
Abstract
Septic cardiomyopathy affects up to 70% of patients with septic shock and the derangement of cardiac mitochondrial function contributes to the likelihood of death. However, at present, there is no specific therapeutic drug available. The peroxisome proliferator-activated receptor (PPAR)-γ coactivator-1α (PGC-1α) and coactivator-1β (PGC-1β) modulate members of the PPARs, which regulate mitochondrial energy metabolism and the production of mitochondrial reactive oxygen species in the heart. This study investigated the potential of the newly developed synthetic antimicrobial peptide 19-2.5 (Pep2.5) to attenuate mitochondrial dysfunction in murine cardiomyocytes stimulated with serum from septic shock patients. Pep2.5 treatment attenuated the suppression of PPAR-α, PPAR-γ (P = 0.0004 and P = 0.0001, respectively) and PGC-1α/β (P = 0.0008 and P = 0.0147, respectively) in cardiomyocytes stimulated with serum from septic shock patients compared with untreated cells. Pep2.5 treatment enhanced the mitochondrial maximum respiration (P < 0.0001), increased cellular ATP levels (P < 0.0001) and reduced the production of mitochondrial reactive oxygen species. Thus, the administration of Pep2.5 may have the potential as a promising therapeutic approach in septic cardiomyopathy by attenuating mitochondrial dysfunction in the septic heart.
Introduction
Sepsis-associated myocardial dysfunction is referred to as septic cardiomyopathy (SIC) and affects up to 70% of patients with septic shock. The presence of SIC indicates a worse prognosis, with mortality rates up to 90%. 1 For several years, a multitude of pathomechanisms have been reported as substantial contributors to myocardial depression, particularly concerning the disturbance of cardiomyocyte energy metabolism.2,3 Derangement of cardiac mitochondrial function leads to several pathological mechanisms, such as depletion of ATP and excessive production of mitochondrial reactive oxygen species (mtROS). 4 In the heart, mitochondrial function is controlled by members of the peroxisome proliferator-activated receptors (PPARs), which are modulated by PPAR-γ coactivator-1α (PGC-1α) and coactivator-1β (PGC-1β). 5 Both, PAMPs, such as LPS, and danger-associated molecular patterns (DAMPs), e.g. heparan sulfate, down-regulate PGC-1α and PGC-1β expression and cause impaired cardiac energy metabolism on the basis of a reduced mitochondrial substrate flux.6,7 Several animal and human studies report a strong correlation between the impairment in cardiac mitochondrial function and the extent of myocardial contractile dysfunction in sepsis.8–10 Thus, there is increasing interest in the development of new therapeutics attenuating mitochondrial dysfunction in SIC.
The synthetic antimicrobial peptide 19-2.5 (Pep2.5) has been developed as a synthetic anti-LPS peptide; however, its activity is not restricted to PAMPs.11–13 Pep2.5 reduces inflammation in cardiomyocytes stimulated with human sepsis serum by interacting with both PAMPs and DAMPs. 12 Moreover, the systemic administration of Pep2.5 limits inflammation and lengthens survival in a murine model of polymicrobial sepsis.14,15 Thus, we hypothesized that Pep2.5 attenuates mitochondrial dysfunction in murine cardiomyocytes stimulated with human sepsis serum (SsP). Our data demonstrate for the first time that Pep2.5 enhances mitochondrial respiration, increases ATP levels and attenuates the production of mtROS in murine cardiomyocytes stimulated with SsP by attenuating the suppression of PPARs and PGC-1α/β.
Materials and methods
Serum sampling
We used sera of patients (n = 10) consecutively admitted to the intensive care unit with septic shock, according to the Third International Consensus Definitions for Septic Shock. 16 All patients, or their legal representative, gave written informed consent. Individuals younger than 18 yr were excluded. All samples are stored in the RWTH centralized Biomaterial Database (RWTH cBMB) of the University Hospital RWTH Aachen. Before inclusion of the first individual, the local ethics committee (University Hospital RWTH Aachen, EK 206_09) approved this study, which was performed in accordance with the Declaration of Helsinki.
Heparan sulfate ELISA
The amount of heparan sulfate in serum of patients with septic shock was determined using ELISA (AMS Biotechnology, Abingdon, UK), as described previosuly. 7 The absorbance was measured at 450 nm on a microplate reader (Sunrise Tecan, Crailsheim, Germany).
Peptide synthesis
Peptide 19-2.5 was synthesized as described before.14,15 Briefly, solid-phase peptide synthesis technique in an automatic peptide synthesizer (model 433 A; Applied Biosystems, Foster City, CA, USA) on Fmoc-Rink amide resin was used. The amino acid sequence of Pep2.5 is GCKKYRRFRWKFKGKFWFWG, with a molecular mass of 2711 kDa.
Cardiomyocytes cell culture
HL-1 cardiomyocytes originally purchased from William Claycomb, Louisiana State University, were cultured as described before.12,17 HL-1 cells were plated on six-well plates for 48–72 h. Four h prior measurements SsP (5%) with or without Pep2.5 (20 µg/ml) were added to the Claycomb medium.
Measurement of oxygen consumption
Oxygen consumption rate (OCR) was assessed by a Seahorse XF96 analyser (Seahorse Bioscience, North Billerica, MA, USA), as described before. 7 Briefly, HL-1 cells were grown in 96-well XF plate (Seahorse Bioscience) up to a cell density of 33,000 cells per well. Cells were incubated for 1 h in assay medium (DMEM supplemented with 4.5 g/l Glc, 2 mM sodium pyruvate and 2 mM glutamine). Four h after stimulating the cells with SsP in the presence or absence of Pep2.5, OCR was measured in pMol/min before and after the addition of inhibitors to obtain several parameters of mitochondrial respiration. First, baseline cellular OCR was measured. Second, oligomycin (1 µM), a complex V inhibitor, was added. Third, carbonyl cyanide-p-trifluoromethoxyphenyl-hydrazon (FCCP; 0.25 µM) was administered, allowing the electron transport chain to function at its maximal rate (maximal respiratory capacity). Finally, antimycin A and rotenone (1 µM each), inhibitors of complex III and I, were added, revealing non-mitochondrial respiration. Results represent the data from five measurements per each time point. The reported values are compared to unstimulated cells and given as relative OCR.
Measurement of cellular ATP content
The Mouse Adenosine Triphosphate ELISA-Kit (Bluegene, Shanghai, China) was used, according to the manufacturer’s instructions to measure relative cellular ATP content. Absorbance was measured at 450 nm on a microplate reader (Sunrise Tecan). Results represent the data from three independent experiments performed in triplicates.
Mitochondrial ROS and confocal microscopy
As described recently, 7 live-cell imaging was carried out by plating HL-1 cardiomyocytes on ibiTreat u-Slide 8 wells (Ibidi, Germany) in Hank's Balanced Salt Solution. The cells were incubated with 5 µM MitoSOX Red (Invitrogen, M36008) and 5 µg/ml Hoechst 33342 (Invitrogen, H3570) and imaged at 37℃ and 5% CO2 in the cell incubator of a LSM 710 confocal microscope (Carl Zeiss, Jena, Germany) using a 40 × , 1.1 NA water immersion objective.
RNA extraction and PCR
Total RNA was extracted as described before. 12 For quantitative real-time PCR the following primers were used: PGC-1α 5′ CGGAAATCATATCCAACCAG 3′ (for) and 5′ TGAGGACCGCTAGCAAGTTTG 3′ (rev); PGC-1β 5′ TCCAGAAGTCAGCGGCCT 3′ (for) and 5′ CTGAGCCCGCAGTGTGG 3′ (rev); PPARα 5′ AACATCGAGTGTCGAATATGTGG 3′ (for) and 5′ AGCCGAATAGTTCGCCGAAAG 3′ (rev); PPARγ 5′ CCACCAACTTCGGAATCAGC 3′ (for) and 5′ AGTGGTCTTCCATCACGGAGGA 3′ (rev). Ribosomal Protein S7 was used as an endogenous normalization control: 5′ GGTGGTCGGAAAGCTATCA 3′ (for) and 5′ AAGTCCTCAAGGATGGCGT 3′ (rev). Results represent the data of triplicate samples, representative of three independent experiments.
Western blot analysis
Immunoblot analysis have been performed as described recently, using mouse anti-PPARα (Santa Cruz Biotechnology, Santa Cruz, California) and anti-PPARγ (Santa Cruz Biotechnology). 7 The bound Abs were detected with Amersham ECL Prime (GE Healthcare Bio-Sciences AB, Uppsala, Sweden) and analyzed on a LAS-4000 system (GE Healthcare Bio-Sciences AB).
Determination of secreted cytokines in HL-1 cells
The amounts of IL-6 (NOVEX, San Diego, CA) and TNF-α (Invitrogen, Camarillo, CA) were determined in the cell supernatant collected 4 h subsequent to stimulation by ELISA according to the manufacturer’s instructions. The absorbance was measured at 450 nm on a micro plate reader (Sunrise Tecan, Crailsheim, Germany).
Statistical analyses
Continuous data are described by means ± SD relative to unstimulated cells. The PCR-derived data were analyzed using a relative expression software tool (REST, http://www.gene-quantification.de/rest.html, rest-mcs-beta-9august 2006). 18 The expression ratios are calculated on the basis of the mean crossing point values for reference and target genes. We used a multiple t-test with Holm-Šídák correction when comparing differences between two groups. We used a one-way-ANOVA and Tukey’s test for multiple comparisons when comparing more than two groups. P-Values ≤ 0.05 were regarded as statistically significant. We performed all calculation and constructed all figures using GraphPad Prism 6 (GraphPad, San Diego, CA, USA).
Results
Study population characteristics and proof of concept
Septic shock patients (n = 10; 50% male) from whom the sera were obtained were 73 ± 7 yr old. All patients met criteria for septic shock according to the Third International Consensus Definitions for Septic Shock. 16 Patients had an Acute Physiology and Chronic Health Evaluation II score (APACHE II) of 23 ± 7. None of the patients died during the 28-d study period. The level of circulating heparan sulfate (HS) in the patients’ sera was 154 ± 37 µg/ml. In all cases, patients had positive blood cultures with either Gram-negative (n = 2) or Gram-positive bacteria (n = 3), mycoplasma cells (n = 3), or polymicrobial results (n = 2). The healthy volunteers had a mean age of 67 ± 19 yr (50% male). As a proof of concept for the present study, we investigated the impact of Pep2.5 on the inflammatory response and cellular ATP levels in cardiomyocytes exposed to normal serum from healthy volunteers and medium alone. Neither the addition of Pep2.5 to normal serum nor to medium alone resulted in any inflammatory response, as we were unable to detect the pro-inflammatory cytokines IL-6 and TNF-α in the cell supernatants of these groups. Moreover, we measured no alteration in relative cellular ATP levels in HL-1 cells exposed to normal serum from healthy volunteers, compared with unstimulated cells (relative fold change to unstimulated cells = 0.95 ± 0.12; P = 0.8682). Similarly, the addition of Pep2.5 to HL-1 cells exposed to serum from healthy volunteers or medium alone did not influence cellular ATP levels [relative fold change to unstimulated cells = 1.02 ± 0.09 (P = 0.7492) and 0.94 ± 0.17 (P = 0.7918), respectively].
Pep2.5 enhances mitochondrial maximum respiration
On a functional level we measured OCR, which serves as an indicator for mitochondrial respiration. The OCR is given as relative value to unstimulated cells (Figure 1). Treatment with Pep2.5 did not influence basal respiration compared with cells incubated with SsP alone (P > 0.05; Figure 1a). The complex V inhibitor oligomycin was added to quantify OCR not linked to ATP synthesis. Pep2.5 treatment did not alter OCR values compared with untreated cells (P > 0.05; Figure 1b). Next, the mitochondrial uncoupler FCCP was added to collapse the inner membrane gradient, allowing the electron transport chain to function at its maximal rate, which is a surrogate of mitochondrial reserve capacity for causes of high energy demand. Relative OCR was significantly increased in HL-1 cells treated with Pep2.5 and SsP, compared with cells stimulated with SsP alone (P < 0.0001; Figure 1c). As the final step, complex III and I inhibitors antimycin A and rotenone were added to shut down the electron transport chain function and determine the non-mitochondrial OCR. Treatment with Pep2.5 did not influence non-mitochondrial respiration, compared with untreated cells incubated with SsP (P > 0.05; Figure 1d).
Effect of Pep2.5 treatment on mitochondrial respiration. HL-1 cells were stimulated for 4 h with serum from SsP (5%) in the absence (dotted line) or presence of Pep2.5 (20 µg/ml, solid line). (a) Initially, baseline cellular relative OCR was measured. (b) Complex V inhibitor oligomycin (1 mM), (c) the mitochondrial uncoupler FCCP (0.25 mM) and (d) the complex III and I inhibitor antimycin and rotenone (each 1 mM) were added. Data are presented relative to unstimulated cells as means ± SD from five measurements. P-Values represent statistical difference between untreated and Pep2.5-treated cells. n.s.: not significant.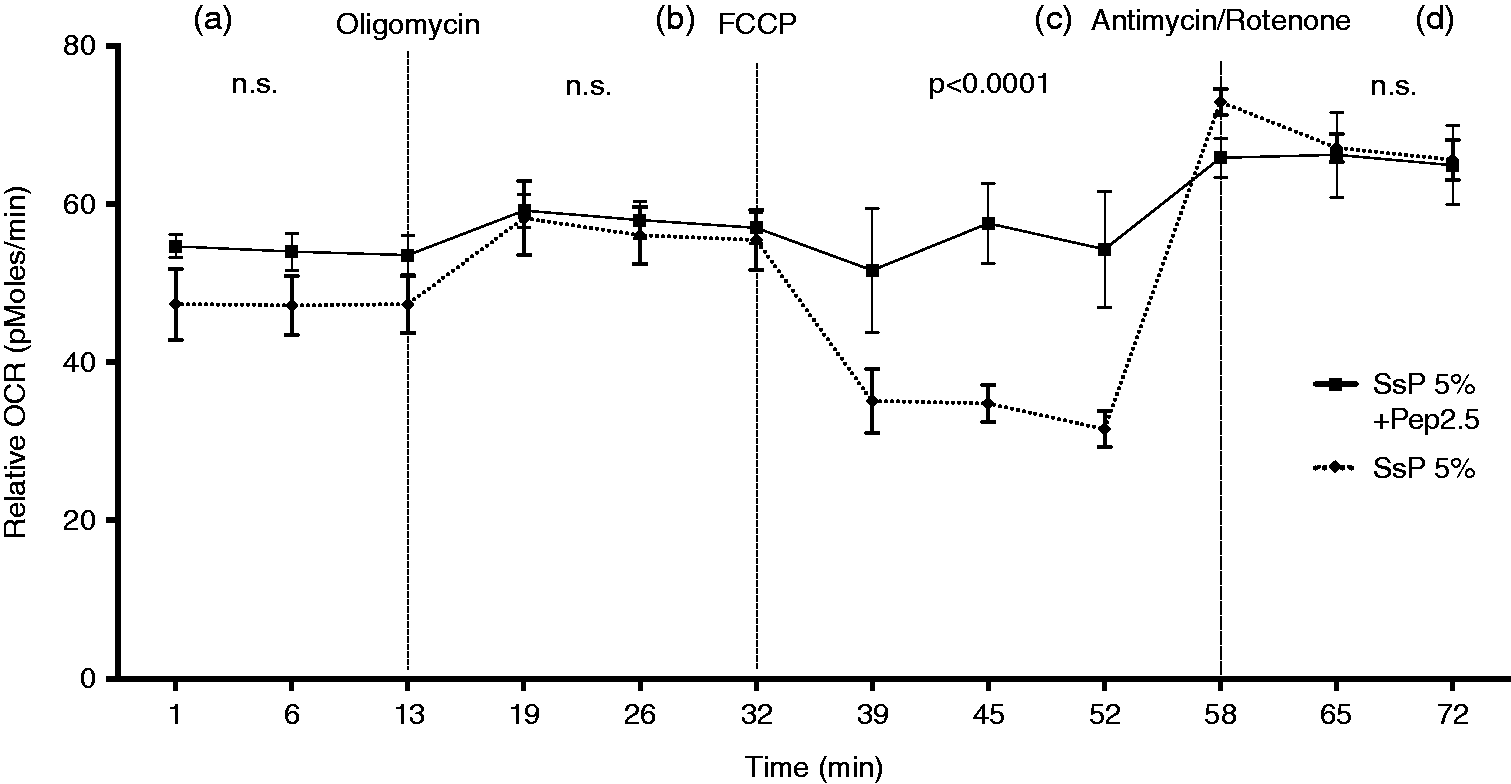
Pep2.5 attenuates ROS production
mtROS in HL-1 cells stained with MitoSOX were visualized by confocal microscopy. Figure 2a shows representative image sections of MitoSOX red fluorescence generated by superoxide radicals in HL-1 cells. Quantitative fluorescence intensity was analyzed by ImageJ software (NIH, Bethesda, MD, USA) and shown in Figure 2b. HL-1 cells stimulated with SsP (Figure 2a, upper left panel), showed an increased MitoSOX fluorescence compared with unstimulated cells (P < 0.0001; Figure 2a, upper right panel). Pep2.5-treatment (Figure 2a, lower left panel) decreased MitoSOX fluorescence intensity compared with untreated SsP-stimulated cells (P < 0.0001; upper left panel). Antimycin A was used as a positive control and is shown in Figure 2 (lower right panel).
Effect of Pep2.5 treatment on mtROS levels. HL-1 cells were stimulated for 4 h with serum from SsP (5%) in the absence or presence of Pep2.5 (20 µg/ml). (a) After staining the cells with MitoSOX (5 µM) and Hoechst 33342 (5 µg/ml) cellular fluorescence intensity was detected by confocal microscopy. Top left: SsP-stimulated cells with no specific treatment (medium); top right: unstimulated cells with no specific treatment; bottom left: SsP-stimulated cells with Pep2.5 treatment; bottom right (positive control): unstimulated cells with antimycin A (50 μM). (b) Quantitative fluorescence intensity was analyzed by ImageJ software and is given relative to unstimulated cells as mean ± SD. Results are representative of three independent experiments performed in triplicate. #P < 0.05 vs. untreated SsP-stimulated cells.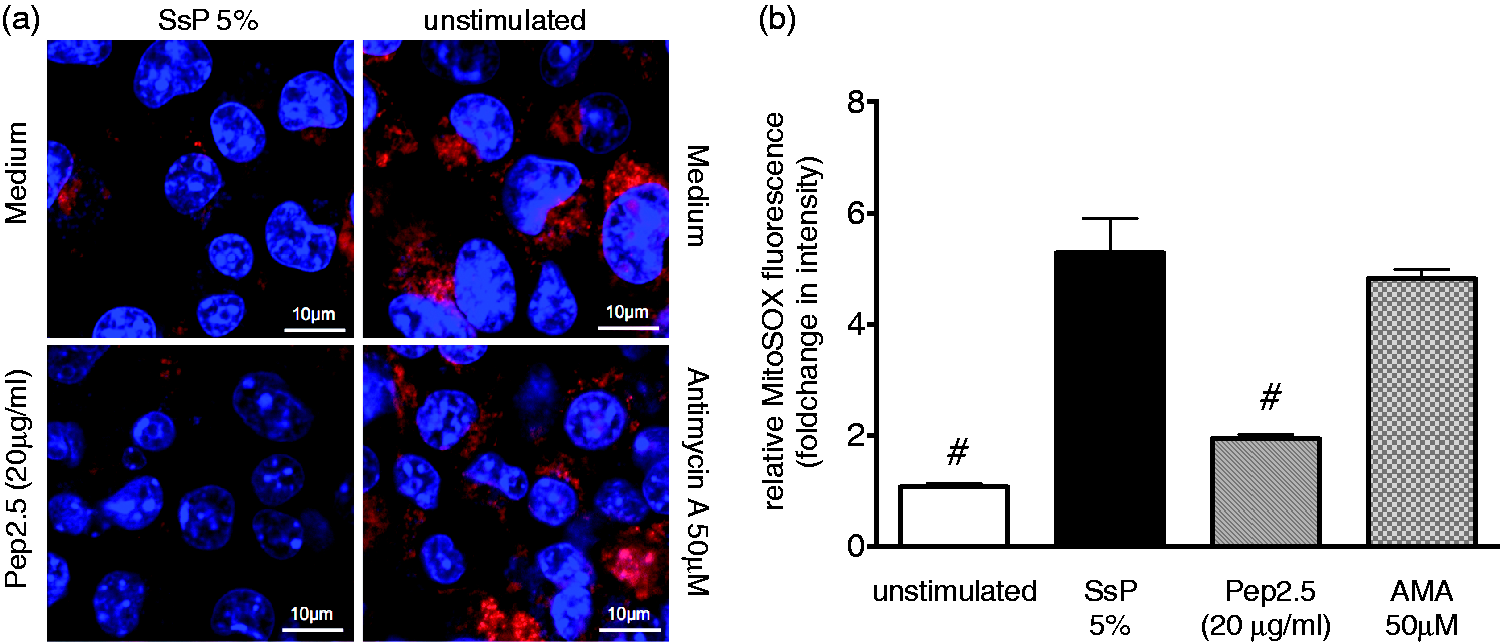
Pep2.5 improves cellular ATP levels
Additionally, we investigated cellular ATP concentrations in HL-1 cells (Figure 3). Relative cellular ATP level significantly increased in HL-cells treated with Pep2.5 compared with SsP-stimulated cells without treatment (P < 0.0001; Figure 3).
Effect of Pep2.5 treatment on cellular ATP content. HL-1 cells were stimulated for 4 h with serum from SsP (5%) in the absence or presence of Pep2.5 (20 µg/ml). Cellular concentrations of ATP were measured using a mouse-specific ATP ELISA. Data are presented relative to unstimulated cells as mean ± SD. Results are representative of three independent experiments performed in triplicate. #P < 0.05 vs. untreated cells.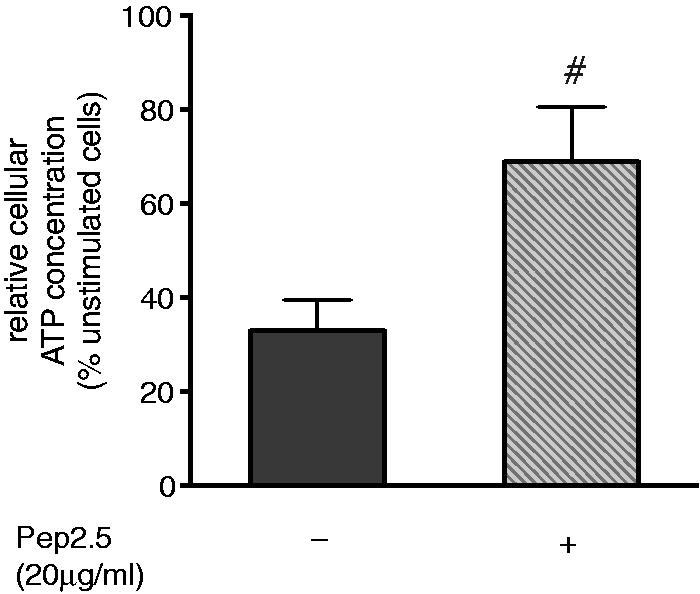
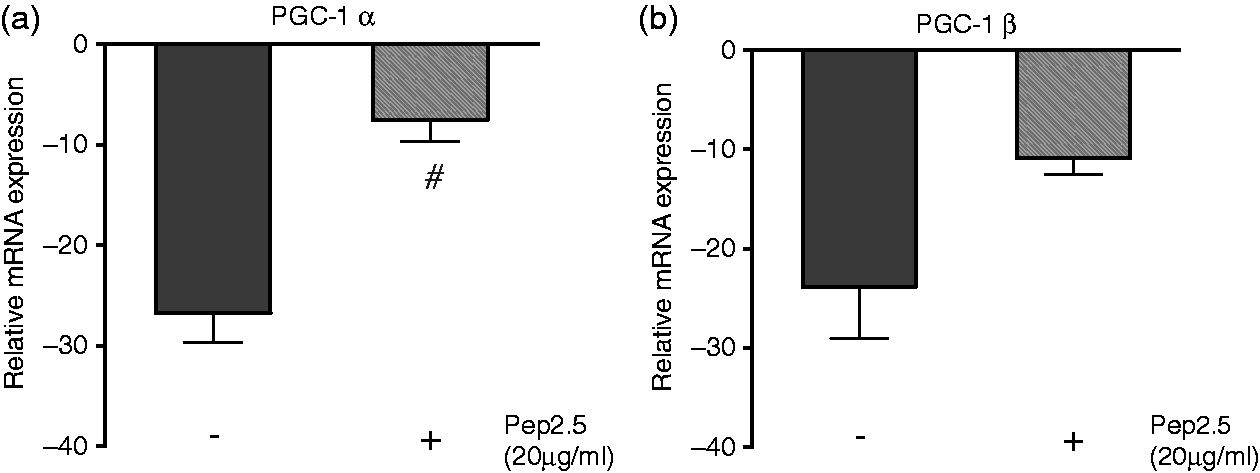
Pep2.5 reduces suppression of PPARs and PGC-1α/β induced by human sepsis serum
To investigate the effect of Pep2.5 on PPAR-γ and PPAR-α, we measured their relative mRNA expression and protein levels in SsP-stimulated HL-1 cells (Figure 4). Treatment with Pep2.5 resulted in a significant up-regulation of PPAR-γ and PPAR-α mRNA compared with cells stimulated with SsP alone (P = 0.0001 and P = 0.0004, respectively; Figure 4a, b). Consistently, treatment with Pep2.5 resulted in a significant up-regulation of PPAR-γ and PPAR-α protein levels, compared with untreated SsP-stimulated cells (P = 0.0118 and P = 0.0009, respectively; Figure 4c, d). As the PPAR-γ coactivators PGC-1α and PGC-1β modulate the activity of PPARs, we also investigated the effects of Pep2.5 treatment on PGC-1α and PGC-1β mRNA levels (Figure 5). HL-1 cells stimulated with SsP and treated with Pep2.5 revealed significantly reduced suppression of PGC-1α and PGC-1β mRNA compared with SsP-stimulated cells without treatment (P = 0.0008 and P = 0.0147, respectively; Figure 5a, b).
Effect of Pep2.5 treatment on PPAR-γ and PPAR-α. HL-1 cells were stimulated for 4 h with serum from SsP (5%) in the absence or presence of Pep2.5 (20 µg/ml). The expression of mRNAs encoding the peroxisome proliferator-activated receptors (a) PPARγ and (b) PPARα were determined by RT-PCR. The protein levels of (c) PPAR-γ and (d) PPAR-α were determined by Western blot. Densitometric analysis of the bands is expressed as relative OD normalized to their corresponding Vinculin band. The values shown are normalized to unstimulated cells. Results are representative of three independent experiments performed in triplicate and presented as means ± SD. #P < 0.05 vs. untreated cells. Effect of Pep2.5 treatment on PGC-1α and PGC-1β. HL-1 cells were stimulated for 4 h with serum from SsP (5%) in the absence or presence of Pep2.5 (20 µg/ml). The expression of mRNAs encoding (A) PGC-1α (a) and (b) PGC-1β were determined by RT-PCR. The values shown are normalized to unstimulated cells. Results are representative of three independent experiments performed in triplicate and presented as means ± SD. #P < 0.05 vs. untreated cells.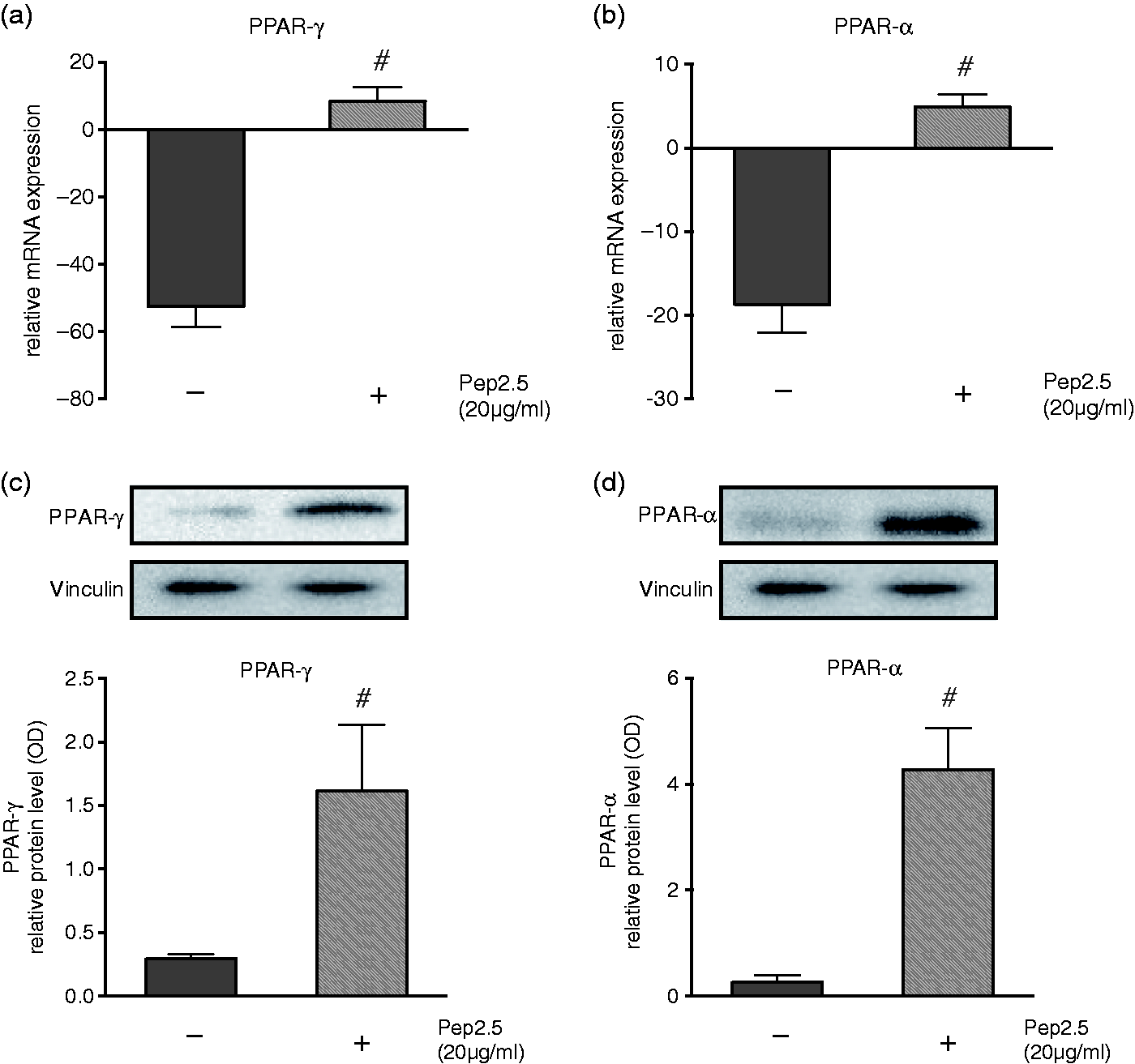
Discussion
In this study, we investigated the therapeutic potential of the synthetic antimicrobial peptide 19-2.5 (Pep2.5) to attenuate mitochondrial dysfunction in cardiomyocytes stimulated with human sepsis serum. Our data indicate that Pep2.5 enhances mitochondrial respiration, increases ATP levels and attenuates the production of mtROS in SsP-stimulated cardiomyocytes by attenuating the suppression of PPARs and PGC-1α/β.
In fact, there are two possible mechanisms responsible for these findings. First, Pep2.5 may not target mitochondrial function per se, but rather modulate cellular response to pro-inflammatory mediators by interacting with circulating pathogen [e.g. LPS; lipoproteins/lipopeptide (LP); lipoteichoic acids (LTA); and peptidoglycans (PGN)] and endogenous cardio-depressants (e.g. heparan sulfate) in the serum. Second, Pep 2.5 may reduce mitochondrial dysfunction by reducing the production of negatively charged mtROS, thereby enhancing mitochondrial function. 19
Enhanced mitochondrial respiration in Pep2.5-treated cardiomyocytes
We recently reported that the continuous parenteral administration of Pep2.5 reduces cytokine release and tissue inflammation in mice subjected to polymicrobial sepsis. 14 In particular, the mRNA expression of the TLR4 co-receptor CD14 is significantly down-regulated in the heart of Pep2.5-treated mice. 14 The importance of TLR4 for mitochondrial function was emphasized in an in vitro model of C2C12 skeletal muscle cells, where TLR4 activation reduced mitochondrial OCR. 6 Notably, CD14-deficient mice experienced no ventricular dysfunction and decreased cytokine mRNA expression after LPS challenge. 20 Both PAMPs and DAMPs are known to act as potent TLR agonists. 21 In this way, both PAMPs and DAMPs suppress PPARs and PGC-1α/β expression in the heart and cause impaired cardiac energy metabolism due to reduced mitochondrial substrate flux.6,7 Circulating heparan sulfate, one of the major DAMPs in serum of septic patients, reduces the maximum respiration after uncoupling from ATP formation by FCCP. However, cells stimulated with heparan sulfate-free serum reveal unaltered basal and maximum respiration compared with unstimulated cells. 7 Of note, Pep2.5-treatment resulted in a significantly increased maximum OCR of cardiomyocytes exposed to serum from septic shock patients suffering from different kinds of infections (i.e. Gram-negative or Gram-positive bacteria, mycoplasma cells and polymicrobial results) compared with untreated cells (Figure 1c). Importantly, Pep2.5 exhibits strong anti-inflammatory activity against both PAMPs and DAMPs. In this way, Pep2.5 inhibits the cytokine release induced by LPS and various non-LPS compounds (e.g. LP, LTA, PGN), as well as heparan sulfates.12,22 It is known that the positively charged Pep2.5 binds to the negatively charged heparan sulfate resulting in a change of their sulfation states, 13 . One could speculate that this change contributes to the inhibiting effects of Pep2.5 on the binding of heparan sulfates to TLR4. However further studies are needed, this could happen in a similar modus by which Pep2.5 changes the aggregate structure of LPS preventing it from binding to TLR4, 11 which decrease the LPS induced PGC-1α and PGC-1β down-regulation. 23 Of note, Pep2.5 interacts with heparanase, the major mammalian endoglycosidase, which cleaves heparan sulfate fragments from their proteoglycans in a more charge-independent, at least partially hydrophobic character. 24
Reduced mtROS and higher ATP levels in Pep2.5 treated cardiomyocytes
As mitochondria are the main producers, as well as targets, of mtROS, the positively charged Pep2.5 may reduce mtROS both by reducing the production of the negatively charged mtROS through its anti-inflammatory activity, as well as by the indirect regulation of mitochondrial function via PPARs and PGC-1α/β. 19 The production of mtROS in the setting of SIC can be viewed as a double-edged sword because mtROS play a crucial role in the defense against xenobiotics and foreign pathogens. However, enhanced mtROS production results in the activation of intrinsic apoptotic pathways as a response to cellular damage. 25 Indeed, naturally occurring antimicrobial peptide are known to play a prominent role in maintaining the balance between the defensive and the cytotoxic capacity of mtROS. 25 Small, synthetic, positively charged basic peptides scavenge ROS directly; however, there have been no studies in models of sepsis, only in ischemia/reperfusion.26,27 One can only speculate that the positively charged synthetic Pep2.5 scavenges ROS in a similar way. However, further in vitro and in in vivo studies are needed to clarify this important issue. Moreover, mtROS induce cardiac inflammation via the generation of circulating heparan sulfate through ectodomain shedding and release of mitochondrial DNA, which both serve as potent DAMPs. 4 This, in turn, leads to even more mitochondrial damage, resulting in a vicious cycle of increasing mtROS levels. 19 In addition to regulating PPARs, the PGC-1 coactivators also control the expression of some enzymes capable of scavenging ROS, 28 which also may explain our findings of lower mtROS levels after Pep2.5 treatment (Figure 2a), with higher levels of PPARs (Figure 4) and PGC-1α/β (Figure 5).
The generation of ATP in the heart mainly depends on fatty acid oxidation, which is regulated by the nuclear receptors PPARα/γ and PGC-1α/β. 29 If ATP levels drop too low, cell death is triggered. It has been shown that the expression of these receptors are down-regulated during polymicrobial sepsis, 30 and that transgenic mice over-expressing PPARγ are protected from LPS-induced cardiac dysfunction by enhancing the production of cellular ATP. 23 The same holds true for the over-expression of PGC-1β. 23 We measured significantly higher cellular ATP concentrations in Pep2.5-treated HL-1 cells (Figure 3), with higher levels of PPARs (Figure 4) and PGC-1α/β (Figure 5). Low cellular ATP content is associated with higher mortality in critically ill patients and is found in non-survivors of sepsis. 31 It was hypothesized that cells turn into a state of hibernation at which they can retain their structural integrity with little ATP usage, albeit losing their normal function. 32 However, during septic shock high levels of cardiac ATP are needed to maintain adequate organ function. 33 Although further animal studies are needed, Pep2.5 treatment may help to maintain the required ATP content in cardiomyocytes, thus improving cardiac function during septic shock.
Conclusion
In summary, our study demonstrates the therapeutic potential of the synthetic antimicrobial peptide Pep2.5 to attenuate mitochondrial dysfunction in cardiomyocytes stimulated with human sepsis serum. Our data indicate that Pep2.5 enhances mitochondrial respiration, increases ATP levels and attenuates the production of mtROS in SsP-stimulated cardiomyocytes by attenuating the suppression of PPARs and PGC-1α/β. Although our findings are limited to in vitro analyses and further studies are needed to clarify effects of Pep2.5 on cardiac dysfunction in vivo, our data suggest a potential of Pep2.5 as a new therapeutic drug in SIC by attenuating mitochondrial dysfunction in septic cardiomyocytes.
Footnotes
Acknowledgments
We thank Dr Claycomb and his co-workers for providing the HL-1 cells and a detailed documentation. The Immunohistochemistry and Confocal Microscopy Unit, a core facility of the Interdisciplinary Centre for Clinical Research (IZKF) Aachen, within the Faculty of Medicine at the RWTH Aachen University, supported this work.
Declaration of Conflicting Interests
The author(s) declared following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: LM has received grants by the Faculty of Medicine at the RWTH Aachen University (START 15/14 and START 46/16) and the Deutsche Forschungsgemeinschaft (DFG, MA 7082/1-1). GM has received honoraria for consulting or lecturing and restricted research grants from the following companies: BBraun, Edwards Life Sciences, Serumwerke Bernburg, Hutchinson Technology, Baxter. KB has a patent for the structure of the synthetic antimicrobial peptide 19-2.5 (Aspidasept, Brandenburg Antiinfektiva, Borstel, Germany): Patent-No: PCT/EP2009/002565, and is chief scientific officer of Brandenburg Antiinfektiva GmbH. TS received travel grants and lecture fees from Astellas Pharma, and lecture fees from Bayer Vital, Astra-Zeneca and B. Braun Melsungen, and is chief medical officer of Brandenburg Antiinfektiva GmbH. All the other authors declare no conflicts of interest
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by intramural grants to Dr Lukas Martin (START 15/14 and START 46/16) and the RWTH centralized Biomaterial Database (RWTH cBMB) of the University Hospital RWTH Aachen.