
Abstract
Indoleamine 2,3-dioxygenase (IDO) is a rate-limiting enzyme for the degradation of tryptophan (Trp) along the kynurenine (Kyn) pathway, and its increased activation is associated with immunologic disorders. Because the specific role of IDO activation is not yet completely clear, the aim of the present study was to establish a pig model of IDO activation for further research. The activation of IDO in pigs was induced experimentally by LPS stimulation in vivo and ex vivo. IDO activation was characterized by measuring Trp, Trp metabolites and IDO protein expression in blood, liver, lung, muscle and different brain areas. The results show that the in vivo LPS administration induced increased plasma concentrations of TNF-α and IL-10, a depletion of Trp and an increase of Kyn, indicating an elevated enzymatic activity of IDO. This was supported by an LPS-induced IDO protein expression in blood, liver and lung. The ex vivo LPS stimulation also resulted in increased TNF-α concentrations and an IDO activation, characterized by an increase of Trp metabolites and IDO protein expression. In conclusion, our data emphasize that the LPS stimulation is a suitable model for IDO activation in the domestic pig, which provides a basis for further research on immunoregulatory IDO functions.
Introduction
Indoleamine 2,3-dioxygenase (IDO) is a rate-limiting enzyme for the degradation of tryptophan (Trp) along the kynurenine pathway to biologically active metabolites such as kynurenine (Kyn), kynurenic acid (Kyna) or quinolinic acid (Quin). This pathway of Trp metabolism is a link between the immune and the nervous system, and plays a crucial role in the regulation of the immune response. The IDO activation is one mechanism of innate immunity and has different physiological functions with a highly cell-type specific pattern of inducibility. IDO is expressed ubiquitously in a constitutive or inducible manner. 1 In many cell types it is induced by specific inflammatory stimuli, such as IFN-γ or LPS. 2 Moreover, LPS may also activate IDO by the production of TNF-α. 3 The depletion of Trp by IDO induction leads to suppression of intracellular pathogens and T-cell proliferation,4,5 and is associated with the development of depressive-like behavior in mice.6,7 An increased activity of IDO was also measured in a number of immunologic and neurologic disorders. For example, it was found that IDO is highly expressed in malignant human tumors leading to an effective immune escape by inhibiting T-cell responses, 8 and, in addition, an increased IDO activity in patients with sepsis is associated with a higher mortality risk. 9 An accumulation of Kyna or Quin was found in several neurologic disorders, for example Alzheimer’s disease, 10 schizophrenia 11 and Huntington disease, 12 suggesting neuroactive functions of these Trp metabolites. However, the immunosuppressive or neuroactive function of IDO activation in these disorders is not yet fully understood, and an animal model with high similarity of its Trp metabolism to that of humans would be necessary for further studies in this field. The pig is a species that closely resembles humans in anatomy, genetics and physiology, and is used increasingly as a model for human in biomedical research. Compared with rodents, pigs have obvious advantages with respect to the relevance for human pathophysiology.13,14 The size of pigs allows the use of instrumentation and monitoring devices similar to those used in humans, and the amount of blood allows for repeated blood sampling and comprehensive analyses. 15 Thus, relating to the current research in the therapeutic effect of IDO inhibition, experiments in pigs are much more likely to be predictive of therapeutic treatments in humans than experiments in rodents. Therefore, the aim of this study was to establish a pig model of IDO activation.
In the present study, we investigated IDO activation in pigs in vivo and ex vivo. The activation of IDO in vivo was induced by a single i.p. LPS administration. The inflammatory immune response to LPS was characterized by the pro-inflammatory cytokine TNF-α and the anti-inflammatory cytokine IL-10. IDO activity was determined by measuring Trp and Kyn in plasma. For the first time in pigs we measured the downstream metabolites Kyna and Quin as markers for the activation of the Kyn pathway after LPS application, and the LPS-induced IDO protein expression in blood and tissues. In addition to IDO activation in vivo, we investigated IDO induction ex vivo by the stimulation of porcine blood with LPS in a whole blood culture system.
Materials and methods
Animals
A total of 50 castrated male pigs from German Landrace, bred and raised in the experimental pig unit of the Leibniz Institute for Farm Animal Biology, were used in two experiments. All pigs received standard processing (oral iron supplementation and castration) within the first 3 d of life. At the beginning of the experiments, pigs were 7 wks old and weighed between 12 and 18 kg. Their health was checked continuously by visual inspection during the wk previous to the experiments. All the procedures involving animal handling and treatment were in strict accordance with the German animal protection law and approved by the relevant authorities (Landesamt für Landwirtschaft, Lebensmittelsicherheit und Fischerei, Mecklenburg-Vorpommern, Germany; LALLF M-V/TSD/7221.3-2.1-005/08, LALLF M-V/TSD/7221.3-2.2-014/09, LALLF M-V/TSD/7221.3-2.2-003/10).
Experimental design
Experiment 1: in vivo LPS administration
One wk before LPS administration, the animals were kept individually. To avoid stress, weighing of the pigs was carried out 3 d before starting the experiments. For the time of LPS and saline administration an approximated body mass (BM) was calculated. Each pen was equipped with a feeder and a nipple waterer. Pigs were fed ad libitum with a commercial diet and had free access to water. On the day of the LPS administration, feed was removed 1 h before treatment to avoid an effect of feed uptake on plasma Trp concentrations. At 8:00 a.m., 20 pigs received a single i.p. LPS administration of 100 µg LPS per kg BM (Escherichia coli O111:B4; Sigma-Aldrich, Deisenhofen, Germany), according to Tuchscherer et al., 16 and dissolved in 3 ml sterile endotoxin-free 0.9% NaCl (physiological saline). A control group of 20 pigs was treated with an equivalent volume (3 ml) of sterile, endotoxin-free saline solution. In preliminary experiments, different doses of LPS (1, 5, 25, 100 µg/kg BM) were tested to determine an appropriate dose in pigs for ensuring reproducible systemic inflammatory responses and alterations in Trp metabolism. Furthermore, the type of application was examined, and the i.p. administration of LPS was chosen because this procedure is less stressful and more practicable in younger pigs compared with i.v. injection. Six h after the i.p. treatment, the animals received 1 kg feed. For analysis of cytokines, Trp metabolites and IDO protein expression, blood samples were taken while piglets were in a supine position by anterior vena cava puncture before treatment (0 h), and 1, 3, 6 and 24 h after LPS administration. The whole procedure lasted <1 min. For collecting tissues, 20 pigs were euthanized by an injection of T61 (Intervet, Unterschleißheim, Germany) 6 h after LPS (n = 10) or saline (n = 10) treatment. The remaining pigs were euthanized 24 h after LPS (n = 10) or saline (n = 10) treatment. The time points for tissue collecting resulted from preliminary experiments concerning the temporal dynamic of IDO activity which revealed the highest Kyn/Trp ratio 6 h after LPS, and alterations in Trp and its metabolites 24 h after LPS. The number of pigs was determined by statistical experimental design to detect differences between the treatment levels (and/or time levels) of at least one residual SD assuming a first kind risk α = 0.05 and that the second kind risk β is not larger than β0 = 0.20.
Experiment 2: ex vivo LPS stimulation in whole blood culture system
Seven-wk-old pigs were kept in group housing with free access to feed and water. Blood was taken as described above at 8:00 a.m. from 10 healthy untreated pigs. The cultivation of porcine blood in whole blood culture system was used for ex vivo LPS stimulation, described previously by Tuchscherer et al.
17
Briefly, heparin-anticoagulated whole blood samples were diluted 1:5 with RPMI-1640 containing 10% FCS, 50 µg/ml gentamycin, 2 mM
Blood and tissue sampling
Blood samples were collected in ice-cooled tubes containing EDTA, subsequently centrifuged at 2000 g for 10 min at 4℃. Then, the blood plasma was stored at −80℃ until analysis. The liver, lung, muscle and brain tissues were removed quickly and amygdala, hippocampus, hypothalamus, pituitary and prefrontal cortex (PFC) were dissected, frozen in liquid nitrogen and stored at −80℃ until analyses. The stereotaxic atlas of the pig brain served as a reference. 18
TNF-α and IL-10 assays
Concentrations of TNF-α and IL-10 were analyzed in blood plasma and brain tissues using a commercially available pig ELISA kit (Invitrogen, Frederick, MD, USA). Analyses were performed according to the manufacturer’s instructions. The sensitivities of the TNF-α and IL-10 assays were 3 pg/ml. The intra- and inter-assay coefficients of variation (CV) of TNF-α were 6.2% and 8.2% respectively. The intra- and inter-assay CVs of IL-10 were 6.3% and 9.4% respectively. For cytokine analysis in hippocampus and PFC, samples were homogenized in ice-cold buffer containing a protease inhibitor cocktail and incubated at 4℃ for 60 min. Homogenate was centrifuged at 20,000 g for 20 min at 4℃, and TNF-α and IL-10 were analyzed in the supernatants. Protein content was determined by the procedure of Lowry et al. 19
Quantification of plasma Trp and metabolites of the Kyn pathway
The method used in this study was described recently by Kiank et al. 20 EDTA plasma (100 µl) was mixed with 50 µl trichloroacetic acid (10 g in 10 ml water), 40 µl water and 10 µl internal standard solutions [d5-phenylalanine (Phe), d4-Kyna, d6-Kyn; Eurisotop, Paris, France], incubated at 4℃ for 12 h (overnight) and centrifuged at 25,000 g for 15 min at 4℃. The supernatant was collected and levels of Trp and its derivates were measured using a Wallac MS2 tandem mass spectrometer (Perkin Elmer, Rodgau, Germany). It was equipped with an electrospray ion source and the configuration allowed the analysis of Trp, d5-Phe, d6-Kyn, Kyn, d4-Kyna, Kyna and Quin. Ions were detected in a positive ion mode using multiple reaction monitoring. Nitrogen served as the collision gas. Flow solvent (0.02% formic acid in 50% aqueous acetonitrile) was delivered at a flow rate of 50 µl/min with an injection volume of 20 µl. Calibration curves were fitted by linear least-square regression and correlated to the concentration of d5-Phe, d6-Kyn and d4-Kyna as internal standards. As an indicator for IDO activation, the ratio of Kyn and Trp was calculated (Kyn/Trp ratio).
Western blot analysis
IDO protein expression was analyzed in plasma, supernatants of whole blood cultures and in several tissues. Tissues were homogenized with radioimmunoprecipitation assay buffer (50 mM Tris-HCl, 150 mM NaCl, 0.5% sodium deoxycholate, 10 mM EDTA, 1% SDS; pH 7.5) including a protease inhibitor cocktail (Sigma Aldrich). After ultracentrifugation at 40,000 g for 60 min at 4℃ (SORVALL® Discovery 90SE; KI Scientific, Lane Cove, Australia) the supernatants were analyzed. The protein concentration in blood plasma and supernatants was quantified using the Biuret reaction. 21 Proteins were separated by 10% SDS-page 22 followed by transfer onto a polyvinylidene fluoride membrane (Millipore, Bedford, MA, USA). After blocking of this membrane for 1 h at 20–22℃ (room temperature) with Tris-buffered saline and Tween 20 (TBST; 20 mM Tris, 130 mM NaCl, 0.1% Tween 20; pH 7.5), containing 5% skimmed milk powder, it was washed three times with TBST. After this, the blot was incubated with TBST + 5% milk powder containing a rabbit anti-hIDO polyclonal Ab (1:2000; AB9898; Millipore) overnight at 4℃. The membrane was washed three times with TBST followed by the incubation with HRP-conjugated anti-rabbit IgG (1:5000; sc-2004; Santa Cruz Biotechnology, Santa Cruz, CA, USA) for 1.5 h at room temperature. Finally, the membrane was washed three times with TBST. The protein signal was detected by enhanced chemiluminescence using ECL-Plus (GE Healthcare Bio-Science AB, Uppsala, Sweden). As a positive control for IDO protein, the IDO(h): 293T Lysate (sc-114136; Santa Cruz Biotechnology) was used. For loading control of tissue samples, the blots were re-probed using a HRP-linked β-actin Ab (1:3000; Cell Signaling Technologies, Frankfurt, Germany). To determine the molecular mass of the target protein we used ECL DualVue Western blotting marker (GE Healthcare Bio-Science AB, Uppsala, Sweden).
Statistical analyses
Statistical analyses were performed using SAS software, version 9.2 (SAS, Cary, NC, USA). The data were evaluated by ANOVA using the mixed procedure. For the data from experiment 1, the model comprised the fixed effects treatment (LPS, saline), time (0, 1, 3, 6, 24 h), replication (2), their two-way interactions and the random effect sow. Repeated measures on the same animal were taken into account using the repeated statement in the MIXED procedure and a compound symmetry block diagonal structure of the residual covariance matrix. For the evaluation of the data from experiment 2, the differences between LPS-stimulated and unstimulated control samples (Δ) of TNF-α, Trp, Kyn, Kyna, Quin concentrations and Kyn/Trp were calculated. The model for the resulting difference variable ΔTNF-α, ΔTrp, ΔKyn, ΔKyna, ΔQuin and ΔKyn/Trp ratio comprised the fixed effect cultivation time (4, 6, 24 h) and the random sow effect. Least-square means (LS-means) and their standard errors (SE) were calculated and tested for each fixed effect in the models described earlier using the Tukey–Kramer procedure for all pair-wise multiple comparisons. Differences were considered significant if P < 0.05.
Results
Experiment 1: IDO activation by in vivo LPS administration
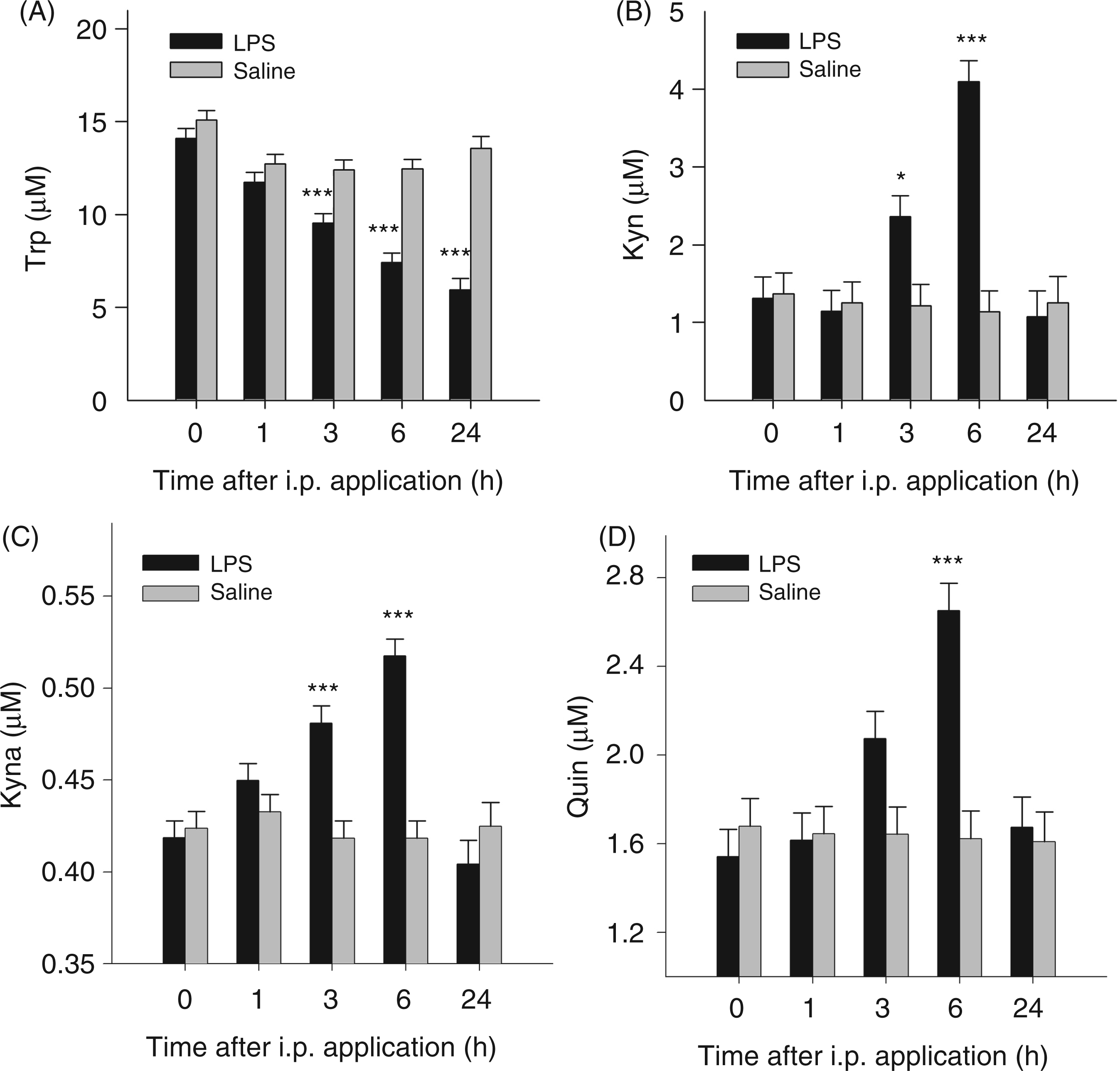
ANOVA revealed a significant effect of the interaction treatment × time on plasma concentrations of TNF-α, IL-10, Trp, Kyn, Kyn/Trp ratio, Kyna and Quin (for all parameters P < 0.001). Plasma concentrations of the cytokines TNF-α (Figure 1A) and IL-10 (Figure 1B) increased significantly 1 h after LPS application compared with the saline-treated group (control), but returned to baseline by 3 h after LPS. Trp concentrations were reduced significantly 3, 6 and 24 h after LPS administration compared with the control group (Figure 2A). The concentrations of Kyn and Kyna were elevated significantly 3 and 6 h after LPS (Figure 2B, C), and the plasma concentration of Quin was increased 6 h after LPS administration compared with the control group (Figure 2D). The Kyn/Trp ratio was increased significantly 3 h after LPS and reached a maximum after 6 h (Figure 3A).
Plasma concentrations of TNF-α (A) and IL-10 (B) after i.p. LPS or saline administration. Cytokines were measured at different time points in blood plasma by ELISA. Results are presented as LS-means + SE. Significant differences between LPS- and saline-treated groups are shown for each time point. For time points 0, 1, 3 and 6 h, n = 20 per group; for time point 24 h, n = 10 per group; ***P < 0.001. Tryptophan metabolites in blood plasma after i.p. LPS or saline administration. Trp (A), Kyn (B), Kyna (C) and Quin (D) were measured at different time points by tandem mass spectrometry. Results are presented as LS-means + SE and significant differences between LPS- and saline-treated groups are shown for each time point. For time points 0, 1, 3 and 6 h, n = 20 per group; for time point 24 h, n = 10 per group; *P < 0.05, ***P < 0.001. Kyn/Trp ratio (A) in plasma after i.p. LPS or saline adminstration. Kyn/Trp ratio is presented as LS-means + SE and significant differences between LPS- and saline-treated groups are shown within each time point. For time points 0, 1, 3 and 6 h, n = 20 per group; for time point 24 h, n = 10 per group; *P < 0.05, ***P < 0.001. IDO protein expression in plasma (B) was analyzed by Western blot analysis. Protein (50 µg) was added into each lane. IDO(h): 293T lysate was used as control. One representative result of three is shown.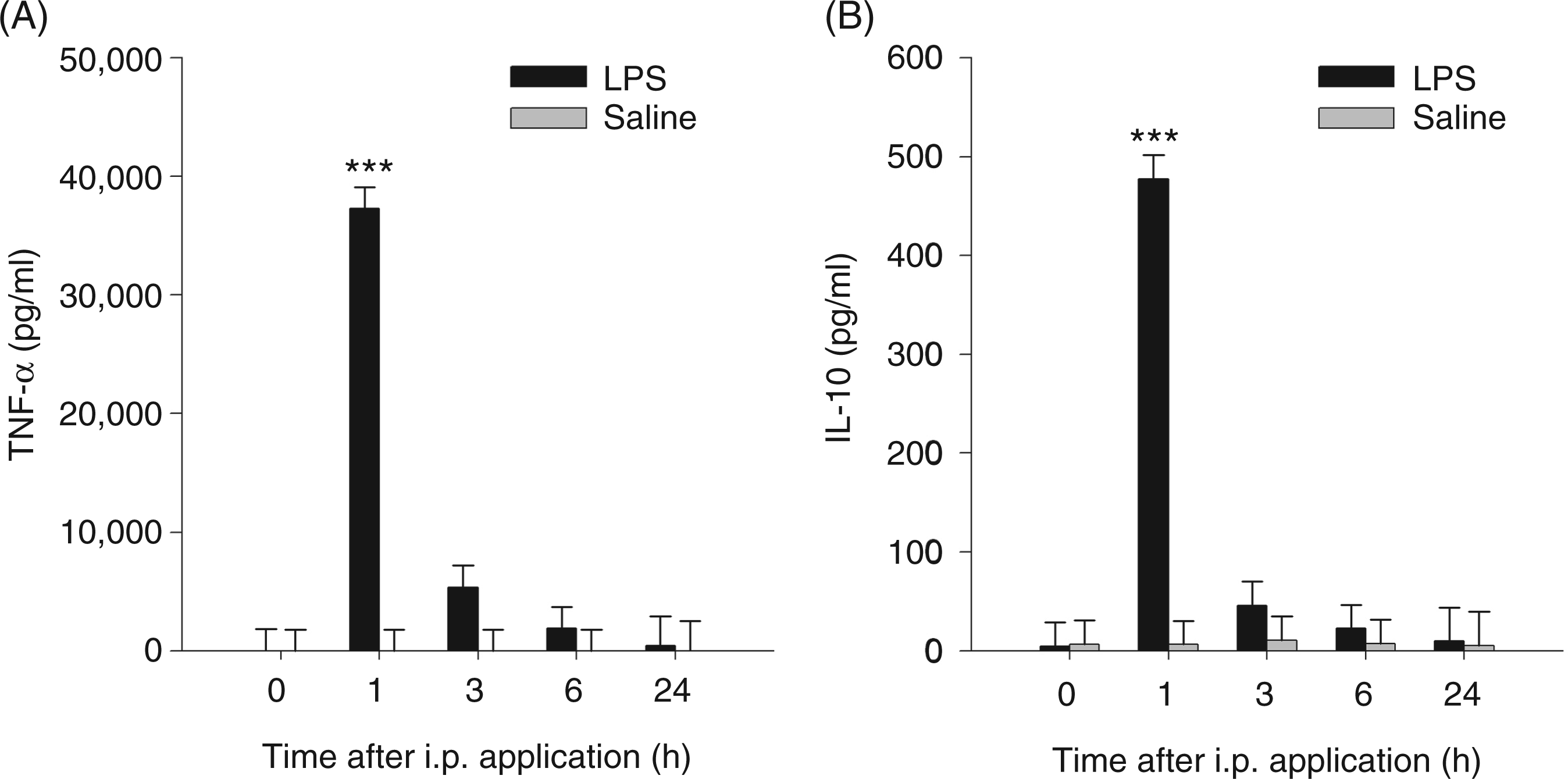
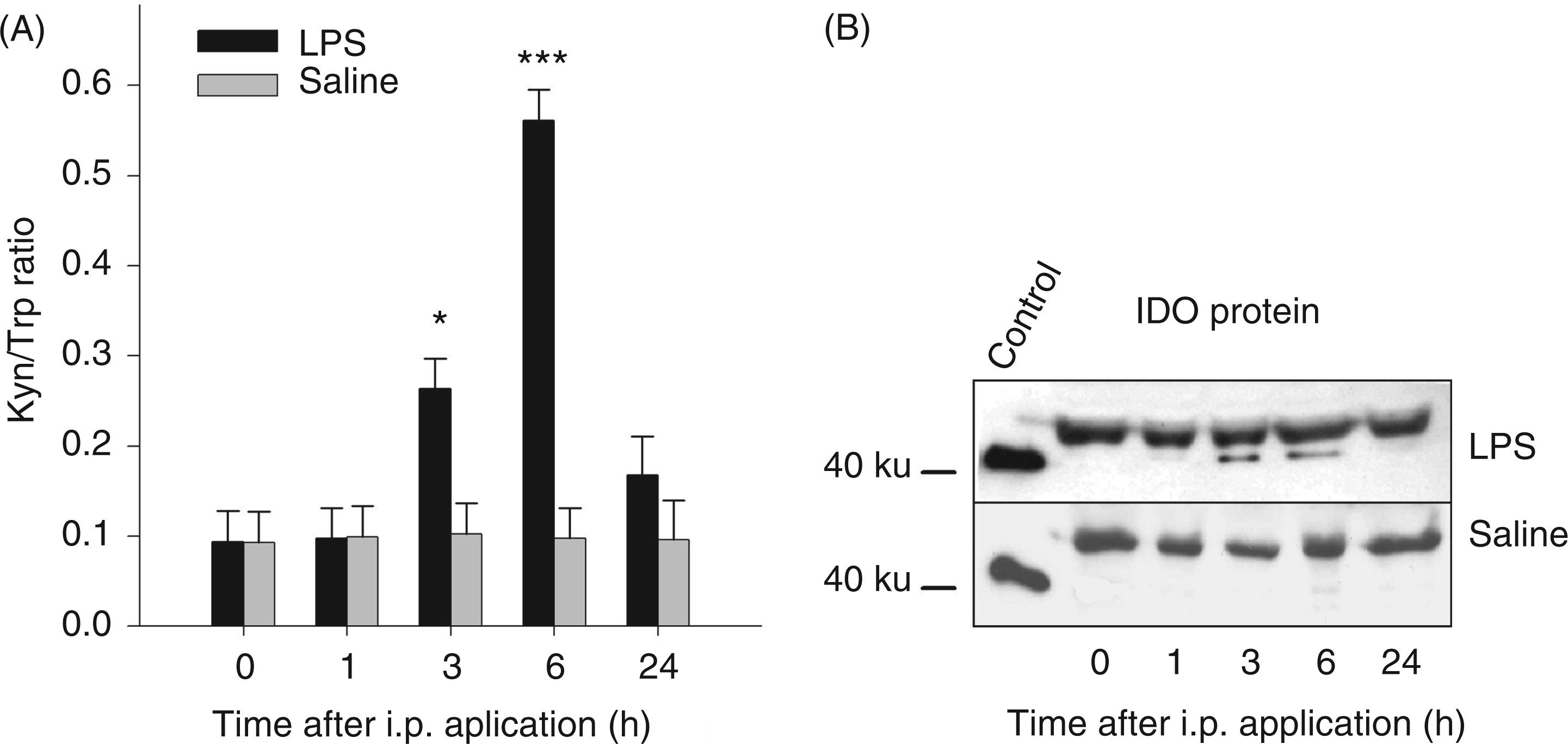
Western blot analyses showed that LPS affected IDO protein expression in blood plasma. One constitutive protein form with a molecular mass of approximately 50 ku was found in the plasma of LPS- and saline-treated groups at all times. A variant with ∼42 ku was induced 3 and 6 h after LPS administration (Figure 3B).
The IDO protein expression in peripheral and brain tissues was analyzed 6 h after LPS application. Western blot analyses of tissue supernatants showed LPS-induced IDO protein expression in the liver and lung, whereas IDO expression in muscle was not affected by the treatment (Figure 4A). In the liver, we found two forms of IDO protein. One variant with a molecular mass of ∼37 ku was detected in both LPS- and saline-treated groups, whereas a second form with a molecular mass of ∼34 ku was only detected in LPS-treated pigs. In the lung, LPS-treated pigs displayed one form of IDO protein with a molecular mass of approximately 34 ku, whereas no protein expression was found in saline-treated pigs (Figure 4A). Analysis of protein expression in different areas of the brain (amygdala, hippocampus, hypothalamus, pituitary and PFC) revealed no detectable IDO protein expression 6 h after LPS (Figure 4B). There were also no changes in concentrations of TNF-α and IL-10 in hippocampus and PFC 6 and 24 h after LPS injection (Table 1).
IDO protein expression in tissues after i.p. LPS or saline (sal) administration. Tissue samples were taken 6 h after treatment. IDO protein expression was detected by Western blot analysis in liver, lung and muscle (A), and in the brain areas amygdala (amy), hippocampus (hipp), hypothalamus (hyp), pituitary (pit) and PFC (B) 6 h after LPS. Protein (20 µg) was added to each lane; for loading control, β-actin was used. IDO (human): 293T lysate was used as a positive control. One representative experiment of three is shown. Concentrations of TNF-α and IL-10 in hippocampus and PFC after in vivo LPS or saline administration. Data are expressed as LS-means ± SE. n = 10 per treatment (6 h), n = 10 per treatment (24 h).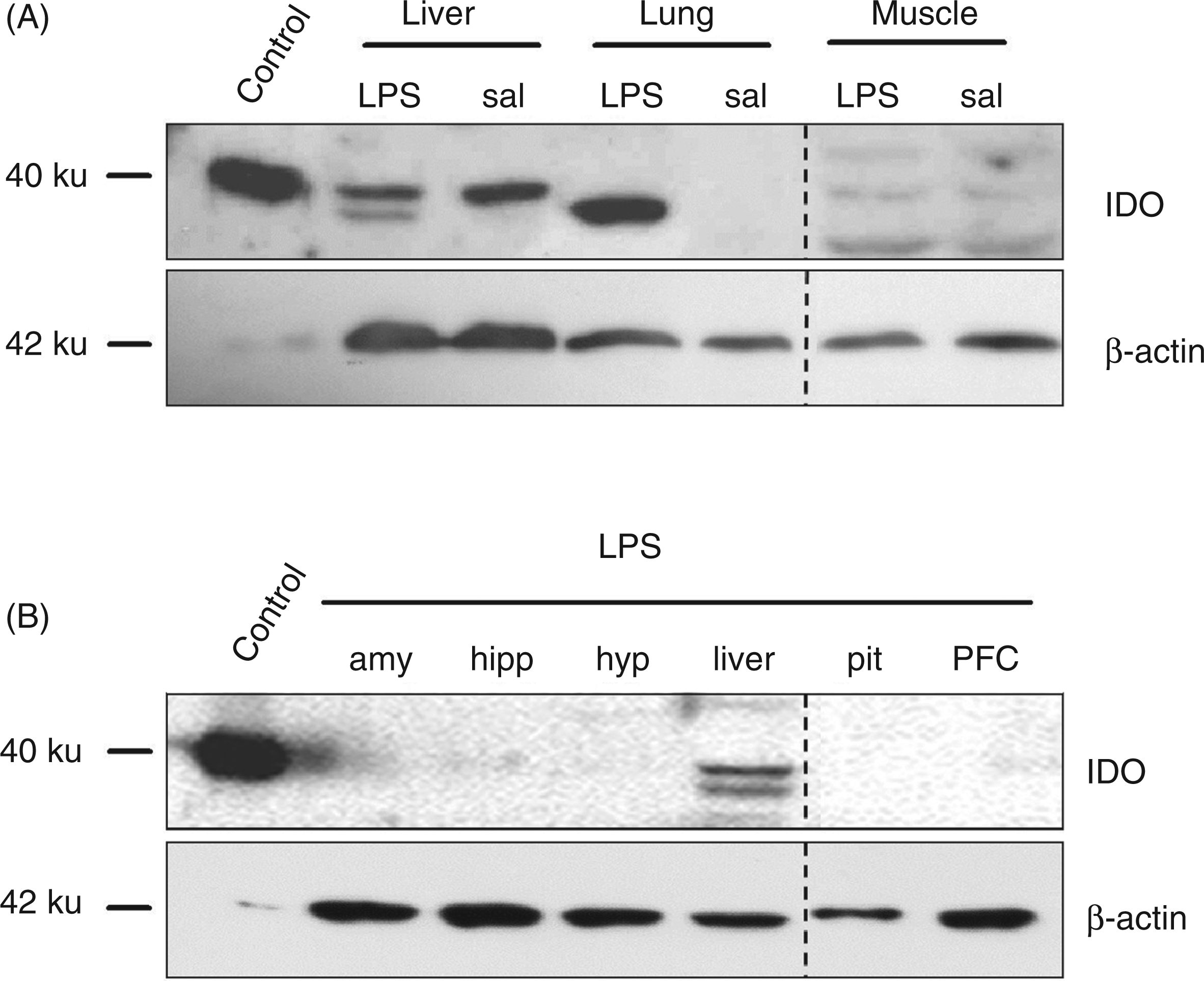
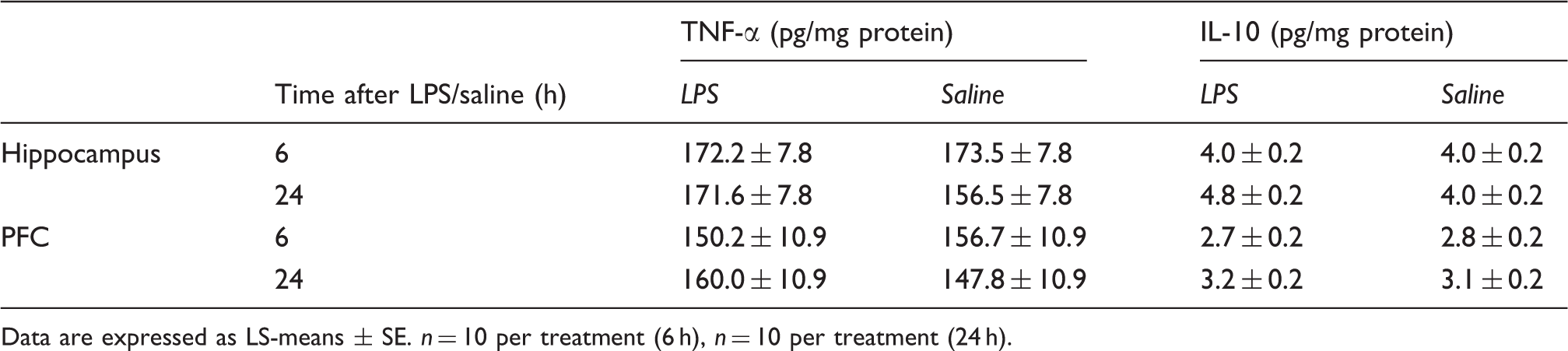
Experiment 2: IDO activation by ex vivo LPS stimulation
ANOVA revealed a significant effect of cultivation time on ΔTrp, ΔKyn, ΔKyn/Trp ratio and ΔQuin (for all parameters P < 0.001), but not on ΔTNF-α. Tukey–Kramer tests showed that the TNF-α concentrations were increased significantly by LPS stimulation of 4, 6 and 24 h compared with unstimulated blood samples (P < 0.001; data not shown). ΔTrp was increased after 6 h of stimulation compared with 4 and 24 h. After 24 h of LPS stimulation, ΔTrp was decreased compared with 4 and 6 h (Figure 5A). ΔKyn was higher after 24 h of LPS stimulation compared with stimulation for 4 and 6 h (Figure 5B). There was no detectable concentration of Kyna in the whole blood cultures (Figure 5C), whereas ΔQuin increased with the cultivation time from 4 to 24 h (Figure 5D). The ΔKyn/Trp ratio was increased significantly in culture supernatants after 24 h LPS stimulation (Figure 6A). In addition, Western blot analysis showed a different protein expression between stimulated and unstimulated blood cells, cultured for 24 h, with a second protein form (∼42 ku) after LPS stimulation (Figure 6B). A constitutive protein form with approximately 50 ku was found in both stimulated and unstimulated samples at all investigated cultivation times.
Concentrations of tryptophan metabolites in whole blood cultures. Blood was cultivated for 4, 6 or 24 h with or without LPS. The metabolites Trp (A), Kyn (B), Kyna (C) and Quin (D) were measured in supernatant by tandem mass spectrometry. Results are presented as LS-means ± SE of differences (Δ) between LPS-stimulated and unstimulated control blood samples: n = 10. *P < 0.05, **P < 0.01, ***P < 0.001. Kyn/Trp ratio (A) and IDO protein expression (B) in whole blood cultures. Kyn/Trp ratio is presented as LS-means ± SE of differences (Δ) between LPS-stimulated and unstimulated control blood samples. Proteins were detected by Western blot analysis. Protein (30 µg) was added to each lane. One representative experiment of three is shown. *** P < 0.001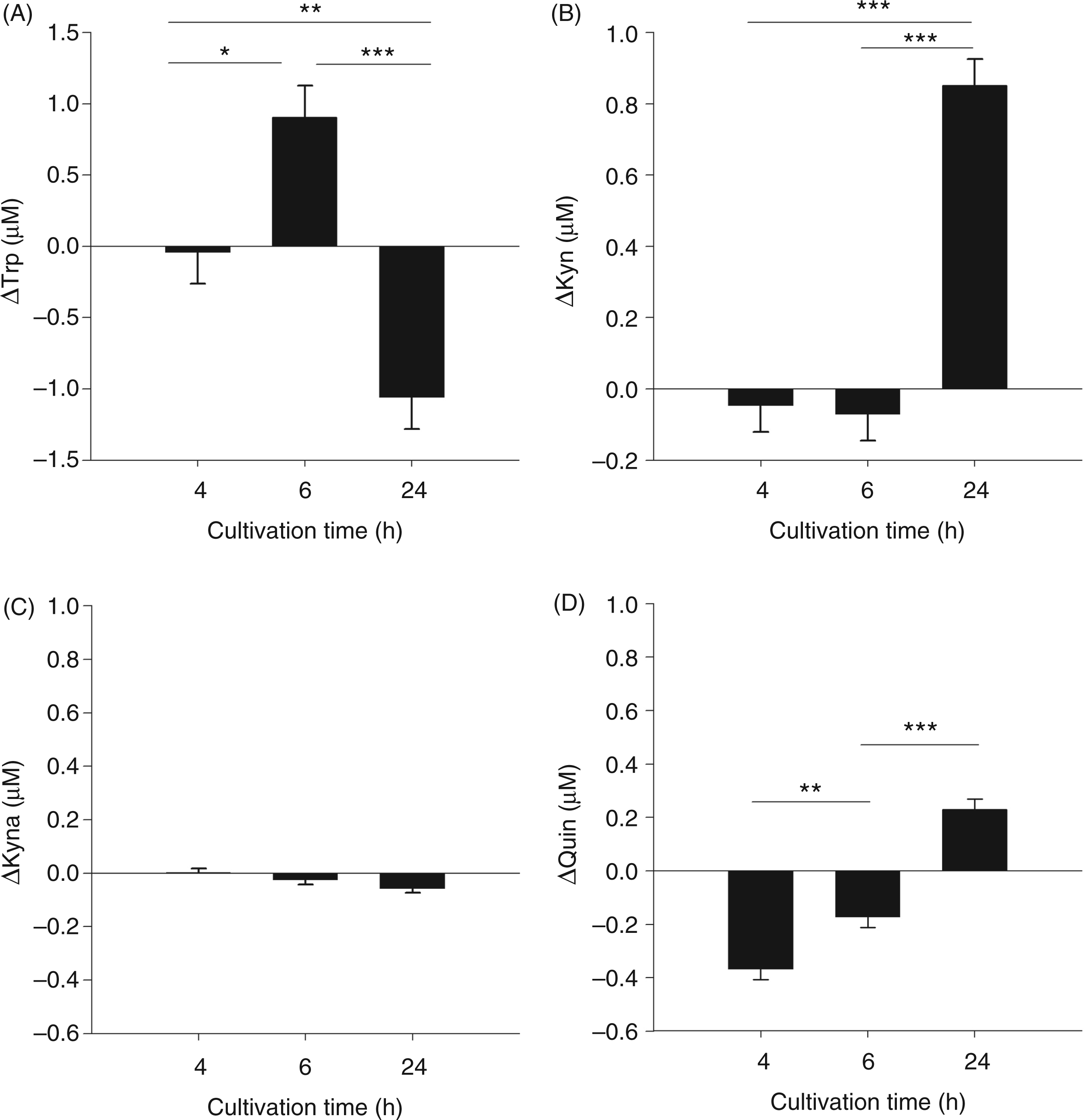
Discussion
The present study investigates the in vivo and ex vivo activation of IDO by LPS in pigs. The results show that the LPS administration in vivo induced increased plasma concentrations of pro- and anti-inflammtory cytokines, indicating activation of the inflammatory immune response. The depletion of Trp and the accumulation of Kyn in plasma and the increased plasma concentrations of the metabolites Kyna and Quin refer to an activation of the Kyn pathway of Trp degradation by LPS administration. These results are supported by Western blot analyses, detecting an LPS-induced IDO protein expression in plasma and tissues.
The ex vivo studies showed that LPS stimulation induced an increase of TNF-α, Kyn and Quin concentrations, and a decrease of Trp concentrations in the supernatant of whole blood cultures. Additionally, we detected LPS-induced IDO protein in the supernatant. Both the IDO protein expression and the degradation of Trp along the Kyn pathway indicate the activation of IDO by ex vivo LPS stimulation.
Previous studies with pigs showed that the in vivo administration of LPS increases the release of the pro-inflammatory TNF-α and the anti-inflammatory IL-10.23–25 In this study we found both TNF-α and IL-10 to be significantly elevated 1 h after LPS challenge, but not later on. The LPS-induced increase of plasma TNF-α indicates the activation of the pro-inflammatory immune response, which is reported to induce IDO activation. 26 This is supported by the present finding showing an increase of TNF-α 1 h and IDO expression 3 h after LPS injection. However, IDO activation may also be caused by LPS directly via TLR4. 27 With respect to IL-10 release, a previous study with human cells indicated that IL-10, in comparison to other T-cell cytokines, is synthesized at a late stage, i.e. 24 to 48 h after stimulation. 28 In contrast, studies in pigs and mice found significantly elevated plasma concentrations of IL-10 3 h or 1 h after i.p. LPS administration respectively.24,29 IL-10 is supposed to be a part of the anti-inflammatory immune response, and primarily mediates immunosuppressive functions by inhibition of cytokine synthesis and antigen presentation, preventing an overshoot of the pro-inflammatory immune response. 30 A study in mice has shown that pretreatment of IL-10 represses IDO production in dendritic cells indicating that the IDO activation is regulated by both pro- and anti-inflammatory cytokines. 29
It was shown that LPS can induce a release of cytokines in the brain.31–33 However, the results of the present study showed that the concentrations of TNF-α and IL-10 were unaffected by LPS treatment, at the time points measured, in the hippocampus and the PFC. It is possible that a single peripheral administration of LPS is not able to induce an increase of the cytokine concentrations in the brain. Alternatively, cytokine release in the brain follows a different temporary dynamics, which may not be covered by the time points chosen in the experiment.
Our data show that in vivo LPS stimulation induced a depletion of plasma Trp. From other studies in pigs, there are already indications that the IDO activity was increased by administration of Freund’s adjuvant.34,35 There, the increased depletion of Trp during the induced inflammation was used as a marker for IDO activation. Further studies showed that plasma Trp levels decreased during chronic lung inflammation.35,36 Although no changes in Kyn concentrations were found in plasma, increased levels of Kyn were detected in lung, lymph node, heart and spleen indicating an inflammatory increased IDO activity. 36 The single LPS administration used in the present study induced an increase of plasma Kyn, resulting in an increased plasma Kyn/Trp ratio after LPS. An enhanced Kyn/Trp ratio is a marker for IDO activation, and in different malignancies, sepsis and trauma patients it is considered as a prognostic indicator for mortality.37,38
In the present study, LPS application induced the accumulation of specific plasma Trp metabolites indicating the activation of the kynurenine pathway. The time courses of Kyna and Quin plasma concentrations were similar to that of Kyn after LPS administration. It was shown in gerbils that i.p. application of LPS together with galactosamine induced an increase of Quin in serum and tissues, and the magnitude of the Quin concentration also correlated with the increase of Kyn. 39 These findings indicate that Kyn is the rate-limiting substrate for the production of Kyna and Quin.
Western blot analyses showed that LPS induced IDO protein expression in blood and tissues. In plasma, the expression of the inducible protein form with a molecular mass of ∼42 ku correlated with the time of elevated plasma Kyn concentrations. In addition to the studies in blood, protein expression was measured in peripheral tissues and in the brain, detecting an LPS-induced IDO expression in the liver and in the lung, but not in the muscle and the analyzed brain areas. These results support previously-described findings of a tissue-specific IDO expression.26,40 Besides the inducible protein form, we detected a constitutively expressed IDO protein form in blood and liver. Western blot analyses in human eosinophils also detected two IDO forms with different molecular masses, one constitutive (∼45 ku) and one IFN-γ-inducible form (∼42 ku), 41 comparable with our results.
In addition to the in vivo experiments, we induced IDO activation in the whole blood culture system by LPS stimulation. The results of these experiments show that LPS induced an increase of TNF-α, a depletion of Trp, an accumulation of the metabolites Kyn and Quin, and IDO protein expression, indicating an activation of IDO by ex vivo LPS stimulation. The increased production of Quin could be the result of activated monocytes, which have been reported to produce Quin after stimulation with immune stimulants. 42 The detected protein pattern in the supernatants after ex vivo LPS stimulation is similar to that found in the plasma after in vivo LPS administration and showed a constitutive and an inducible IDO protein form. Thus, LPS is able to stimulate IDO activity in pigs, both in vivo and ex vivo.
In summary, the present study characterizes in vivo and ex vivo IDO activation in pigs. IDO activation was induced by LPS and described by a depletion of Trp and an accumulation of the metabolite Kyn in plasma and whole blood cultures. The LPS-induced accumulation of the metabolites Kyna and Quin indicates the activation of the Kyn pathway. IDO protein expression was detected for the first time in pigs and found to be inducible by LPS. The results of this study demonstrate that a single administration of LPS is an appropriate model for the activation of IDO in pigs, which could serve as a basis for further biomedical research on immunoregulatory IDO functions.
Footnotes
Funding
Acknowledgements
The authors are grateful to Dr Cornelia Müller for assisting in quantification of Trp and its metabolites. We thank the staff of the Research Unit Behavioural Physiology for technical assistance, particularly Birgit Sobczak and Regina Wal. Furthermore, we thank the staff of the experimental pig unit for their assistance with the handling of the animals.
Conflict of interest
The authors do not have any potential conflicts of interest to declare.