
Abstract
Neonatal innate immunity is distinct from that of adults, which may contribute to increased susceptibility to infection and limit vaccine responses. B cells play critical roles in protection from infection and detect PAMPs via TLRs, that, when co-activated with CD40, can drive B-cell proliferation and Ab production. We characterized the expression of TLRs in circulating B cells from newborns and adults, and evaluated TLR- and CD40-mediated naïve B-cell class-switch recombination (CSR) and cytokine production. Gene expression levels of most TLRs was similar between newborn and adult B cells, except that newborn naïve B cells expressed more TLR9 than adult naïve B cells. Neonatal naïve B cells demonstrated impaired TLR2- and TLR7- but enhanced TLR9-mediated cytokine production. Significantly fewer newborn naïve B cells underwent CSR to produce IgG, an impairment also noted with IL-21 stimulation. Additionally, co-stimulation via CD40 and TLRs induced greater cytokine production in adult B cells. Thus, while newborn naïve B cells demonstrate adult-level expression of TLRs and CD40, the responses to stimulation of these receptors are distinct. Relatively high expression of TLR9 and impaired CD40-mediated Ig secretion contributes to distinct innate and adaptive immunity of human newborns and may inform novel approaches to early-life immunization.
Introduction
B cells play a critical role in protection from infectious diseases. While they are best known for their contribution to adaptive immunity via production of soluble immunoglobulins (Abs, Igs), innate responses of B cells also affect the subsequent adaptive immune response. 1 B cells express TLRs, 2 a family (10 members in humans) of innate PRRs that recognize conserved microbial molecular patterns and initiate MyD88-dependent and MyD88-independent signaling events, 3 and stimulation of B cell TLRs can induce cytokine production and influence their proliferation and activation. 4 In particular, B cells express TLR9, which is responsive to CpG motif-rich bacterial DNA and to synthetic CpG DNA. 5
While maternal Abs provide some protection from infection at birth, endogenous neonatal B-cell responses to immunization and infection are often inadequate, and as circulating levels of maternal IgG diminish with age (half-life ∼21–30 d), this contributes to a general susceptibility to infection. 6 Overall, innate immune responses of murine and human neonatal leukocytes such as monocytes and dendritic cells are different from adult counterparts,7,8 with characteristic impaired inflammatory/Th1-polarizing cytokine (e.g. TNF-α and IL-12p70) production, while anti-inflammatory cytokines, such as IL-10, are elevated.9–11
Multiple studies have described the ontogeny of murine B-cell function. 6 For example, murine neonatal B-cell production of IL-10 suppresses pro-inflammatory responses of dendritic cells thereby inhibiting Th1-polarizing immune responses.12,13 Immune responses can be species-specific, and relatively little is known regarding human neonatal B-cell-mediated innate immune responses. Only limited data are available regarding the expression of TLRs on human neonatal B cells, and regarding the responsiveness of human neonatal B cells to TLR agonists. Two studies have reported that human cord blood B cells respond to CpG stimulation by producing cytokines and up-regulating genes that are involved in plasma cell differentiation.14,15 Because TLR agonists may be used as vaccine adjuvants and characterization of age-specific TLR function may inform adjuvanted vaccine development, we sought to more completely characterize the expression of TLRs in human circulating B cells and B-cell subsets from both newborn and adult subjects, and to evaluate TLR-mediated cytokine production and Ig class-switch recombination by circulating newborn (cord blood) and adult (peripheral blood) B cells. To characterize B-cell ontogeny, our approach entailed comparing newborn and adult total naïve B cells, as this subset represents the vast majority of all neonatal B cells, 16 and has been the subject of prior studies in this area.17–19 We have found that neonatal naïve B cells exhibit distinct functional expression of TLRs with relatively high TLR9 expression and TLR9-mediated cytokine induction and impaired CD40-mediated responses, including Ig production and class switching, providing fresh insights into the ontogeny and innate function of this key leukocyte population.
Materials and methods
Blood collection
Peripheral blood was collected after informed consent was obtained from healthy adult volunteers (mean age 32.4 yr) according to Boston Children’s Hospital institutional review board-approved protocols, and newborn cord blood (mean gestational age 39.1 wk) was collected immediately after elective cesarean section delivery (epidural anesthesia) of the placenta. Births to HIV-positive mothers were excluded. Human experimentation guidelines of the US Department of Health and Human Services, the Brigham and Women’s Hospital, Beth Israel Medical Center and Boston Children’s Hospital were observed, following protocols approved by the local institutional review boards. Number of repeats (n) indicates the number of independent experiments. No subject was studied more than once in each of the different experiments. Blood was collected into syringes containing a final concentration of 20 U/ml pyrogen-free heparin (Sagent Pharmaceuticals, Schaumberg, IL, USA), and processed within 2 h of collection.
FACS
FACS was utilized to acquire highly pure B-cell subsets for analysis of TLR mRNA expression. Mononuclear cells were stained with Abs targeting CD19 (APC-Cy7), CD24 (PE-Cy7), CD27 (PerCP-Cy5.5), CD38 (BV-605) and IgD (BV-421). Cells were sorted using a FACSAria II cell sorter (BD Biosciences, Frederick, MD, USA). The gating strategy has been previously described. 16 In brief, B cells were CD19+, and subsets were naïve B cells (IgD+, CD27–), early memory (IgD+, CD27+), class-switched memory (IgD−, CD27+, CD38+/−) and plasmablasts (IgD−, CD27+, CD38++). B-cell subpopulations were sorted into tubes containing RPMI 1640 media (Invitrogen, Carlsbad, CA, USA) supplemented with 10% FBS (HyClone; VWR, Radnor, PA, USA), centrifuged at 500 g for 10 min and cell pellets were re-suspended in Buffer RLT (Qiagen GmbH, Hilden, Germany) for immediate RNA isolation. We seldom recovered sufficient adult plasmablast cells or any subset in newborn samples (save naïve) for subsequent work-up, and they were not included in this analysis.
Mononuclear cell isolation and magnetic bead naïve B-cell isolation
PBMC or cord blood mononuclear cells (CBMC) were isolated from heparinized whole blood by Ficoll density gradient centrifugation. Naïve B cells were isolated by negative selection. Non-naïve B cells (CD27+ B cells, T cells, NK cells, monocytes, dendritic cells, granulocytes, platelets and erythroid cells) were labeled with a cocktail of biotinylated CD2, CD14, CD16, CD27, CD36, CD43 and CD235a Abs and magnetically labeled with Anti-Biotin MicroBeads for depletion (Naïve B Cell Isolation Kit II, human; Miltenyi Biotec, Auburn, CA, USA). To improve purity, the isolated naïve B-cell fraction was subsequently labeled with CD19 microbeads (Miltenyi Biotec) for positive selection, resulting in highly pure naïve B-cell populations. Only samples with naïve B-cell purities of >90% were used for assays (average purity CD20+ IgD+ was 96% for adults and 95% for cord blood isolates), and cell isolates were also evaluated by flow cytometry (LSR Fortessa; BD Biosciences) to evaluate for specific impurities (CD27+ IgD+ cells < 0.2%, CD27+ IgD− < 0.04%, CD3+ and CD14+ both < 0.6%, see ‘Flow cytometry’ section for more details). Flow cytometry analysis of a representative sample is shown in Supplementary Figure 1. Panel A indicates the percentage of CD20+ events, panel B the low frequencies of CD14+ and CD3+ events, and panel C shows the percentage of CD20+ cells that are IgD+ and CD27−. For select experiments, total CD19+ B cells were isolated from adult PBMC fractions utilizing CD19 microbeads (Miltenyi Biotec), for which the purity (CD20+) was always > 90%, and the average specific subsets or impurities were CD3+ (1.5%) and CD14+ (0.7%).
Flow cytometry
For analysis of TLR expression on B-cell subsets, mononuclear cells were stained with CD19 (APC-Cy7), CD27 (PerCP-Cy5.5), CD38 (BV-605), IgD (BV-421) and TLR4 (FITC). For staining of intracellular TLRs, cells were subsequently fixated with 4% paraformaldehyde (Alfa Aesar, Ward Hill, MA, USA) in PBS for 30 min at 4℃. Fixated cells were washed in 1 × BD Perm/Wash (BD Biosciences) and stained with TLR7 (PE) and TLR9 (APC) in 1 × Perm/Wash. Stained cells were washed with Perm/Wash and subsequently with PBS before measurement of fluorescent intensity on an LSR Fortessa (BD Biosciences). For purity analysis of B cells isolated using magnetic beads, cells were stained with CD20- (PE-Cy7), CD3- (FITC), CD14- (APC-Cy7), CD27- (PerCP-Cy5.5), CD40- (APC) and IgD- (BV-421) specific Abs, and subsequently re-suspended in PBS with 1% paraformaldehyde before measurement of fluorescent intensity on an LSR Fortessa. Fluorescent intensities were analyzed using Flowjo software version 10 (Tree Star Inc., Ashland, OR, USA).
Cell culture
Isolated B-cell populations were plated in round-bottom 96-well plates (Corning, Tewksbury, MA, USA) in 100 µl RPMI 1640 media (Invitrogen, Carlsbad, CA, USA) supplemented with 10% FBS (HyClone; VWR), and stimulated with indicated reagents prepared in an additional 100 µl media to achieve the described reagent concentrations, and were incubated at 37℃ in a 5% CO2 incubator for the indicated assay-specific amount of time before utilization.
RNA purification and cDNA synthesis
Total RNA was isolated from sorted B-cell subpopulations using the RNeasy Mini Kit with RNase-free DNase treatment (Qiagen). Up to 500 ng mRNA was reverse-transcribed to cDNA using the RT2 First-strand Kit (SABiosciences, Frederick, MD, USA), according to the manufacturer's instructions.
qRT-PCR
Expression levels of selected genes were assessed by qRT-PCR analysis using an ABI 7300 real-time PCR system machine and software (Applied Biosystems, Foster City, CA, USA). The baseline adjustment method of the ABI 7300 software was used to determine the cycle threshold (Ct) in each reaction. A melting curve was constructed for each primer pair to verify the presence of one amplicon-specific peak and the absence of primer dimerization. All samples were amplified in triplicates and the mean was used for further analysis. Relative expression of TLR mRNA was compared with that of the ‘housekeeping’ genes hypoxanthine phosphoribosyltransferase1 (HPRT1) and β-actin (ACTB) using the ΔΔ-Ct method. Relative comparison between populations was similar for each housekeeping gene, and results shown here are for comparison with HPRT1, the expression levels of which were more similar to the expression levels of TLR mRNA.
Reagents
The following commercially available reagents were used at the concentrations indicated in the figure legends: heat-killed Listeria monocytogenes (HKLM, TLR2) R848 (TLR7/8), CpG (ODN 2395, TLR9) (InvivoGen, San Diego, CA, USA), IL-2 (R&D Systems, Minneapolis, MN, USA), human recombinant CD40L (MegaCD40L; Enzo Life Sciences, Farmingdale, NY, USA) and recombinant human IL-21 (Cell Signaling, Danvers, MA, USA).
B-cell IgM and IgG ELISpot
At the end of the incubation period, 96-well membrane plates (Cat# MSIPS4W; Millipore, Billerica, MA, USA) were coated with anti-human IgG or IgM Abs diluted in PBS (Cat# SELB002 – IgG, SELB003 – IgM; R&D Systems). After 24 h incubation at 4℃, the plates were blocked at room temperature (20℃) with PBS and 1% BSA for 2 h. Plates were washed with PBS and 104 (newborn IgM, adult IgM and adult IgG) or 105 (newborn IgG and adult IgG) B cells were added to duplicate wells and incubated at 37℃ overnight (10–14 h). Plates were subsequently washed three times with PBS/0.5%Tween and then three times with PBS. Kit isotype-specific detection Abs were added and further incubated at 4℃ for 24 h. Plates were washed three times with PBS and streptavidin HRP (Invitrogen, Carlsbad, CA, USA) was added for 30 min at room temperature. After washing plates with PBS, 50 µl TMB (Cat# 3651-10; Mabtech, Cincinnati, OH, USA) substrate was added. When spots were clearly developed (15–30 s), plates were washed quickly with distilled water. Spots were counted using an ELISpot plate reader (CTL, Shaker Heights, OH, USA). When spot counts were below the limits of detection, a value of half the lower limit of detection was assigned.
Cytokine measurement by multi-analyte fluorescent bead-based array
The cytokine profile of B-cell culture supernatants was analyzed using multi-analyte bead array (Millipore). Results were obtained with a MAGPIX system with Luminex xPONENT software (both from Luminex Corp., Austin, TX, USA). Cytokine concentrations were determined using Milliplex Analyst (version 3.5.5.0; Millipore) software. When cytokines were below the limits of detection, a value of half the lower limit of detection for that particular cytokine was assigned.
Statistical analysis
The indicated statistical tests, as described in the figure legends, were performed using GraphPad Prism version 5.0 b for Mac (GraphPad Inc., La Jolla, CA, USA).
Results
TLR expression by newborn and adult circulating B cells
mRNAs encoding TLRs 1, 2 and 4–10 were detected in circulating naïve B cells, isolated by FACS, from both adult and newborn subjects; only TLR3 was not detected (Figure 1A). The highest levels of transcription were detected for TLRs 6, 7, 9 and 10, with moderate levels of TLRs 1, 2 and 4, and low levels of TLR5 and TLR8. There were no statistically significant differences in mRNA expression of any individual TLR between populations (newborn and adult), although there was a trend towards elevated TLR9 in neonatal naïve B cells compared with adult counterparts. Protein expression of select TLRs (4, 7 and 9) was also evaluated by flow cytometry, which indicated modestly elevated expression of TLR4 and TLR9 on newborn naïve B cells relative to adult counterparts (Figure 1B). Comparisons were made within each experiment (at least two adults in each run), owing to variable mean fluorescent intensity between experiments.
TLR expression by adult and newborn circulating B cells. (A) Naïve B-cell TLR mRNA expression is similar in newborn subjects (n = 7) relative to adults (n = 9): no statistical significance, one-way ANOVA with Bonferroni post-test correction. (B) Flow cytometry evaluation of naïve B-cell expression relative to adult subjects from the same experimental run (mean fluorescent intensity, n = 6 each population) demonstrated greater TLR4 and TLR9, but similar TLR7 expression, in the newborn. Paired Student’s t-tests, *P < 0.05, **P < 0.01, ***P < 0.001.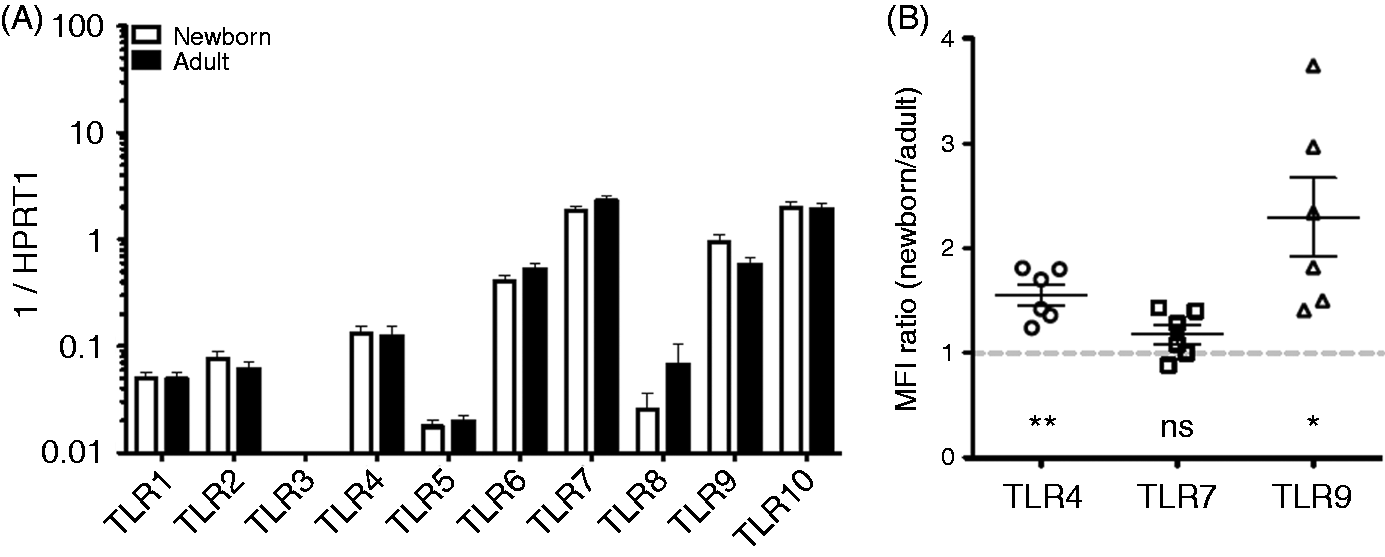
Adult B cells were also sorted into subpopulations, and TLR mRNA expression evaluated, revealing statistically significant increases in TLR4, TLR6 and TLR9 with maturation from naïve B cells to class-switched (IgD–) memory B cells (Supplementary Figure 2A). Protein expression of TLRs 4, 7 and 9, was also evaluated by flow cytometry on these subpopulations, which indicated increased expression for each with progression to IgD+ memory cells and IgD− memory cells (Supplementary Figure 2B).
TLR-Mediated B-cell cytokine production
Newborn and adult naïve B cells and adult total CD19+ B cells isolated by magnetic bead assay were stimulated with agonists for TLR2 (HKLM), TLR7/8 (R848), TLR9 (CpG), a combination of R848 and CD40L, or vehicle control, and cultured for 24 h in a CO2 incubator at 37℃ before evaluation of culture supernatants for a panel of cytokines utilizing a multiplex assay. Results from naïve cells for IL-10, CCL3 (also known as MIP-1α), IL-6 and TNF-α are shown in Figure 2, while data for all measured cytokines under these conditions, including CXCL8 (IL-8), IP-10, GM-CSF, IFN-α2, IFN-γ, MCP-1, IL-12p40, IL-12p70 and IL-1β, are shown in Supplementary Tables 1 and 2. Following stimulation of TLR2 or TLR7, neonatal naïve B cells produced generally lower or similar levels of cytokines than adult counterparts, with significantly lower levels of IL-6 following TLR7 activation and lower levels of TNF-α following TLR2 activation. In contrast, treatment with CpG (TLR9) elicited significantly higher levels of IL-10 and TNF-α from newborn naïve B cells than from adult naïve B cells (Figure 2). Total CD19+ B cells (adult) secreted considerably higher levels of some cytokines (IL-6, IL-8, MCP-1, CCL3 and TNF-α) than naïve B cells from the same subjects (Supplementary Tables 1 and 2).
Distinct TLR-mediated cytokine production by adult and newborn circulating B cells. Naïve B cells were stimulated with HKLM (109 cells/ml), R848 (10 µM), CpG (1 µM) or vehicle control for 24 h before evaluation of supernatant cytokines by multiplex assay. Neonatal B cells demonstrated impaired TLR2- and TLR7- but enhanced TLR9-mediated responses. n = 6–8, Student’s t-tests, *P < 0.05, **P < 0.01.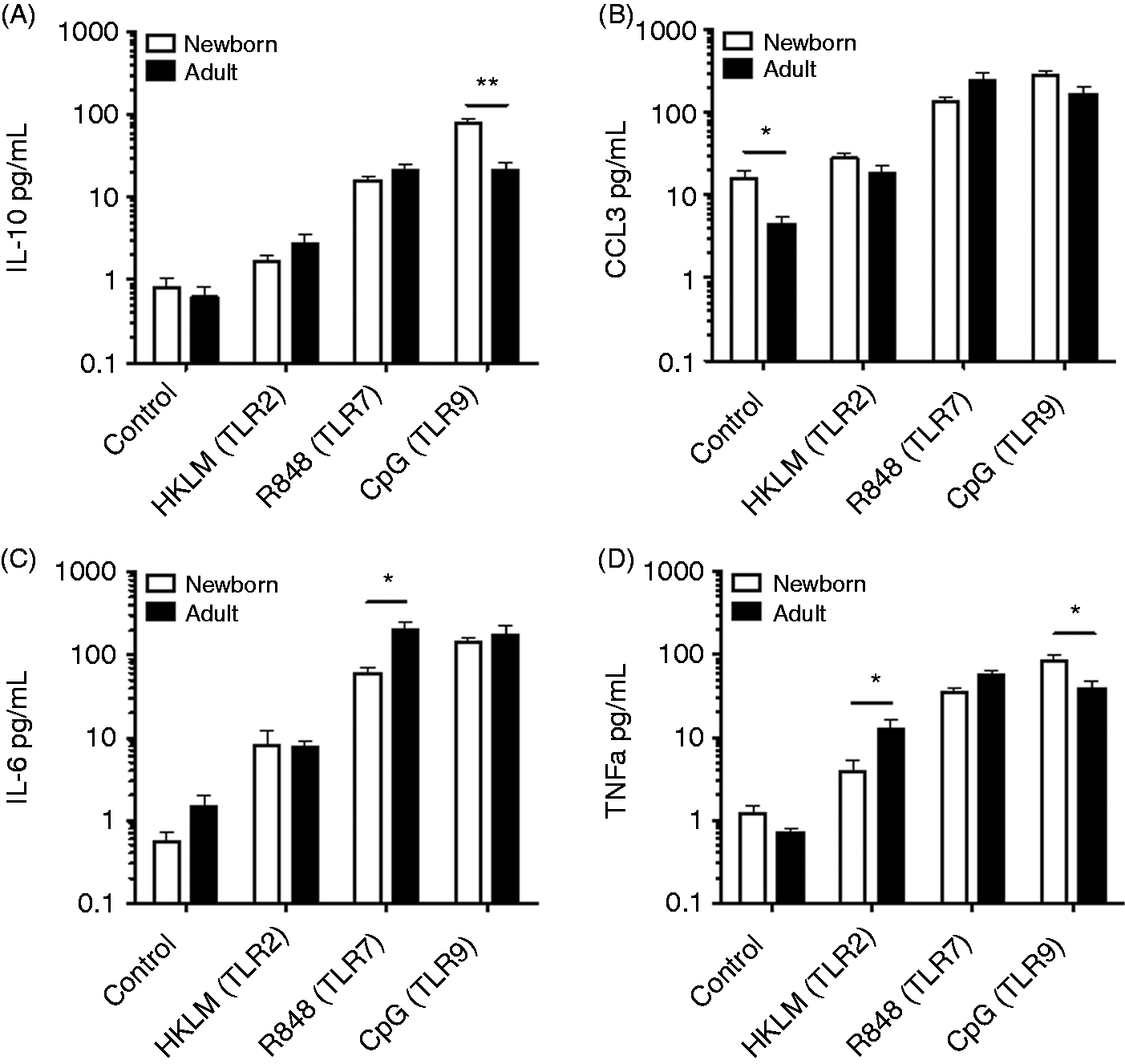
CD40 stimulation modulates B-cell cytokine production
CD40 signaling plays a critical role in B-cell biology, particularly for induction of class-switch recombination. Accordingly, we next evaluated how CD40 stimulation influenced TLR-mediated cytokine production from naïve B cells. CD40L caused significantly enhanced production of IL-10, IL-6 and TNF-α during TLR7/8 stimulation with R848 (Figure 3), although to a lesser extent in newborn naïve B cells than in adult naïve B cells, suggesting differences in CD40 signaling between these populations.
Impaired CD40 enhancement of cytokine production in newborn circulating B cells. Naïve newborn and adult B cells were stimulated with R848 (10 µM) or R848 plus recombinant CD40L (20 ng/ml) for 24 h before evaluation of supernatant cytokines by multiplex assay. n = 6–8, Student’s t-tests, *P < 0.05, **P < 0.01.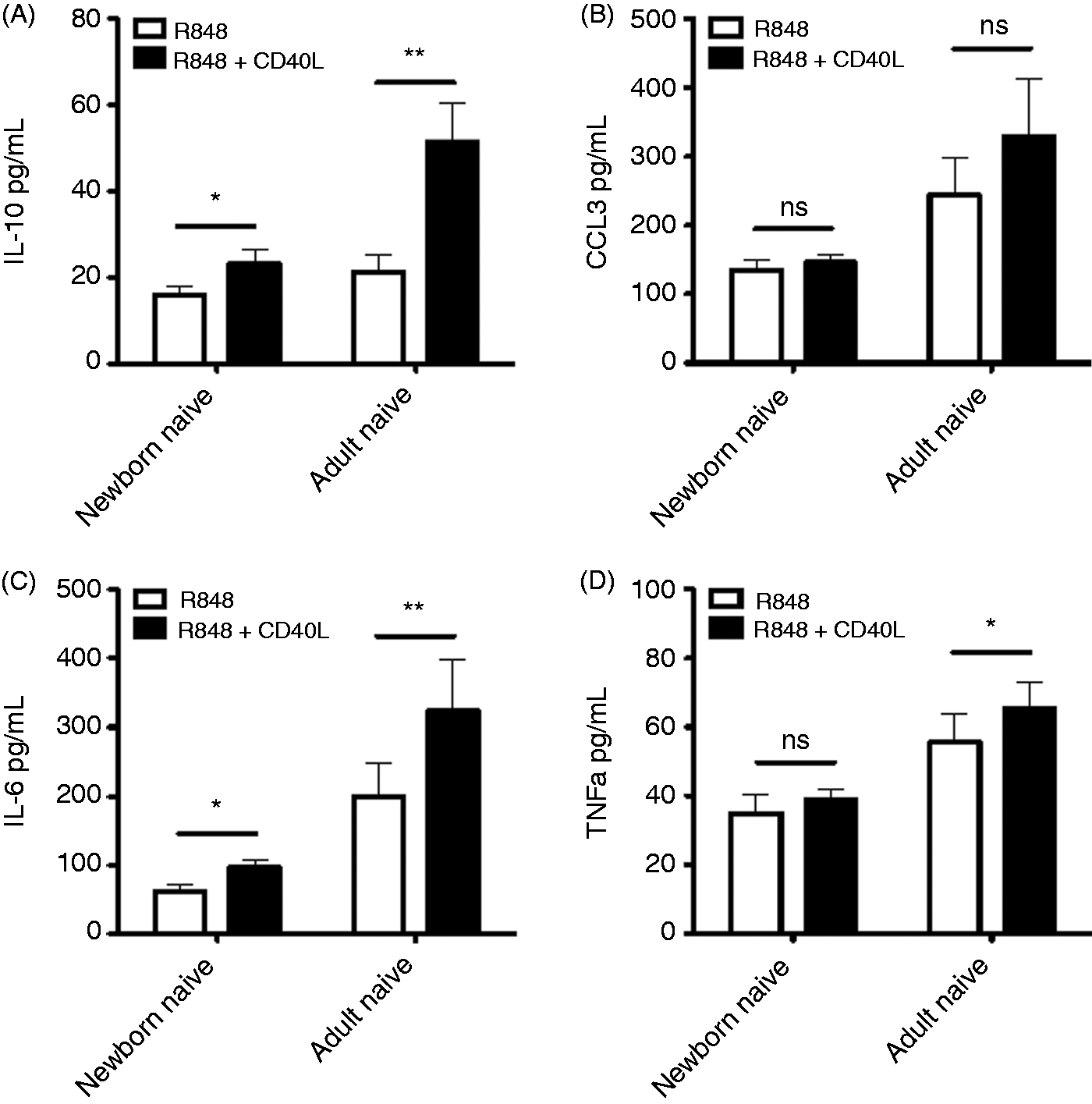
Circulating newborn naïve B cells are impaired in CD40/TLR-mediated CSR
TLR stimulation, in combination with CD40 activation, can lead to B-cell Ig CSR, even in the absence of B-cell receptor (BCR) triggering.20–22 Naïve B cells from both newborn cord blood and adult peripheral blood were stimulated with soluble CD40L and IL-2 (to simulate T-cell help) in conjunction with select TLR agonists in culture for 80 h, after which supernatants were evaluated for cytokine production (by multiplex assay) and cells were re-plated for ELISpot assay to enumerate IgM- and IgG-secreting cells. The profile of cytokine production following stimulation of both CD40 and TLRs for several days in culture was similar to that observed with only TLR stimulation, with the exceptions that stimulation of CD40 in conjunction with TLR9 stimulation promoted IL-10 production in adult naïve B cells to levels similar to those observed in newborn naïve B cells (Figure 4), and that for some of these conditions we detected production of additional cytokines, including IFN-γ, IL-12p40 and IL-12p70 (Supplementary Tables 1 and 2). Data for other cytokines produced under these conditions are shown in Supplementary Tables 1 and 2, including data for both newborn and adult naïve B-cell isolates and adult total CD19+ B-cell isolates. Supplementary Figure 3 provides an overview of the cytokine secretion patterns as a fold-increase over control conditions.
Neonatal naïve B cells demonstrate distinct TLR/CD40-mediated cytokine production. Naïve B cells were stimulated with the following: control condition, IL-2 (30 ng/ml); HKLM condition, IL-2, CD40L (20 ng/ml) and HKLM (109 cells/ml); R848 condition, IL-2, CD40L and R848 (10 µM); CpG condition, IL-2, CD40L and CpG (1 µM). Cells were cultured with stimulation for 80 h before evaluation of supernatant cytokines by multiplex assay. n = 3–5 for HKLM condition; n = 6–8 for control, R848 and CpG conditions. Student’s t-tests, *P < 0.05, **P < 0.01.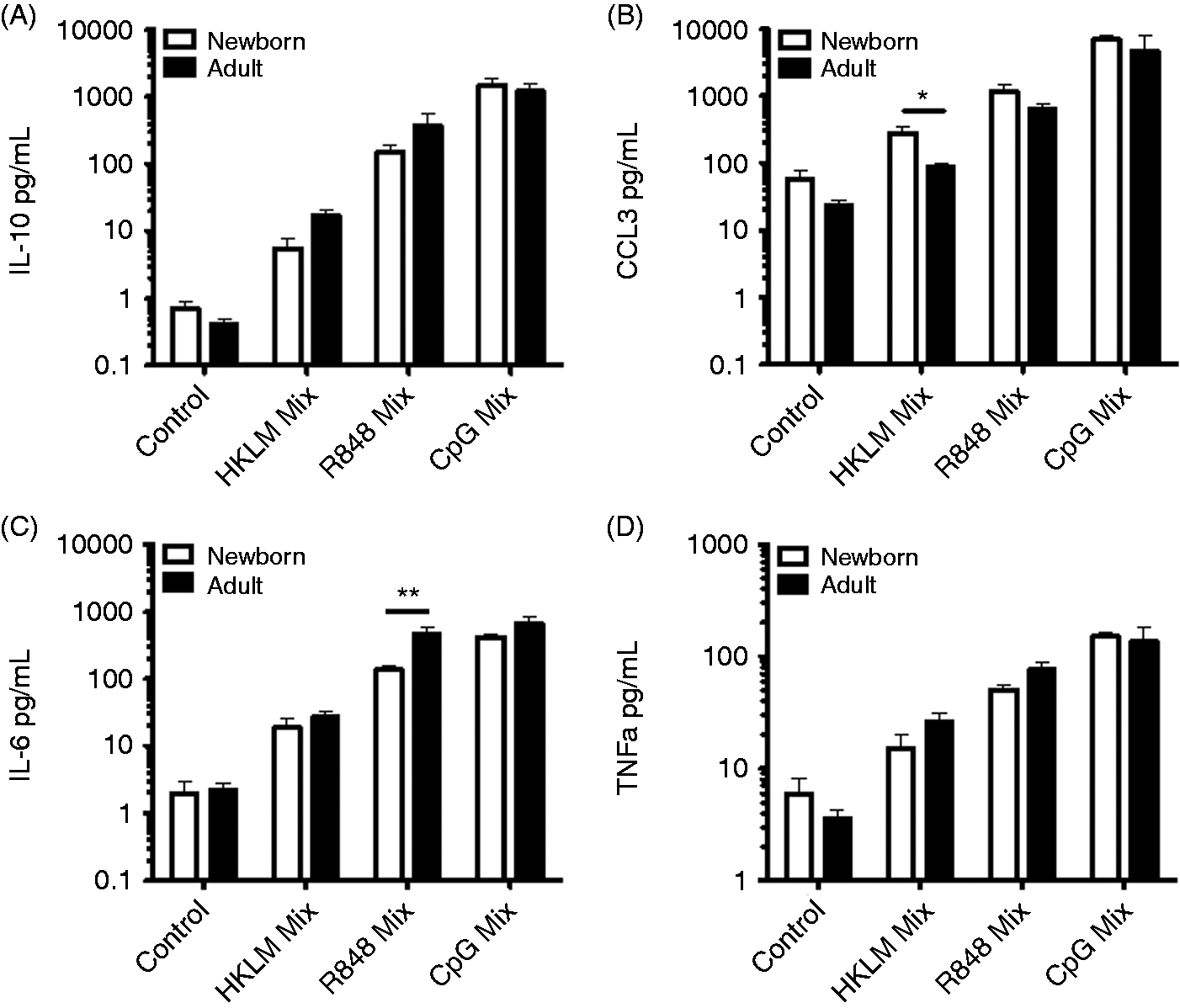
CSR to yield IgG-secreting B cells with dual CD40 and TLR stimulation was significantly impaired in neonatal naïve B cells compared with adult naïve B cells (Figure 5). The percentage of cells secreting IgG was significantly lower for newborn naïve B cells relative to adult counterparts for both TLR7/8/CD40-stimulating and TLR9/CD40-stimulating conditions. Newborn naïve B cells exhibited significantly greater IgG CSR in response to TLR9/CD40 stimulation than for TLR7/8/CD40 stimulation. Dual CD40 and TLR stimulation induced similar numbers of IgM-secreting cells in both populations, with the exception that a higher percentage of newborn naïve B cells secreted IgM in response to TLR2/CD40 stimulation. As expected, adult total CD19+ B cells, which include CD27+ memory B cells, yielded a higher percentage of IgG-secreting cells under all conditions. Because this population was not included for newborn samples, the adult total B-cell data were not compared with other populations in a statistical analysis.
Deficient TLR/CD40-mediated Ab CSR in newborn naïve B cells tested in vitro. Naïve B cells or total B cells (adult) were stimulated with the following; control condition, IL-2 (30 ng/ml); HKLM condition, IL-2, CD40L (20 ng/ml) and HKLM (109 cells/ml); R848 condition, IL-2, CD40L and R848 (10 µM); CpG condition, IL-2, CD40L and CpG (1 µM). Cells were cultured with stimulation for 80 h before cells were transferred to ELISpot assay plates (104 cells per well for IgM, 105 for IgG). (A) IgG and IgM spots from naïve B cells for one assay representative of six, with stimulation conditions as indicated above. (B) Quantification of IgM- (left) and IgG- (right) producing cells. n = 3–5 for adult total B cells, n = 3–5 for naïve B cell HKLM condition, n = 6 for naïve B-cell control, R848 and CpG conditions. Two-tailed, unpaired Student’s t-tests for comparison between populations (adult/newborn), paired tests for different stimuli within population, *P < 0.05, **P < 0.01.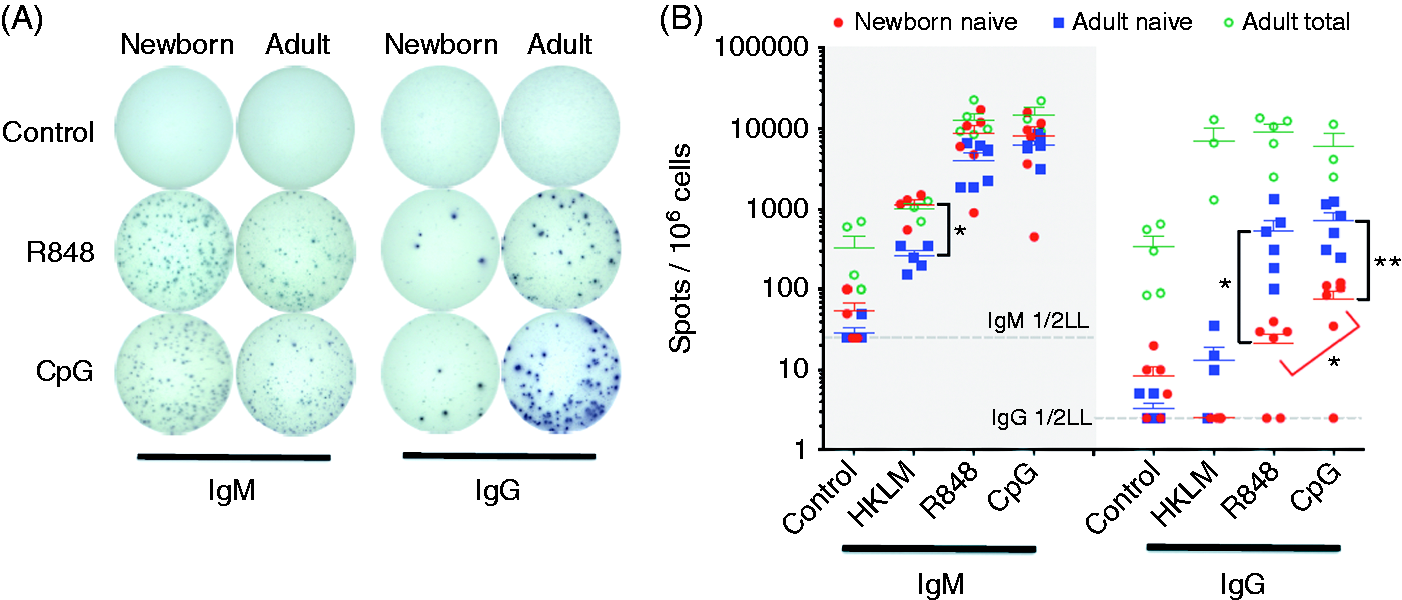
As production of IL-21 by T follicular helper cells can enhance CSR and plasma cell differentiation during T-cell-dependent B-cell responses,23,24 we evaluated the effect of addition of IL-21 in vitro. Addition of recombinant human IL-21 appeared to further enhance CSR in response to TLR9/CD40 or TLR7/8/CD40 stimulation in both newborns and adults (Figure 6). However, even in the presence of IL-21, the percentage of cells secreting IgG was significantly lower for newborn naïve B cells relative to adult counterparts (P < 0.01). Stimulation of newborn naïve B cells with TLR agonists also resulted in fewer up-regulation of activation markers CD80 and CD86, especially in the presence of IL-21 (Figure 6).
Neonatal naïve B cells demonstrate impaired IL-21-induced class switching and expression of co-stimulatory receptors CD80 and CD86. Naïve B cells were stimulated with IL-2 (30 ng/ml) and CD40L (20 ng/ml) alone, or in combination with R848 (10 µM), CpG (1 µM) and/or IL-21 (50 ng/ml). Cells were cultured with stimulation for 80 h before cells were transferred to ELISpot assay plates (104 cells per well for IgM, 105 for IgG). (A) IgG and IgM spots from naïve B cells, with stimulation conditions as indicated above. (B–D) Quantification of surface expression of CD25, CD80 and CD86 by flow cytometry. Fluorescence intensities are shown as MFI fold increase over the control condition. n = 4 for all conditions. two-tailed, unpaired Student’s t-tests for comparison between adults and newborns is indicated. *P < 0.05, **P < 0.01, ***P < 0.001.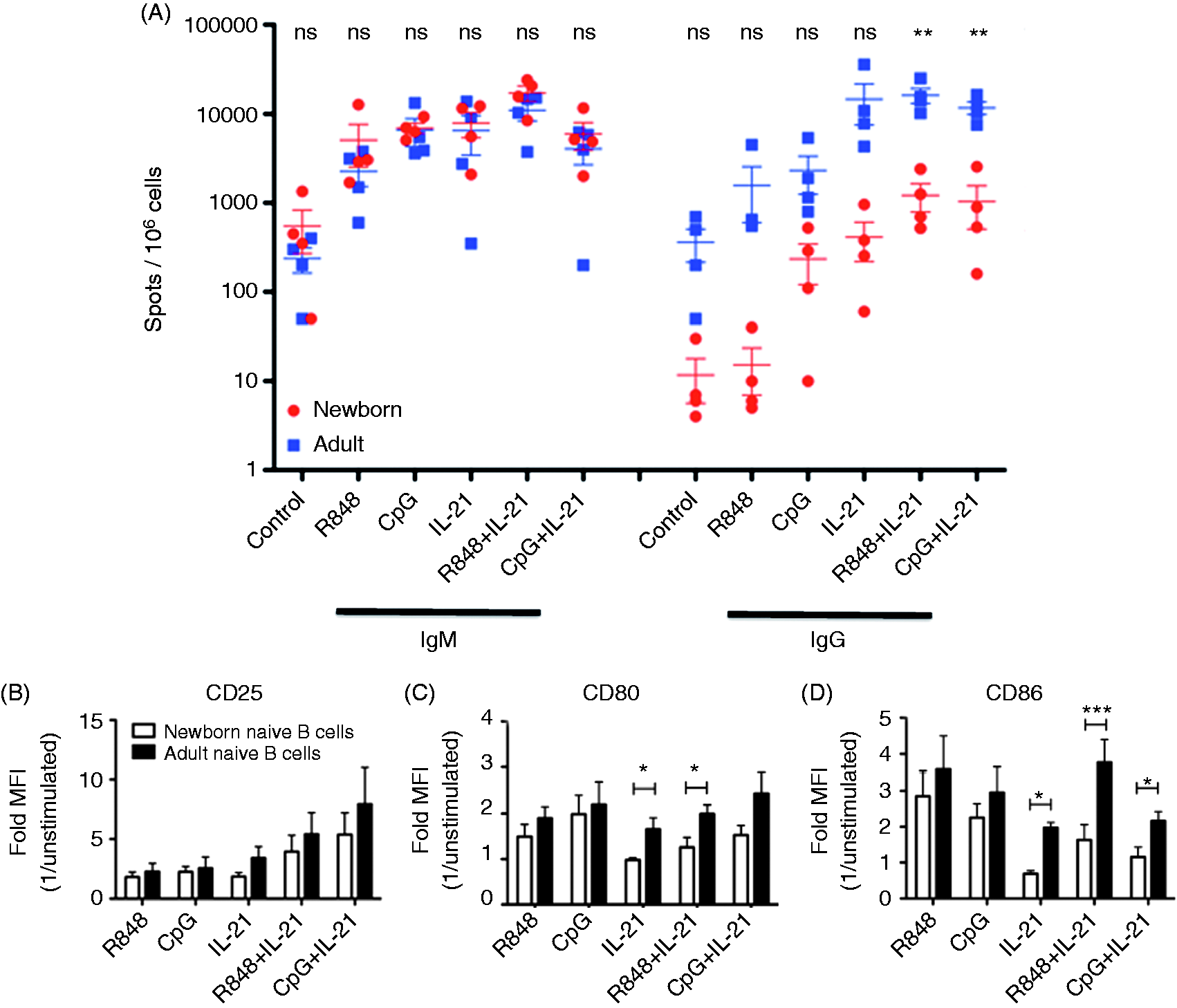
Discussion
Herein we compared, for the first time, the functional expression of TLRs on subsets of circulating CD27− naïve and CD27+ memory B cells of human newborns and adults. Although expression of most TLRs was quite similar for naïve B cells from newborns and adults, TLR9 expression was greater in newborn naïve B cells. Moreover, we noted significant differences with age related to TLR-mediated cytokine induction and the capacity to transition to IgG secretion.
TLRs are critical regulators of B-cell function, and TLR ligation can serve as a co-stimulatory signal in B-cell activation. 25 TLR expression has been evaluated by qRT-PCR of total B cells from human adult peripheral blood isolated by indirect magnetic bead isolation, 26 and by PCR of flow-sorted adult peripheral blood B-cell subsets with qPCR of TLRs 2, 4, 7 and 9. 2 These studies came to distinct conclusions, with the latter finding ‘low to undetectable’ expression of all of the TLRs in naïve B cells. 2 Our study has the advantages of utilizing flow-sorted cells (based on IgD, CD27, and CD38 expression levels) for greater purity and subset analysis, qRT-PCR for quantification of TLR expression on B-cell subsets and a relatively higher number of study participants (n = 9 per age group). While TLR expression on cord blood T cells and monocytes has been characterized, 27 expression of TLRs 1–10 in circulating B cells from cord blood has not previously been evaluated. In this study, we characterized TLR mRNA expression for newborn cord naïve B cells and circulating adult B-cell subsets, including naïve, early memory and class-switched memory B cells. TLRs 6, 7, 9 and 10 were highly expressed on newborn and adult naïve B cells, TLRs 1, 2 and 4 showed moderate expression and very low levels of TLR5 and TLR8 expression were observed. Of note, while heat-killed L. monocytogenes activates via TLR2, live L. monocytogenes expresses a ligand of TLR10, 28 which is one of the mostly highly expressed TLRs on human B cells. At the protein level, newborn naïve B cells display greater TLR4 and TLR9 expression than adults. Adult memory B-cell subsets demonstrated elevated mRNA for TLR7 and elevated mRNA and protein TLR9 expression compared with adult naïve B cells. Elevated TLR9 expression in adult memory B-cell subsets is in agreement with prior studies. 2 Overall, differences in TLR expression likely do not account for all of the distinct responsiveness to TLR stimulation between adult and newborn circulating naïve B cells. Additional features newborn B cells that may contribute to their distinct response to TLR stimulation includes impairment in purine salvage due to diminished levels of CD73. 29
We also evaluated TLR-mediated cytokine production by B cells in this study, both for B cells stimulated with single TLR agonists for 24 h (Supplementary Table 1) and for B cells treated with a combination of TLR agonists, CD40L and IL-2 for 80 h (Supplementary Table 2). Newborn naïve B cells produced significantly greater levels of IL-10 following stimulation with CpG (TLR9) than adult naïve B cells, while producing similar levels following stimulation of TLR2 or TLR4. IL-10 production was enhanced by dual stimulation of TLR7/8 and CD40 in both newborn and adult naïve B cells, although to a greater degree in adult cells than newborn ones. IL-10 production by B cells can suppress inflammation,12,13 and murine B-cell IL-10 production regulates neonatal dendritic cell function, 13 an activity that has earned them the name ‘B regulatory cells’. 30 Of note, naïve adult B cells demonstrate greater IL-10 production in response to CD40 stimulation than memory cells. CCL3 (formerly known as ‘MIP-1α’) is a T-cell chemoattractant that modulates activity of a variety of leukocytes, and is produced by B cells following BCR engagement but not following CD40 stimulation. 31 CCL3 concentrations were moderately up-regulated following TLR7 or TLR9 stimulation in our assay, and were higher in control naïve B-cell supernatants from newborns relative to adults.
TNF-α production by murine B cells plays a critical role in maturation of follicular dendritic cells, 32 and is involved in murine B-cell-mediated up-regulation of IFN-γ from T cells. 33 We detected TNF-α production by human B cells, which varied between newborn and adult populations and in a TLR-specific manner. Neonatal naïve B cells demonstrated less TLR 1/2 (Pam3CSK4)-mediated TNF-α but higher TLR9 (CpG)-mediated TNFα than adult counterparts. IFN-γ, IL-12p40 and IL-12p70 production were detected only at low levels with 24 h of TLR stimulation but were increased after 80 h of culture with stimulation of both TLRs and CD40. Production of IL-12p40 by human B cells is enhanced by CD40 ligation.34,35 The relatively high neonatal B-cell expression of TLR9 coupled with greater TLR9-mediated cytokine production is notable, particularly in relation to studies of newborn mice wherein TLR9 agonists such as CpG markedly enhanced responses to vaccinal antigens such as HBV. 36
We noted a marked impairment in the ability of neonatal B cells to class-switch to IgG in response to combined TLR and CD40 signaling. Newborn naïve B cells demonstrated an equal if not greater ability to secrete IgM but significantly impaired capacity to undergo CSR and secrete IgG. Impurity of CD27+ cells in these assays is unlikely to account for the differences observed, as the level of CD27+ B cells was slightly higher for the newborn B-cell isolates, and did not correlate with the percentage of IgG-secreting cells. Previous reports have demonstrated impaired IgG and IgA secretion in vitro by newborn total B cells relative to adult total B cells following CpG stimulation, 37 or dual pokeweed mitogen/donor CD4 T cell stimulation, 38 although comparisons were among total B-cell fractions, and adult samples would have contained significantly more memory B cells. We detected no differences in CD40 expression between newborn and adult B cells (data not shown), in agreement with other studies. 39 Despite adult-level CD40 expression, neonatal B cells demonstrated diminished CD40 responsiveness, including limited ability of CD40L to enhance newborn B-cell cytokine production, as previously noted. 40 Of note, neonatal T cells express very low levels of CD40L,40,41 suggesting another mechanism whereby capacity to undergo CSR may be impaired in vivo.
Although we provide novel human data with fresh insights into ontogeny and potential translational implications, our study does have limitations. Firstly, as vaccine adjuvants are administered together with a target antigen, further studies will be required to characterize neonatal B-cell responses to candidate adjuvants in the context of vaccinal antigen. Secondly, naive B cells from newborns contain more immature or transitional (CD24hi/CD38hi) B cells than adult naïve B cells, 16 such that differences would be expected when directly comparing newborn and adult naïve B cells. Our approach was selected in order to further characterize neonatal B cells, including functional TLR expression, in the context of prior work,14,15,42 and to model neonatal naïve B-cell responses to TLR agonists that may serve as vaccine adjuvants,43,44 as any of the neonatal naïve B cell subsets may respond, and indeed may affect one another. Indeed, the differences we report, including robust neonatal naïve B-cell functional expression of TLR9, were not predictable from the relative proportion of immature B cells.
While the development of new vaccine adjuvants is a growing field, 45 less attention has been given to human B-cell responses to adjuvants. Of note, the magnitude and persistence of Ab responses against pandemic H1N1 influenza in Rhesus macaques may involve direct triggering of TLRs on B cells and dendritic cells, as well as on T-cell help. 46 Additionally, components of subunit vaccines, such as pneumococcal conjugate vaccine polysaccharides, 47 while directly interacting with B cells, may lack the adjuvant activity to induce and optimally shape an immune response in early or later life. Indeed, most infant vaccines require multiple doses to achieve protective Ab titers. Accordingly, increased appreciation of B-cell immune ontogeny may inform development of rationally designed age-specific vaccine formulations, which may include adjuvants that more effectively enhance B-cell responses in early life, potentially providing dose-sparing effects or possibly even single shot protection. TLR9 stimulation enhances humoral immune responses in both neonatal and adult mice,36,48,49 as well as in human clinical trials.50,51 TLR7/8 agonists may also be effective at enhancing Ab responses indirectly through activation of other APCs that subsequently influence B-cell activation. 46 To the extent that our in vitro studies of human newborn B cells reflect their potential responses in vivo, our current work suggests that TLR9 agonists may be particularly effective in activating human newborn and adult B cells for vaccination strategies wherein enhanced Ab production is desirable.
Supplemental Material
Supplemental material for Distinct TLR-mediated cytokine production and immunoglobulin secretion in human newborn naïve B cells
Supplemental Material for Distinct TLR-mediated cytokine production and immunoglobulin secretion in human newborn naïve B cells by Matthew A Pettengill, Simon D van Haren, Ning Li, David J Dowling, Ilana Bergelson, Jop Jans, Gerben Ferwerda and Ofer Levy in Innate Immunity
Footnotes
Acknowledgments
We thank the Labor and Delivery staff at The Brigham & Women’s Hospital, Boston, MA and Beth Israel Deaconess Medical Center Boston, MA, for their kind assistance with sample acquisition. Drs Michael Wessels and Gary Fleisher are thanked for their support of the Precision Vaccines Program.
Declaration of Conflicting Interests
The author(s) declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: OL’s laboratory has received sponsored research support from Crucell (Johnson & Johnson), MedImmune and 3 M Drug Delivery Systems, companies that develop adjuvants and/or vaccines.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: OL’s laboratory is supported by an internal Boston Children’s Hospital award to the Precision Vaccines Program, Global Health (OPPGH5284) and Grand Challenges Explorations (OPP1035192) awards from the Bill & Melinda Gates Foundation and by National Institutes of Health (NIH) grants 1R01AI100135-01, 3R01AI067353- 05S1 and U01AI124284-01, as well as National Institute of Allergy & Infectious Diseases Adjuvant Discovery Program, Contract No. HHSN272201400052C. MP was supported by NIH Training Grant T32 HD055148. SvH was supported by an Early career Award from the Thrasher Research Fund.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.