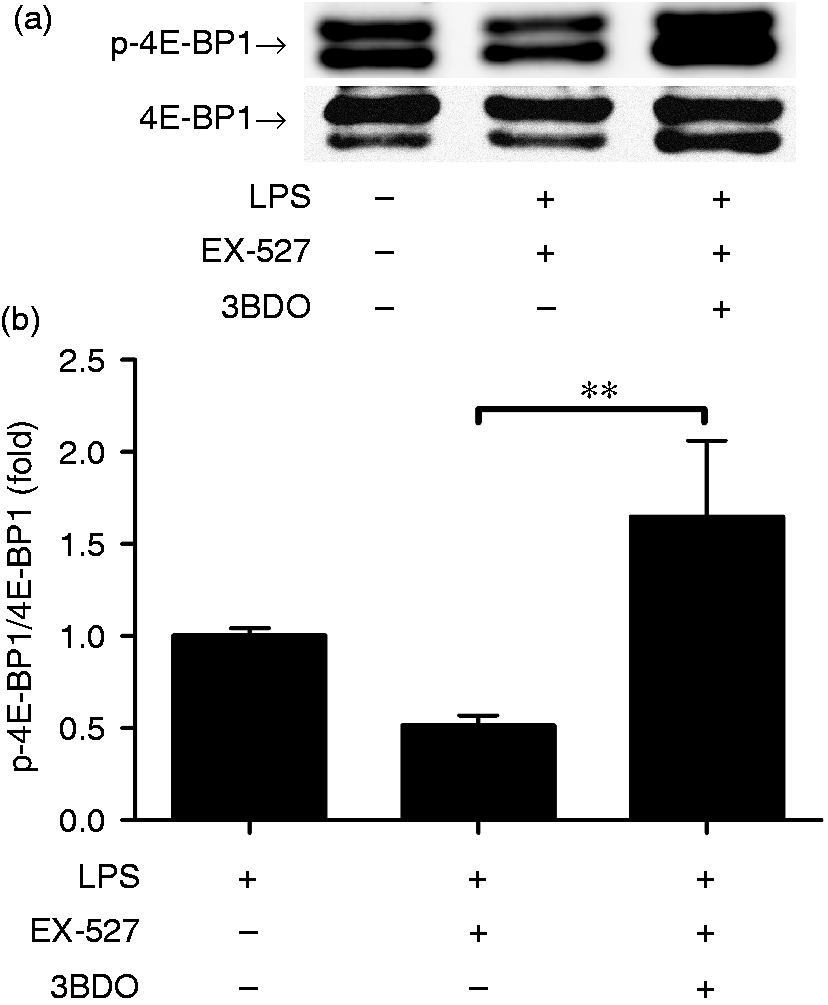
Discussion
SIRT1 is one of the interesting HDACs that attracts wide interests in a series of research fields, including inflammatory disorders. 5 It is generally thought that SIRT1 might suppress inflammation via its deacetylation activities. 7 However, just like the dual effects associated with other post-translational modifications (PTMs), such as phosphorylation, polyubiquitination and SUMOylation,26,27 SIRT1-mediated deacetylation might not always be associated with negative effects on inflammatory response.12,28 In the present study we found that treatment with the selective SIRT1 inhibitor EX-527 alleviated LPS-induced systemic inflammation and ALI. Therefore, the SIRT1 inhibitor EX-527 might have beneficial effects on endotoxemia-associated ALI.
Inflammation is a serial of complex molecular responses orchestrated by coordinated expression of inflammation-associated genes. 29 PTMs on histones play crucial roles in the regulation of the transcription of these genes. 30 Lysine acetylation is a well-known PTM that is reversely catalyzed by the histone acetyltransferases (HATs) and HDACs. 30 Acetylation of histones could neutralize the positive charge of this amino acid, which is usually associated with open chromatin architecture and transcriptional activation. 31
However, decreased acetylation by inhibition of HATs did not suppress pro-inflammatory gene expression.32,33 On the contrary, up-regulation of acetylation by inhibition of HDACs commonly resulted in suppressed inflammatory response and alleviated inflammatory injury.34,35 In the present study, we found that treatment with the selective inhibitor of SIRT1, a class III HDAC, significantly attenuated LPS-induced inflammatory injury. Consistently, the anti-inflammatory properties of SIRT1 inhibitors were confirmed in experimental animals with hepatitis, sepsis and asthma.11–13 In addition, inhibition or silence of SIRT2 also exhibited anti-inflammatory effects.28,36 Therefore, treatment with pan-HDAC inhibitors or selective HDAC inhibitors might be associated with suppressed inflammation, but the underlying mechanisms remains unclear.
In addition to suppressed inflammatory response, treatment with the SIRT1 inhibitor EX-527 also suppressed LPS-induced up-regulation of TF and PAI-1, crucial factors involved in inflammation-associated pathological coagulation. 4 TF is the initiation factor of the extrinsic coagulation pathway. 37 TF is expressed by the cells surrounding blood vessels, TF might initiate the coagulation response when the blood vessel is damaged and TF is exposed to the coagulation cascade. 37 Inflammatory stimuli such as TNF-α could induce the expression of TF in blood-contacting cells, such as endothelial cells and monocytes, and abnormally activates the coagulation response. 38 In addition, PAI-1, a serine protease inhibitor that functions as the principal inhibitor of fibrinolysis, is also induced by inflammatory stimuli. 39 Enhanced coagulation and suppressed fibrinolysis are usually associated microcirculatory disturbance, which is another important pathological mechanism underlying the development of ALI. 20 Thus, the beneficial effects of EX-527 might also attribute to the suppressed induction of TF and PAI-1.
Histones were the first group of proteins identified as targets for acetylation modification, but a global analysis has recently mapped >1750 proteins that can be acetylated, 31 suggesting that acetylation might have profound regulatory effects under various physiological or pathophysiological conditions. It was reported that acetylation of p65 was required for the full activity of NF-кB, which was associated with increased expression of pro-inflammatory genes. 7 On the contrary, acetylation of mitogen-activated protein kinase phosphatase-1 (MKP-1) enhances the interaction of MKP-1 with p38, which is associated with decreased phosphorylation of p38 and the suppressed inflammatory response. 40 Therefore, acetylation modifications might have both positive and negative effects on inflammation-related proteins. It is possible that SIRT1 might have both beneficial or detrimental effects under certain inflammatory conditions.
Several inflammation-related proteins have been reported to the inflammation-regulatory activities of SIRT1, such as NF-кB, activator protein 1, high mobility group box-1 protein, and signal transducer and activator of transcription 3.7,41,42 It seems that deacetylation of these target proteins by SIRT1 might be associated with suppressed inflammatory response.7,41,42 In addition, recent studies indicated that acetylation modification plays crucial roles in the regulation of mTOR and some regulatory functions of SIRT1 might be associated with mTOR.21,22
mTOR is a nutrient sensor that acts as a crucial regulator in cell proliferation, cell survival, protein synthesis, autophagy and transcription.
43
mTOR is a serine/threonine protein kinase that belongs to the phosphatidylinositol 3-kinase-related kinase family.
43
4E-BP1 is a representative downstream target of mTOR, which inhibits cap-dependent translation by binding to eIF4E.
44
Phosphorylation of 4E-BP1 by mTOR resulting in its dissociation from eIF4E, and eIF4E then interacts with the eukaryotic translation initiation factor 4G (eIF4G),
44
which is crucial for the assembly of the translation initiation complex (Figure 9). Several studies have found that inflammatory stimuli could activate mTOR.45,46 In addition, inhibition or deletion of mTOR attenuated LPS-induced ALI.47,48 Therefore, mTOR might be a pivotal modulator in the development of ALI.
The schematic diagram of the mechanisms underlying the beneficial effects of EX-527 on LPS-induced acute lung injury. The exposure to LPS activates the expression of pro-inflammatory cytokines (e.g. TNF-α and IL-6), and coagulation-related factors (e.g. TF and PAI-1), which play crucial roles in excessive activation inflammation and the abnormal activation of coagulation. Transcription and translation are the primary steps of gene expression, mTOR promotes translation via phosphorylation of 4E-BP1. 4E-BP1 inhibits cap-dependent translation by binding to the eIF4E. Phosphorylation of 4E-BP1 by mTOR results in its dissociation from eIF4E, and eIF4E then interacts with the eIF4G, which is crucial for the assembly of the translation initiation complex and the activation of translation. SIRT1 has been suggested to play crucial roles in the activation of mTOR. Treatment with the selective SIRT1 inhibitor suppressed the activation of mTOR and subsequently inhibited the induction of pro-inflammatory cytokines and coagulation-related factors.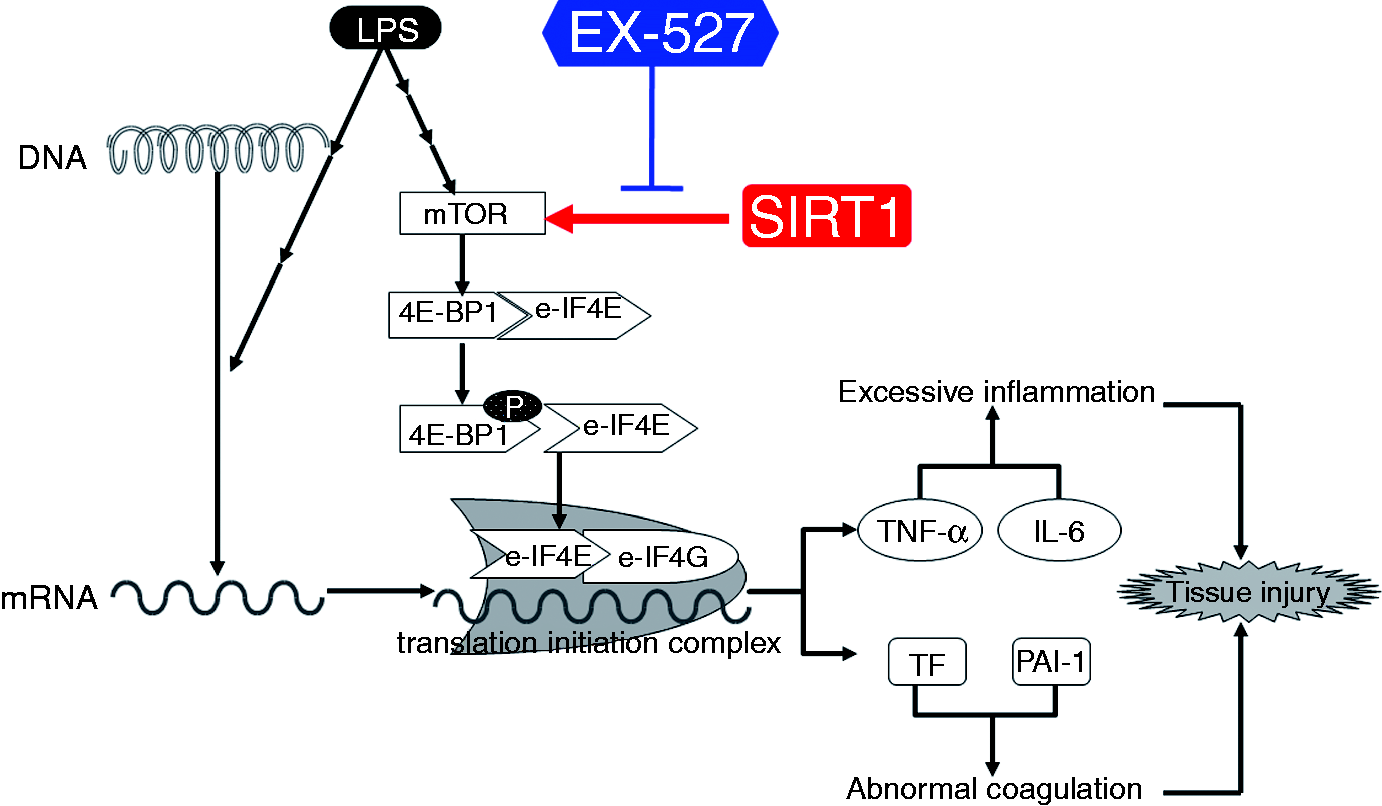
We then questioned whether mTOR was involved in the anti-inflammatory effects of EX-527. Our results indicated that treatment with EX-527 suppressed LPS-induced activation of mTOR, co-administration of mTOR activator reversed the suppression of mTOR and partially abolished the anti-inflammatory effects of EX-527. These data suggest that suppression of mTOR might contribute to the anti-inflammatory activities of EX-527.
Taken together, this study found that selective inhibition of SIRT1 resulted in suppressed inflammation and alleviated lung injury in mice with endotoxemia. More interestingly, the results indicated that the anti-inflammatory effects of the SIRT1 inhibitor might be partially attributed to the suppression of mTOR. Although the molecular mechanisms underlying the pathophysiological significance of SIRT1 in inflammatory disorders still require further intensive investigation, the present study suggested that SIRT1 selective inhibitors might have potential value for the pharmacological intervention of inflammatory disorders.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the grants from the National Nature Science Foundation of China (No. 81370179, 81671953).