
Abstract
Porphyromonas gingivalis produces different LPS isoforms with significant structural variations of their lipid A and O-antigen moieties that can affect its pro-inflammatory and bone-resorbing potential. We show here, for the first time, that P. gingivalis LPS isolated from W83 strain is highly sialylated and possesses significantly reduced inflammatory potential compared with less sialylated ATCC 33277 strain LPS. Nevertheless, the reduction in the endotoxin activity is not mediated by the presence of sialic acid LPS moieties as the sialic acid-free LPS produced by the mutant W83 strain exhibits a similar inflammatory potential to the wild type strain. Furthermore, our findings suggest that the interaction between the sialic acid LPS moieties and the inhibitory CD33 receptor is prevented by endogenously expressed sialic acid on the surface of THP-1 cells that cannot be out-competed by sialic acid containing P. gingivalis LPS. The present study also highlights the importance of endogenous sialic acid as a ‘self-associated molecular pattern’ and CD33 receptors in modulation of innate immune response as human gingival fibroblasts, which do not express CD33 receptors, and desialylated THP-1 cells have both been found to have much higher spontaneous IL-8 production than naïve THP-1 cells.
Introduction
The ability to synthesize, take up, use and catabolize sialic acid is prevalent among commensals and pathogens that colonize the oral cavity and the intestinal, respiratory and urogenital tracts. 1 Periodontal diseases are caused by dental plaque bacteria of the so-called ‘red complex’, which can use different molecular mechanisms and strategies to evade or overcome host defences. 2 Recent findings suggest that the host immune response is initially subverted by these keystone pathogens and is subsequently over-activated by pathobions, leading to polymicrobial synergy and formation of dysbiotic subgingival microflora. 3 One of the mechanisms by which periodontopathogens can manipulate the immune responses of macrophages and monocytes is through their surface glycosylation. 4 It has been shown that bacterial surface-expressed O-linked glycans and especially terminally present sialic acid can help them evade the host innate immune attacks and also modulate T-cell responses during periodontal inflammation. 5 Porphyromonas gingivalis, the keystone periodontal pathogen, utilizes a panel of virulence factors that can cause deregulation of the innate immune and inflammatory responses. P. gingivalis LPS is one of its most potent virulence factors. Whilst LPS recognition by the host is essential for clearing infections of this invading periodontal pathogen, at the same time it triggers a robust inflammatory reaction and is responsible for alveolar bone resorption. The host’s inflammatory response can effectively be modulated by modifications of either the lipid-A core of LPS or its oligo-/polysaccharide domain (O-antigen). 6
Sialic acids are a large family of sugars containing 9 C atoms, which predominantly derive from N-acetylneuraminic acid (NANA). 7 NANA (the most abundant and best-studied sialic acid) is usually a terminal sugar residue on the oligosaccharide chains of eukaryotic cells where it functions in recognition and anti-recognition phenomena. 8 Sialic acids are transferred using α-(2→3) or α-(2→6) linkages to sub-terminal sugars by a family of sialyltransferases and are abundantly expressed on the periphery of mammalian immune cell surface glycans and serum proteins. 9 The mammalian glycome contains numerous sialylated glycans that can be recognized as ligands by special receptors present on the cell surface, called Siglecs (sialic acid-binding immunoglobulin like lectins). It is assumed that this recognition is important for modulating the function of Siglecs as regulators of adhesion, cell signaling and endocytosis. 10 The local concentration of sialic acid on the surface of immune cells is very high; as a consequence, the binding sites of Siglecs are usually masked by low-affinity cis interactions with endogenous glycans on the same cell. 11 However, high-affinity exogenous ligands can out-compete cis ligands and more potently engage an inhibitory Siglec, proving that binding of Siglecs to trans ligands can occur dynamically in the presence of cis ligands. 12
Siglecs are a family of immunoreceptor tyrosine-based inhibition motif (ITIM)-containing receptors that can recognize sialylated glycans and function as regulators of immune cells. CD33-related Siglecs are mainly expressed by cells of the innate immune systems such as monocytes (Siglec-3), macrophages, NK cells, neutrophils, eosinophils, DCs and mast cells. 13 CD33-related Siglecs have also been found expressed on cells of the nervous system (Siglec-4), placental trophoblasts (Siglec-6) and epithelial cells (Siglec-12) but not on fibroblasts.14,15 Ligand engagement of these inhibitory receptors causes phosphorylation of the ITIMs. This allows the recruitment of at least one of the SH2-domain-containing protein tyrosine phosphatases 1 and 2 (SHP1, SHP2) as well as inositol 5’-phosphatase (SHIP). These phosphatases can subsequently dephosphorylate relevant intracellular substrates thus regulating cellular activation by attenuating or terminating tyrosine phosphorylation signal transduction. 16
Sialic acid is an important molecule for a number of bacteria, too, serving as a substrate for capsule synthesis, LPS sialylation and as a potential growth factor. 17 A variety of pathogens have been able to exploit sialic acid by expressing it on their surface, thus evading the immune system, or binding to host cells via sialic acid-recognizing receptors, which mediate cell adhesion, protein–protein interactions and immune response. 18 Some bacteria have the potential to incorporate sialic acid into their LPS as a terminal residue of the carbohydrate part of the molecule. Sialylated LPS glycoforms play a key role in pathogenicity of Haemophilus influenzae and Neisseria meningitidis. 19
There is a growing body of evidence that sialic acid and sialidases play a key role in the life, host mimicry and consequences of periodontal pathogen colonisation. 20 P. gingivalis, as a member of the ‘red complex bacteria’, primarily inhabits the gingival sulci and periodontal pockets. Sialic acid is abundantly present in human serum, which is the major component of the gingival crevicular fluid, while salivary mucins and gingival connective tissue fibronectins also contain sialic acid. P. gingivalis is able to acquire sialic acid from the host, and use it as an important factor that contributes to biofilm formation, capsule biosynthesis and pathogenicity of P. gingivalis. 21 However, the role of sialylation of P. gingivalis LPS in the interaction with the host cells is still unknown. Thus, the aim of this study was to examine the sialic acid content in LPS extracted from two different strains of P. gingivalis (P. gingivalis ATCC 33277 and P. gingivalis W83) and to determine the influence of LPS sialylation on its inflammatory potential. Our results show for the first time that P. gingivalis W83 LPS is highly sialylated but the presence of sialic acid as an LPS carbohydrate moiety does not play a significant role in modulating the inflammatory potential of the LPS of this periodontal pathogen.
Materials and methods
P. gingivalis ATCC 33277 LPS was obtained from InvivoGen (San Diego, CA, USA). P. gingivalis W83 LPS was extracted following the method reported by Darveau and Hancock. 22 Sialic-acid free LPS extracted from P. gingivalis W83 ΔPG0083 (ΔW83), sialyltransferase defective mutants, was a kind gift from Dr. Charles Shelburne (University of Michigan, Ann Arbor, MI, USA). This strain is not able to incorporate sialic acid into its LPS and was constructed and validated in the same way as described by Sweier et al. 23 Briefly, to replace the PG0083 gene an ermF cassette was constructed using the following primers (5’-3’): ACTGAATCCGATGGTGCAACTG and TAAGGCGAGTCCGACAC- GATAG and was introduced into P. gingivalis W83 by electroporation. Similar to the wild type strain, W83 ΔPG0083 mutants were able to adhere and invade a cultured human cell line, but they were not pigmented and the disruption of the PG0083 gene had a significant effect on their growth.
Cell culture
THP-1 cells (human monocytic leukemia cell line) were purchased from ECACC (European Collection of Cell Cultures, Salisbury, UK) and maintained in RPMI 1640 medium (Invitrogen, Carslbad, CA, USA) supplemented with 2 mM glutamine, 10% FCS, penicillin (100 units/ml) and streptomycin (100 µg/ml) (Invitrogen). The cells were cultured between 103 and 106 cells/ml in a 5% CO2 atmosphere at 37℃.
Human gingival fibroblasts (HGF) were established from explants of healthy gingival tissues obtained during routine clinical procedures. 24 The study was approved by the Research Ethics Committee of Northern Ireland, participant information sheets were provided and written informed consent was obtained from patients wishing to participate in the study. Cells were cultured in DMEM (Invitrogen) supplemented with 10% heat-inactivated FCS, 2 mM glutamine, 100 U/ml penicillin and 100 µg/ml streptomycin in a humidified atmosphere of 5% CO2 at 37℃. Cells between passages 5 and 13 were used for assays. Twenty-four h before treatment the medium was changed to 1% FCS DMEM supplemented with the same concentration of antibiotics and glutamine.
SDS-PAGE
Whole-cell lysates were treated with sample loading buffer and fractionated on a 10% SDS polyacrylamide gel at 35 mA. Proteins were transferred from the gel to a nitrocellulose membrane using a Novex Mini Cell wet system apparatus (Invitrogen). The proteins were electrophoresed onto the membrane at 200 mA for 1.5 h.
Immunoblotting
Following protein transfer, nitrocellulose membranes were rinsed in PBS/0.1% Tween 20 and blocked with 5% milk for 1 h. Membranes were probed using respective primary Abs at the relevant dilutions in appropriate blotting solution. Subsequently, the membranes were washed six times for 5 min in PBS/0.1% Tween 20. Membranes were probed with an appropriate HRP-conjugated secondary Ab for 1 h in 5% milk dissolved in PBS/0.1% Tween 20 at 1:5000 concentration. The wash step was repeated as before and proteins were then detected using the chemiluminescence method. Mouse monoclonal anti-CD33 Abs were obtained from Abcam (Cambridge, UK) and rabbit anti-IκBα Abs from Cell Signaling (Danvers, MA, USA). To allow for visualization of proteins, membranes were covered for 5 min in chemiluminescence detection reagent Super Signal West-Femto (Thermo Fisher Scientific, Waltham, MA, USA). The Syngene G Box imaging system was used to expose blots as per manufacturer’s instructions. Images were analyzed using GeneSnap software.
Immunoprecipation
Two hundred μl Protein G magnetic beads (Millipore,) were pre-associated with 5 μl of phospho-tyrosine- 4G10 (Millipore, Billerica, MA, USA) Abs on a rotor for 90 min at 4℃. The Ab-bound beads were pelleted at 2500 g for 1 min and washed in 0.5% TBST. The whole-cell lysates were then added to the beads and incubated on rotation at 4℃ for 2 h. Afterwards, the beads were pelleted and washed three times in 0.5% TBST before being re-suspended in 5 μl 5 × Western loading buffer with 10% β-mercaptoethanol and 15 μl PBS. The samples were denatured at 95℃ for 5 min and placed on ice prior to loading onto a gel.
Lectin blot analysis
Sialic acid content in LPS was assessed using lectin blot analysis as described by Cao et al. 25 Two types of biotinylated lectins (Vector Laboratories, Burlingame, CA, USA) were used: Sambucus nigra lectin (SNA), which binds to α-(2→6)-sialic acid residues, and Maackia amurensis lectin (MAA), which interacts with α-(2→3)-linked sialic acid. Human serum was used as a positive control, ddH2O was used as a negative control.
One μl 1 mg/ml different LPS preparations (1 µg LPS, corresponding to 108 bacterial cells) with positive and negative controls were added directly onto the nitrocellulose membrane and allowed to air dry. The membrane was then incubated in 0.1% PBST for 1 h at room temperature. Lectins were dissolved in 0.1% PBST at a final concentration of 1 μl/ml and incubated with the membrane overnight at 4℃. The membrane was washed with PBST and incubated with Streptavidin HRP in PBST (1:5000). Having been washed, the membranes were developed using West-Femto chemiluminescence substrate and visualized in the Syngene G box.
ELISA
THP-1 cells were resuspended at 5 × 105 cells/ml and human gingival fibroblast at 8 × 104 cells/ml in corresponding media and treated with LPS for 4 h. Cell-free supernatants were removed and stored at –80℃ until assessed for TNF-α (PeproTech, Rocky Hill, NJ, USA) and IL-8 (R&D Systems, Minneapolis, MN, USA). Absorbance was read at 405 nm (Tecan GENios spectrofluorometer). Cytokines concentrations were extrapolated from the standard curve.
Flow cytometry
THP-1 cells and human gingival fibroblasts (1 × 106 cells/ml and 80,000 cells/ml, respectively) were washed in PBS, spun at 500 g for 5 min and re-suspended in a 100 µl PBS supplemented with 2% FCS. Cells were then stained with 20 µl FITC conjugated anti-human CD33 Ab (eBioscience, San Diego, CA, USA) or respective isotype control for 30 min at 20℃ in the dark. Cells were centrifuged again and washed twice with 1 ml PBS and re-suspended in 500 ml fresh PBS. Flow cytometry analysis was carried out on FACS CANTO II. Results were analyzed using FlowJo software (Tree Star Inc., Ashland, OR, USA).
Cell viability
Cell viability was determined with the Cell Proliferation Kit [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)] (Promega, Southampton, UK). THP-1 cells (5 × 105 cells/well) were treated with 1 µg/ml different LPS preparations for 4 h together with untreated control samples. Cells were subsequently incubated in a 96-well plate with MTT solution for a further 4 h. The absorbance of formazan dye was measured using a Tecan GENios plate reader, at 570 nm, with a reference wavelength of 690 nm.
Statistical analysis
Differences between the means of treatments were analyzed by the Student’s t-test using GraphPad Prism version 4 (GraphPad Software, San Diego, CA, USA). Differences between multiple treatments were compared by one-way ANOVA followed by Tukey’s post hoc test. Values are expressed as the mean ± SEM. A value of P < 0.05 was considered to represent a statistically significant difference.
Results
Sialic acid detection
Sialic acid appears as a terminal sugar in polysaccharide chains. In order to examine the content of sialic acid in LPSs from two different P. gingivalis strains (ATCC 33277 and W83), the lectin blot method was used with two sialic acid binding lectins as Abs [SNA and MAA specific for α-(2→6) and α-(2→3) bond between the sialic acid and the terminal N-acetyl galactosamine, respectively]. Human serum was used as a positive control because of the high concentration of sialylated proteins present in it, while LPS extracted from sialyltransferase-defective P. gingivalis W83 mutants (ΔW83) was used as a negative control. Lectin blot analysis revealed a high concentration of sialic acid within the W83 LPS and a much more attenuated signal for the ATCC 33277 strain. Both α-(2→6) and α-(2→3) linked sialic acids were abundantly present within the W83 LPS. Sialic acid was not detected in LPS extracted from P. gingivalis ΔW83 mutants (Figure 1).
Inflammatory potential of LPS
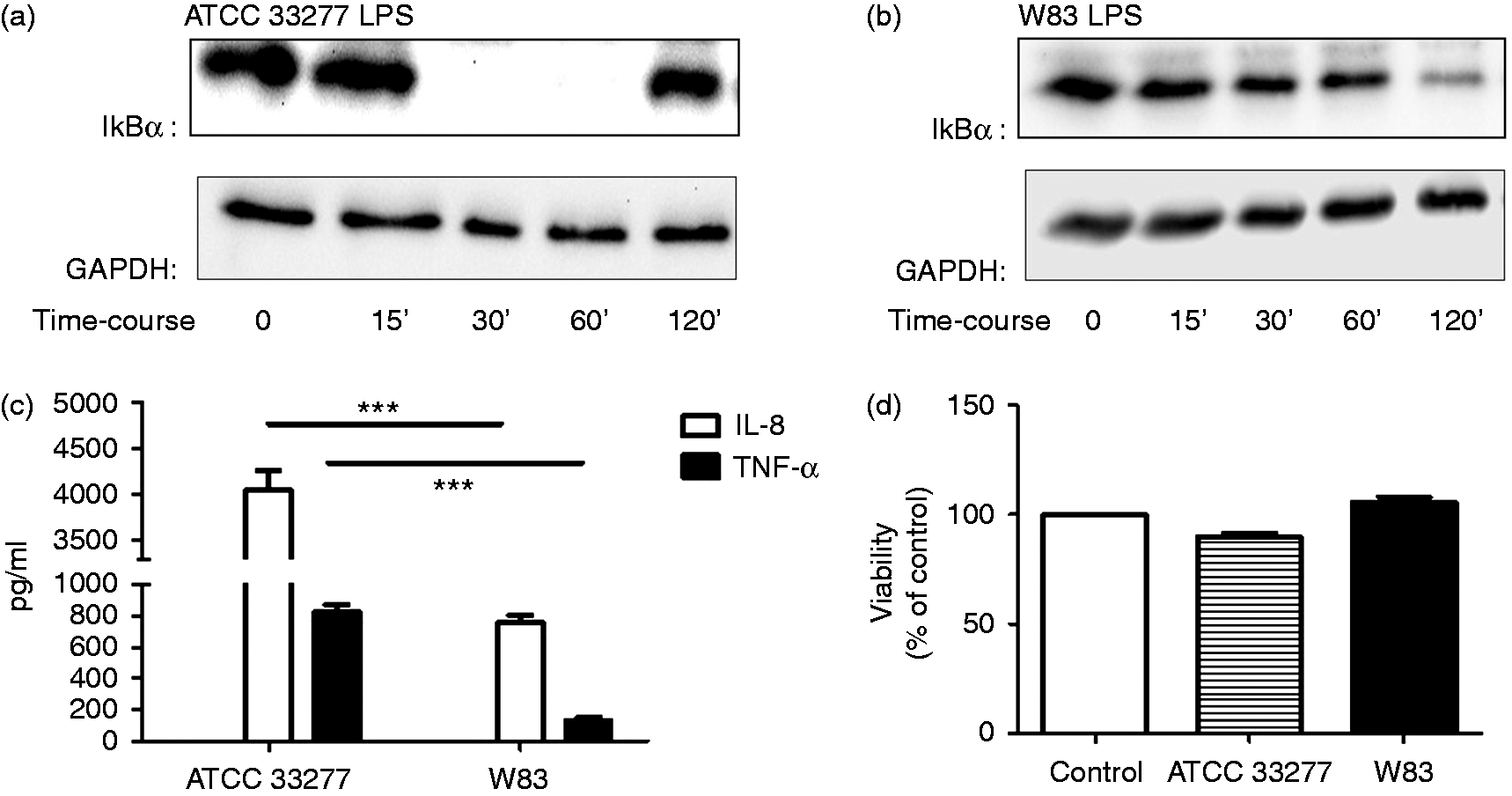
Inflammatory potential of the two types of P. gingivalis LPS was examined in a THP-1 cell model by monitoring IκBα degradation (Western blotting) after 15, 30, 60 and 120 min of LPS challenge, and by measuring TNF-α and IL-8 production in cells supernatants (ELISA) after 4 h. Complete IκBα degradation occurred as early as after 30 min in the cells treated by P. gingivalis ATCC 33277 LPS, while in the cells treated with P. gingivalis W83 LPS only partial degradation was observed after 2 h of treatment (Figure 2a, b). Similarly, P. gingivalis ATCC 33277 LPS triggered a signficantly higher production of both TNF-α and IL-8 compared with W83 strain LPS (P < 0.001) (Figure 2c). Cell viability was not affected by either of these two preparations (Figure 2d).
Lectin-blot detection of sialic acid LPS content from different P. gingivalis strains. (a) SNA lectin, (b) MAA lectin. 1: human serum; 2: P. gingivalis ATCC 33277; 3: P. gingivalis W83; 4: P. gingivalis ΔW83. The results are representative of three different experiments. Inflammatory potential of LPS extracted from P. gingivalis ATCC 33277 and P. gingivalis W83. (a, b) THP-1 cells were challenged with 1 µg/ml (a) ATCC 33277 LPS or (b) W83 LPS for 15, 30, 60 and 120 min. GAPDH was used as a loading control. (c) Production of TNF-α and IL-8 by THP-1 cells challenged with the two P. gingivalis LPS isoforms for 4 h. (d) Cell viability measured by the MTT assay. Values represent means ± SEM (n = 3).
The influence of LPS sialylation on its inflammatory potential
In order to examine the influence of sialic acid LPS moieties on the pro-inflammatory response of innate immune cells, THP-1 cells were treated for 4 h with different concentrations of P. gingivalis W83 LPS, which was shown to be highly sialylated, and LPS isolated from sialyltransferase gene (PG00083) knockout P. gingivalis W83 strain (ΔW83), and the production of TNF-α and IL-8 was measured in the supernatants (Figures 3 and 4).
IL-8 production by THP-1 cells treated with different concentrations of P. gingivalis W83 LPS and P. gingivalis Sia-free ΔW83 LPS. Values represent the mean ± SEM (n = 3). TNF-α production by THP-1 cells treated with different concentrations of P. gingivalis W83 LPS and P. gingivalis Sia-free ΔW83 LPS (n = 3).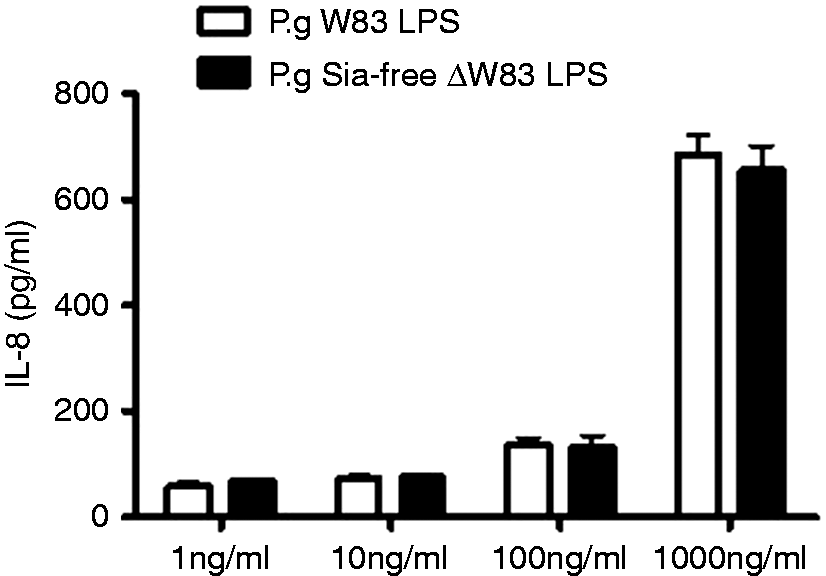
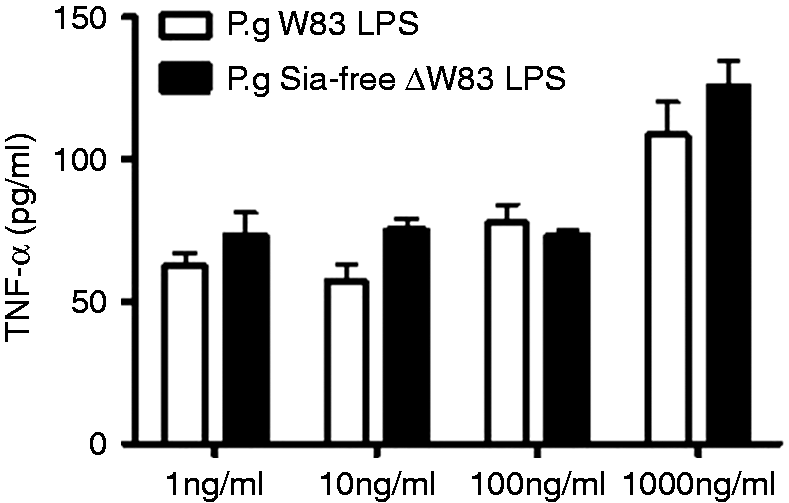
There was no difference in IL-8 and TNF-α levels produced by THP-1 cells treated with sialic acid free P. gingivalis ΔW83 LPS (Sia-free LPS) compared with sialic acid containing P. gingivalis W83 LPS in all concentrations examined.
CD33 activation
Upon activation of CD33, the tyrosine in position 340 in its intracellular tail becomes phosphorylated and is able to recruit SHP-1, SHP-2 or SHIP. THP-1 cells were treated with P. gingivalis ATCC 33277 LPS, P. gingivalis W83 LPS and P. gingivalis ΔW83 LPS for 4 h, and phosphorylation of tyrosine residues within CD33 molecules was examined. Immunoprecipitation of THP-1 whole-cell lysates with anti-phoshotyrosine Abs was carried out followed by immunoblotting for CD33 (Figure 5).
CD33 phosphorylation. THP-1 cells were treated with: (1) medium alone; (2) 1 µg/ml P. gingivalis ATCC 33277 LPS; (3) 1 µg/ml P. gingivalis W83 LPS; and (4) 1 µg/ml P. gingivalis ΔW83 LPS and the cell lysate was immunoprecipitated with anti-phosphotyrosine Abs and immunoblotted for CD33. Whole-cell lysate immunoblotted for CD33 served as a loading control. The results are representative of three different experiments.
The results revealed that CD33 was phosphorylated at tyrosine residues in the cells treated with all types of LPSs in addition to control cells incubated in the medium alone. These results show that CD33 is endogenously expressed in THP-1 cells and activated regardless of the type of LPS treatment. This is possibly due to the presence of sialic acids at the surface of monocytes as a constituent of glycoproteins, which interact with CD33 in cis interaction and block trans interaction of sialic acid present in LPS and CD33. Constitutive repressor activity of self-expressed sialic acid with CD33 receptors may be important for maintaining low levels of pro-inflammatory cytokine production during the resting state of the immune system.
CD33 inhibits spontaneous secretion of IL-8
As HGFs do not express CD33 (Figure 6b), the spontaneous secretion of IL-8 by THP-1 cells, HGFs and neuraminidase (NA)-treated THP-1 cells and HGFs over 4 h without any treatment was investigated. Half a million of THP-1 cells, NA-treated THP-1 cells and 8 × 104 HGFs and NA-treated HGFs per 1 ml of corresponding media were incubated for 4 h and IL-8 levels in cell supernatants were determined by ELISA. The concentration of IL-8 was adjusted according to the cell number (Figure 6c). Spontaneous production of IL-8 was significantly higher in NA-treated THP-1 cells and human gingival fibroblasts compared with THP-1 cells, suggesting that endogenous sialic acid-CD33 interaction is likely to play an important role in inhibiting basal secretion of pro-inflammatory cytokines. NA treatment of HGFs did not affect the level of spontaneous IL-8 production by these cells.
CD33 expression in (a) THP-1 cells and (b) HGFs. (c) Spontaneous production (over 4 h) of IL-8 by THP-1 cells, NA (Sigma, St. Louis, MO, USA) treated THP-1 cells, HGF and NA-treated HGFs, adjusted according to cell number used in the experiments (n = 3).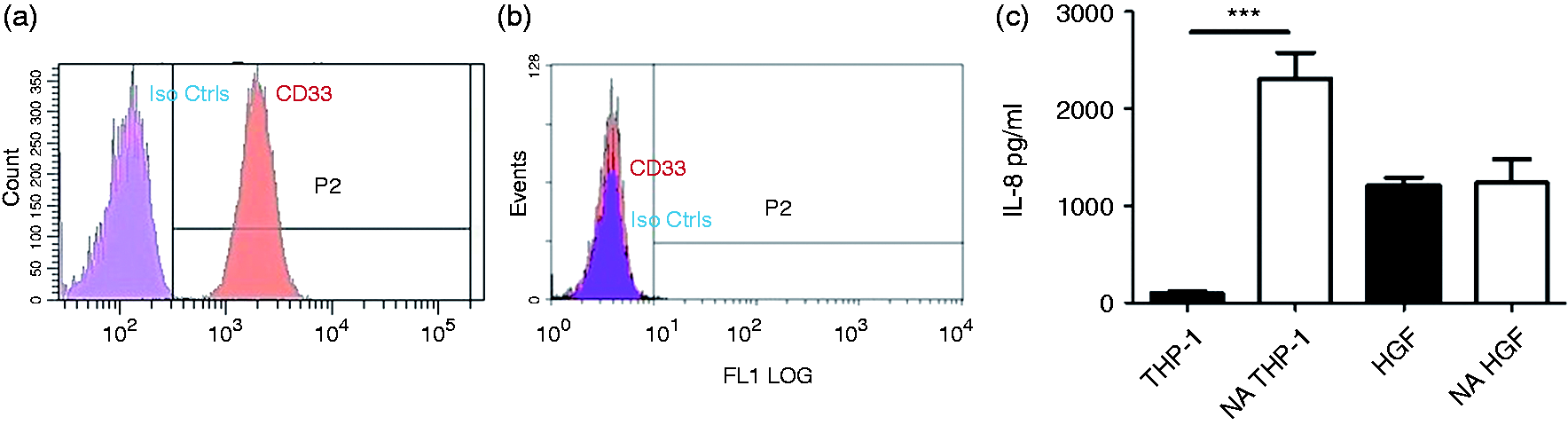
Discussion
Sialic acid residues are highly expressed in eukaryotic cells and act as a marker for self in the immune system. 26 Pathogenic bacteria also use this molecule for biological mimicry, to coat themselves in sialic acid and increase resistance to components of the host’s innate immune response, or they can use it as nutrient. 27 Structural variations of bacterial LPS can have a major impact on the host immune response and clinical disease outcome. Sialylation of LPS is an important mechanism used by the human pathogen H. influenzae to evade the innate immune response of the host and escape killing by human serum, 28 while the inflammatory response against Campylobacter jejuni is in direct correlation with the level of its LPS sialylation. 29 In addition, it has been shown that the invasive strains of N. meningitidis have highly sialylated LPS compared with carrier strains and that these structural features are likely to be important in the ability of the disease-causing bacteria to evade recognition by LPS-specific bactericidal IgG and complement-mediated lysis. 30 Here, we show for the first time, that P. gingivalis W83 LPS is highly sialylated, containing both α-(2→6) and α-(2→3) attached sialic acid moieties, while P. gingivalis ATCC 33277 LPS expresses a much lower level of sialylation. Many previous studies have compared virulence-associated activities and disease-promoting characteristics between P. gingivalis strains and have found the ATCC 33277 strain to be avirulent, while the W83 strain is generally considered virulent. In addition, a cluster of genes involved in the synthesis of highly sialylated capsular polysaccharides, transposase, helicase and fibronectin type 3 are present in W83, while it was not found in strain ATCC 33277. 31
We compared the inflammatory potential of these two types of P. gingivalis LPS and found a rapid IκBα degradation and much higher production of TNF-α and IL-8 in human monocytes challenged by LPS extracted from the ATCC 33277 strain compared with W83 LPS. While the measurement of TNF-α and IL-8 is not a comprehensive assessment of inflammatory potential, they have been shown to be good indicators of periodontal tissues damage. 32 Detection of LPS by TLR expressed on innate immune cells triggers a robust and essential inflammatory reaction. The interaction between LPS and TLR components depends on the chemical composition of the lipid A core of the LPS molecule (level of phosphorylation and acylation) and can be modulated by the O-antigen polysaccharide tail. 33 In our case, highly sialylated W83 LPS triggered a significantly lower inflammatory response in THP-1 cells compared with less sialylated ATCC 33277 LPS. Similarly, sialylation of Helicobacter bizzozeronii LPS has been shown to play a crucial role in attenuation of the host’s inflammatory response to this prominent animal pathogen. 34
P. gingivalis is capable of synthesizing heterogeneous isoforms of the lipid A molecule, which are subtly different from each other in structure. These subtle changes in the lipid A chemical composition have been shown to greatly influence the host’s immune response to P. gingivalis LPS. 35 In order to examine the influence of sialic acid O-antigen moieties of LPS on the host’s inflammatory response and exclude lipid A variations, sialyltransferase-deficient W83 mutants were grown under the same conditions as the wild type W83 and their LPS was extracted using the same method. These mutants produced sialic acid-free LPS that triggered a similar level of TNF-α and IL-8 in THP-1 cells as the wild type LPS, suggesting that sialic acid is not an important determinant of P. gingivalis LPS inflammatory potency.
When CD33 receptor activation by LPS extracted from ATCC 33277, W83 and ΔW83 strains was examined by immunoblotting, CD33 was found to be activated in cells treated with all three types of LPS in addition to the control, untreated cells. This is probably a result of the inability of sialylated P. gingivalis LPS to displace and outcompete endogenously expressed sialic acid on the surface of THP-1 cells from cis-mediated interaction with CD33 receptors. Ishida et al. have found that CD33 activation by endogenous ligands negatively regulates TLR intracellular signaling and production of IL-12. 36 Similarly, pre-treatment of sialic acid has been shown to efficiently prevent LPS-induced acute renal failure and TLR- mediated apoptotic signaling in a rat model. 37 On the contrary, desialylation of dendritic cells significantly improves their phagocytic activity and immunological potency through an actin-independent mechanism. 38 Interestingly, hyperglycemia down-regulates CD33 expression in human monocytes, and higher levels of TNF-α, IL-8 and IL12p70 were detected in the plasma of patients with type 2 diabetes compared with healthy individuals. 39 In addition, an increase of α-(2→6)- and α-(2→3)-sialylation has been observed on the surface of endotoxin-tolerant macrophages that responded to a repeated LPS challenge with significantly reduced production of TNF-α. 40
It is not possible to induce endotoxin tolerance in HGFs, 41 and they have been shown to play a crucial role in maintaining chronic inflammation. 42 We showed here that HGFs do not express CD33 receptors and their basal (spontaneous) IL-8 production was much higher than in THP-1 cells. Interestingly, the spontaneous IL-8 production was significantly increased in THP-1 cells treated with NA, suggesting an important role of endogenous sialic acid–CD33 interaction in preventing production of pro-inflammatory cytokines during the resting state of the immune system.
Our data, in combination with previous studies, suggest that although sialylation of P. gingivalis cell surface components may provide additional benefits to this prominent periodontal pathogen in biofilm formation and escaping complement killing, it does not affect the inflammatory potential of its LPS. While sialylation of pathogens’ surface ligands and attenuation of the host inflammatory response might reflect immune evasion by pathogens, it may also represent a protective response that serves to prevent excessive inflammation in the host. Interactions between oral pathogens and Siglecs is a field of investigation that is likely to continue expanding in scope and dental clinical practice importance.
Footnotes
Acknowledgements
We are immensely grateful to Dr Charles Shelburne, University of Michigan, School of Dentistry, for sharing his sialic acid-free P. gingivalis LPS with us.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.