
Abstract
Purple sea urchins (Strongylocentrotus purpuratus) express a highly variable set of immune genes called Sp185/333 by two subtypes of coelomocytes: the polygonal and small phagocytes. We report that the Sp185/333 genes and their encoded proteins are also expressed in all of the major organs in the adult sea urchin, including the axial organ, pharynx, esophagus, intestine and gonads. After immune challenge, there is an increase in the level of Sp185/333 mRNA in cells associated with the intestine and axial organ. The Sp185/333 proteins increase in the axial organ, pharynx, esophagus and intestine after challenge. However, the proportion of Sp185/333-positive cells only increases in the axial organ, while there is no change in that proportion in the other organs after challenge. The size range of the major Sp185/333 proteins expressed by organs is broader (5 kDa to > 250 kDa) compared with those in coelomocytes (∼40 kDa to < 250 kDa). Images of the different organs do not clarify whether coelomocytes or parenchymal cells express the Sp185/333 proteins. The increase in levels of Sp185/333 transcripts, protein expression and Sp185/333-positive cells in the axial organ in response to challenge suggests that this organ may have an important role in immunity for this species.
Introduction
The purple sea urchin (Strongylocentrotus purpuratus) has a complex and sophisticated immune system composed of a large repertoire of gene models encoding proteins capable of recognizing and responding to a vast array of pathogens.1–6 There are a number of immune gene families in vertebrates that are greatly expanded in the sea urchin. Examples include 253 TLRs (Homo sapiens possess ∼10), 203 nucleotide-binding oligomerization domain-like (NLR) receptors (H. sapiens have 20) and 218 scavenger receptor cysteine-rich (SRCR) domain proteins (H. sapiens have about 16).1,2,6 Included among the immune response genes is the Sp185/333 gene family, which was identified originally based on significant up-regulation in coelomocytes (immune cells) responding to immune challenge3,7 and significant sequence similarity to an uncharacterized differential display sequence DD185, 7 and an expressed sequence tag EST333. 8 Because the deduced proteins have no matches to other known proteins, the gene family was named for these two unknown sequences and called 185/333, 3 which was revised to Sp185/333 to indicate the species and genus given that these genes have also been identified in other echinoids. 9
Estimates of the Sp185/333 gene family size ranges from 40–60 genes per genome, 10 and each gene is small, with two exons, of which the first encodes a signal sequence and the second encodes the mature protein.10–12 The extraordinary sequence diversity among the genes and messages derives from mosaic patterns of blocks of sequence called ‘elements’, in addition to single nucleotide polymorphisms (SNPs) and/or small insertions/deletions (indels) within individual elements.11–13 The structure of the second exon is likely the result of gene recombination, duplication, deletion and conversion generating a set of very young extant genes.10,14 RNA editing has been inferred 15 based on comparisons between the perfect full-length open reading frames from 170 genes sequenced from three animals 13 and the cDNA sequences from the same animals, 11 of which half encode truncated and missense proteins due to SNPs and small indels that are not encoded by the genes. It is noteworthy that some of the missense amino acid sequences predicted from cDNA sequences can be identified from proteins isolated from coelomocytes, 9 indicating that edited mRNAs are translated. Overall, the Sp185/333 system is a family of tightly clustered, similar genes that undergo processes of diversification, and mRNA that is edited to generate a broad array of proteins.
All of the deduced Sp185/333 proteins share overall structural similarities that include a hydrophobic N-terminal signal sequence, a glycine-rich region with an integrin binding motif (RGD), a histidine-rich region and a C-terminal region.11,12 Except for the α-helical prediction for the hydrophobic signal sequence, the amino acid sequences of the Sp185/333 proteins do not enable informative predictions of secondary structures or folding patterns, which suggests that they may be intrinsically disordered. The size of the Sp185/333 proteins isolated from coelomocytes range from 30 kDa to > 200 kDa,9,15 which is larger than predicted from the cDNA sequences (4–55.3 kDa). 11 This appears to be due multimerization, which may be augmented by post-translational modifications and results in as many as 264 distinct proteins in an individual sea urchin. 9
The Sp185/333 proteins are localized to two subtypes of coelomocytes—polygonal phagocytes and small phagocytes—where they associate with membranes and are localized to perinuclear vesicles, and are present on the cell surface of small phagocytes.15,16 Following immune challenge in vivo, the proportion of Sp185/333-positive (Sp185/333+) cells in the coelomic cavity significantly increases relative to pre-challenge. 15 However, it is not known whether Sp185/333 proteins are present in various organs and tissues throughout the body. Therefore, we evaluated both immunoquiescent (immune down-regulated) sea urchins and animals challenged with LPS and found that Sp185/333 proteins are present in the axial organ, pharynx, esophagus, intestine and gonads. Sp185/333 gene expression increases in intestine, axial organ and coelomocytes after immune challenge compared with immunoquiescent animals. Sp185/333+ cells are present in all organs, and the proportion of Sp185/333+ cells in the axial organ and coelomocytes is higher in immune-challenged animals compared with non-challenged control animals. Similarly, the Sp185/333 protein content is higher in the pharynx, esophagus, intestine, axial organ and coelomocytes from immune-challenged animals compared with samples from non-challenged control animals. The size range of the major Sp185/333 proteins in organs is < 5 kDa to > 250 kDa, which is broader than that observed previously for coelomocytes.9,15 There are no observable changes in the repertoires of Sp185/333 proteins in organs from the immune-challenged animals compared with control animals. Overall, these results indicate that the Sp185/333+ cells are not restricted to the coelomic cavity, but are found throughout the organs and tissues. These immune response proteins are either produced by somatic cells within the organs, in addition to being produced by coelomocytes, or the Sp185/333+ coelomocytes are present in all tissues throughout the body.
Materials and methods
Sea urchins
Purple sea urchins, S. purpuratus, were supplied by Marinus Scientific (Long Beach, CA, USA) and maintained in aquaria as described. 17 Animals were fed weekly with commercial rehydrated kelp (Quickspice, Commerce, CA, USA).
All animals (n = 22) chosen for the study were either immunoquiescent (Iq) through acclimation to the aquaria for > 8 months without significant disturbance,17,18 or were not allowed to acclimate to an aquarium [non-acclimated (N-Ac); n = 2] and used within 2 wk of shipping.
Immunological challenge
Experimental groups, based on analysis performed on sea urchin coelomocyte and organ samples.
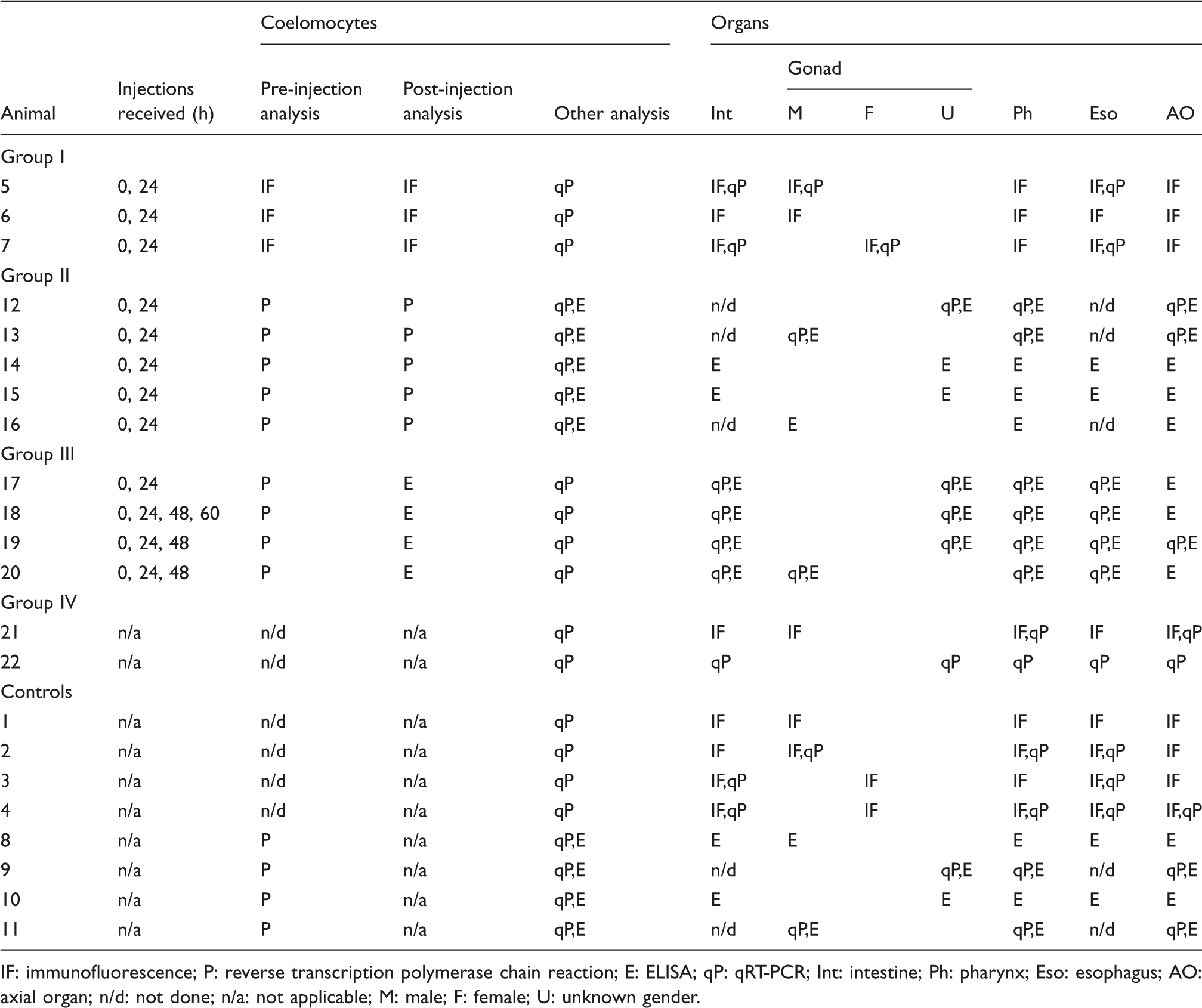
IF: immunofluorescence; P: reverse transcription polymerase chain reaction; E: ELISA; qP: qRT-PCR; Int: intestine; Ph: pharynx; Eso: esophagus; AO: axial organ; n/d: not done; n/a: not applicable; M: male; F: female; U: unknown gender.
Antisera
Three polyclonal rabbit antisera (anti-185-66, −68, and −71) were raised against three conserved synthetic peptides present in the most common Sp185/333 cDNA sequence, as described previously.9,15 Anti-185-66 recognizes a sequence near the N-terminus and adjacent to the leader, anti-185-68 recognizes a sequence in the middle of the proteins near the RGD motif and anti-185-71 recognizes a sequence near the C-terminus.9,15
Immunocytology
Coelomocyte collection
Whole coelomic fluid (fluid plus cells; 200–300 µl) was collected 15 min prior to first injection and 24 h after the last LPS injection. Fluid was withdrawn directly from the coelomic cavity by piercing the peristomial membrane with a 23-gauge needle attached to a 1-ml syringe. The syringe was pre-loaded with 0.3–0.5 ml ice-cold calcium and magnesium-free sea water that included 70 mM EDTA and 50 mM imidazole, pH 7.4 (CMFSW-EI). 20 After withdrawal, the whole coelomic fluid was mixed with additional CMFSW-EI that was added to the contents in the syringe for a final dilution of whole coelomic fluid of 1 : 3 in CMFSW-EI. Syringe contents were expelled into a 1.5-ml microcentrifuge tube on ice. Cells were counted with a hemocytometer.
Coelomocyte fixation
Cells (1 × 105 in 100 µl CMFSW-EI) were spun onto poly-
Tissue preparation
Whole organs or parts of organs (axial organ, pharynx, esophagus, intestine and gonad) were
removed from N-Ac (n = 1), Ch (n = 3) and control
(n = 4) sea urchins, and placed immediately in prefix (4% paraformaldehyde;
0.000025% glutaraldehyde in CCM) with gentle rocking at rt for approximately 8 h, followed by
stationary incubation at 4
Tissue sections
A cryostat (Hacker-Bright Safecut 7000 or Microm HM 505E; Zeiss, Oberkochen, Germany) was used to
cut 3–10-µm frozen sections of organs according to machine specifications. Both longitudinal and
transverse sections were obtained for pharynx, intestine and axial organ, and longitudinal sections
were cut for esophagus and gonads. Sections were bound to Fisherbrand® Superfrost®/Plus microscope
slides (Thermo Fisher Scientific, San Jose, CA, USA) by incubation at 25–35
Coelomocyte and tissue staining
Fixed coelomocytes and tissue sections were incubated with blocking solution (2% v/v normal goat serum and 1% BSA in PBS) in a humid chamber at rt. Cells or sections were incubated with an equal mixture of anti-185-66, −68 and −71 antisera (1 : 4000 dilution in blocking solution) plus mouse monoclonal anti-actin Ab (1 : 600 dilution; MP Biomedicals, Solon, OH, USA) for 1 h at rt in a humid chamber. Slides were washed with PBS as described above and incubated with a mixture of goat anti-rabbit-Ig (GαR-Ig) conjugated to AlexaFluor 568 (1 : 400 dilution; Pierce Biotechnology, Rockford, IL, USA) and donkey anti-mouse-Ig (DαM-Ig) conjugated to AlexaFluor 488 (1 : 200 dilution; Pierce Biotechnology) for 1 h at rt in a humid chamber. Negative controls included normal rabbit serum substituted for the primary antisera using the same dilution in blocking buffer followed by the secondary Ab. Samples were washed with PBS as described above and mounted with ProLong® Gold Antifade with 4′,6-diamidino-2-phenylindole (DAPI; Invitrogen, Carlsbad, CA, USA).
Microscopy
Whole fixed organs or freshly dissected unfixed organs were viewed and digital images were recorded using a Wild M5 dissecting microscope. Coelomocytes and tissue sections were examined with an Axioplan fluorescence microscope (Carl Zeiss Microscopy, Jena, Germany). Cells were examined with either a 10 × NA 0.25 plan phase, 20 × NA 0.5 or 40 × NA 0.75 planapochomatic phase contrast objective lens connected to a CCD camera (Hitachi, Tokyo, Japan). The Axioplan microscope was used to collect epifluorescent images of stained cells. Images were also recorded for ovary and axial organ on an Axio Examinar upright spectral confocal laser-scanning microscope (CLSM, Zeiss LSM 710). Images were captured using either 20 × /0.8 M27 D = 0.55 or 63 × /1.40 Oil DIC plan-apochromatic phase contrast objective lens.
Sp185/333+ cell quantitation
The Olympus MicroSuite™ B3SV software program was used to quantify the number of Sp185/333+ cells compared to the total number of cells. Cells in 5–13 fields were counted for each slide. Between 1 and 3 sections were analyzed for each tissue.
RT-PCR
Sample preparation and RNA isolation
Coelomocytes were collected as described above, counted on a hemocytometer, and approximately
105–107 cells were transferred to 1.5-ml microcentrifuge tubes and centrifuged
at 10,000 g for 1–2 min at 4
Reverse transcription
Total RNA samples were reverse transcribed using SuperScript™ III reverse transcriptase
(Invitrogen) with 7-µM random hexamer primers (Operon Technologies, Huntsville, AL, USA) according
to the manufacturer’s instructions. An initial PCR was performed with SpL8 primers
(SpL8F: CAG CGT AAG GGA GCG GGA AGC GTC TT; SpL8R: GTT TGC CGC AGA
AGA TGA ACT GTC CCG TGT A) that amplified sea urchin SpL8 (encodes a homologue of
the human ribosomal L8 protein; GenBank accession number R62029) to examine the cDNA quality and to
evaluate the presence of genomic DNA (the primers were positioned on either side of a small intron).
Sp185/333 gene expression was evaluated with a pair of primers (F5; GGA ACY GAR GAM
GGA TCT C and 3′UTR; AAA TTC TAC ACC TCG GCG AC) that amplified a 200-base pair fragment, which
corresponded to the 3′ end of the coding region plus part of the 3′ untranslated region
(3′UTR).
12
PCR reactions
employed 0.4 µM of each primer, 0.2 mM of each deoxynucleotide, 2 mM MgCl2, 1 × company
supplied buffer, 0.1 U of Taq DNA polymerase (Thermo Fisher Scientific), plus 2 µl
of the reverse transcriptase reaction (cDNA template) in a total volume of 12.5 µl. Reactions were
performed in either an iCycler IQ in non-quantitative mode (Bio-Rad Laboratories, Hercules, CA,
USA), a PTC-200 Peltier Thermo Cycler (MJ Research, Ramsey, MA, USA) or a GeneAmp PCR System 9600
(PerkinElmer, Waltham, MA, USA). The PCR program was 95
Quantitative RT-PCR
Quantitative RT-PCR (qPCR) was performed on the same cDNA samples used for RT-PCR.
SpL8 cDNA was amplified with SL8F (CAC AAC AAG CAC AGG AAG GGA) and ASL8 (AGC GTA
GTC GAT GGA TCG GAG T), and a region of the Sp185/333 sequence was amplified using
the primers F5 and 3′UTR (see above). Each qPCR reaction contained 1 × Absolute QPCR SYBR
Fluorescein Mix (ABgene, Surrey, UK), 0.5 µM of each primer, and 1–2 ng of cDNA in a total volume of
20 µl. A standard curve was generated using five 10-fold serial dilutions
(103–107 plasmids/sample) of a Sp185/333 gene cloned from
coelomocyte genomic DNA (GenBank accession number EF607663.1). qPCR reactions were performed on an
iCycler IQ (Bio-Rad Laboratories) in duplicate using the following program: 95
ELISA
Sample preparation
The axial organ, pharynx, esophagus, intestine and gonads were dissected from Iq
(n = 4) and N-Ac (n = 9) animals. Each sample of whole coelomic
fluid was centrifuged (see above), the cell-free coelomic fluid was decanted and the pelleted cells
were frozen in liquid nitrogen. Tissue samples were placed directly into 1.5-ml microcentrifuge
tubes and frozen immediately in liquid nitrogen. All samples were stored at −70
Protein content analysis and ELISA
Total protein content in each duplicated sample was determined using a Micro BCA™ Protein Assay Kit (Pierce Biotechnology). The second aliquot (see above) for each sample was thawed, adjusted to 0.136 M NaCl by adding distilled water and samples were distributed in duplicate into 96-well ELISA plates (Corning, Corning, NY, USA). Samples of 10 µg, 5 µg and 2.5 µg of total protein were distributed into wells containing TBS (20 mM Tris, 0.136 M NaCl, pH 7.4) for a final volume of 100 µl per well. Proteins were bound to EIA/RIA plates (Corning) at rt with gentle agitation for 2 h followed by 3 washes of 5 min each with 150 µl per well of TBST (TBS containing 0.5% Tween-20). Plates were blocked with 4% BSA in TBS at rt with gentle agitation for 1.5 h followed by 3 washes with TBST (5 min each). Plates were incubated with 100 µl of anti-185 antisera (1 : 1000 dilution of each anti-185-66, −68 and −71 in TBS) per well at rt for 1.5 h with gentle agitation. Plates were washed and incubated with GαR-Ig conjugated to alkaline phosphatase (1 : 15,000 dilution in TBS; Sigma-Aldrich) at rt for 1.5 h with gentle agitation. Samples were washed again and incubated with 50 µl of p-nitrophenyl phosphate (1 mg/ml in 1 M diethanolamine, pH 9.8; Sigma-Aldrich) for 10 min in the dark. The reaction was stopped by adding 50 µl of 3 M NaOH and absorbance was read at 405 nm on a Spectra MAX microplate spectrophotometer (Molecular Devices, Sunnyvale, CA, USA). Controls substituted buffer for coelomic fluid proteins or the primary antisera were substituted with normal rabbit serum (1 : 1000 dilution in TBS).
Western blots
Sample preparation
Freshly collected samples were frozen and stored at −70
Protein separation and transfer
Samples (1–50 µg/sample) were heated to 70
Ab staining
Membranes were incubated with an equal mixture of anti-185-66, −68 and −71, or pre-bleed serum (1 : 15,000 dilution for each antiserum in blotto) while rocking at rt for 1.5 h followed by 3 washes with TNT. Membranes were incubated with GαR-Ig conjugated with HRP (GαR-Ig-HRP; 1 : 30,000 dilution in blotto; Pierce Biotechnology) while rocking for 1.5 h at rt. Membranes were washed twice in TNT and once in TN, followed by 5 min in either Super Signal West Pico Chemiluminescent Substrate System (Pierce Biotechnology) or Western Lightning™ Chemiluminescence Reagent (PerkinElmer). Membranes were exposed to Classic Blue Autoradiography film (MidSci, St. Louis, MO, USA).
Statistical analysis
Statistical analysis of the results from tissue and coelomocyte samples were performed using SAS v. 9.1.3 (SAS, Carey, NC, USA). The relative gene expression ratio (R) for qPCR was calculated using the equation (Etarget)ΔCPtarget( control-sample )/(Eref)ΔCPref( control-sample ), where target refers to the Sp185/333 target sequence and ref refers to the SpL8 reference sequence. 22 Data sets of raw measurements for immunofluorescence (IF) and ELISA, or R values for qPCR, were log10-transformed to approximate the normal overall distributions and statistical significance was assessed by ANOVA in general linear models regression procedures. Bonferroni correction was applied to the pairwise means comparison in order to contrast groups of values, such as different treatments, organs and individual animals, and the significance levels shown are post-Bonferroni. Experimental error was addressed by including replicate measurements among model parameters.
Results
Preliminary analysis of immune activation and definitions of experimental groups
Individual sea urchins were assessed for their level of immune activation by evaluating small
samples of coelomocytes. Results from the preliminary analyses were used to select animals for a
more extensive analysis of coelomocytes and organs for Sp185/333 gene expression
and protein content that are reported below. The preliminary analysis of the coelomocytes employed
three different approaches prior to and after challenge with LPS, and were designed to detect
changes in (i) the percentage of coelomocytes that were Sp185/333+, (ii) the level of
Sp185/333 transcripts, and/or (iii) the Sp185/333 protein content. Animals were
categorized into four experimental groups (I–IV) based on the combination of method(s) used in the
preliminary analysis of coelomocytes to determine whether the animals had responded to the LPS
challenge (Figure 1; Table 1). Flow chart of preliminary experimental analysis of coelomocytes and organs. The immune status of
the sea urchins that were chosen for analysis is indicated as either immunoquiescent (Iq) or as not
acclimated to the aquaria (non-acclimated; N-Ac) with unknown immune status. The area bounded by the
light blue box indicates preliminary analysis that was done on coelomocytes to determine whether sea
urchins had responded to LPS and had become immune challenged (Ch). Coelomocytes were evaluated
using different combinations of analyses (groups I–III; indicated in bold colors), including (i) an
evaluation of the number of Sp185/333+ cells (light green), (ii) an evaluation of
Sp185/333 gene expression (light orange) (iii) and an evaluation of the S185/333
protein content (lavender). Three groups of controls were employed: Iq animals that were evaluated
for Sp185/333 gene expression (dark gray), another group of Iq animals that were
not evaluated (light gray), and a third group of animals that were N-Ac and were not evaluated (dark
purple). After the preliminary analyses were completed on coelomocytes, selected sea urchins were
sacrificed and the organs and coelomocytes were collected for further evaluation (area bounded by
the pink box). Results from the post-challenge samples are reported in this article. Colors of the
boxes indicating the groups and controls are also used in Figures 2, 5, 9 and 10. *Indicate that data from control animals were combined for
some analyses (see Table 1).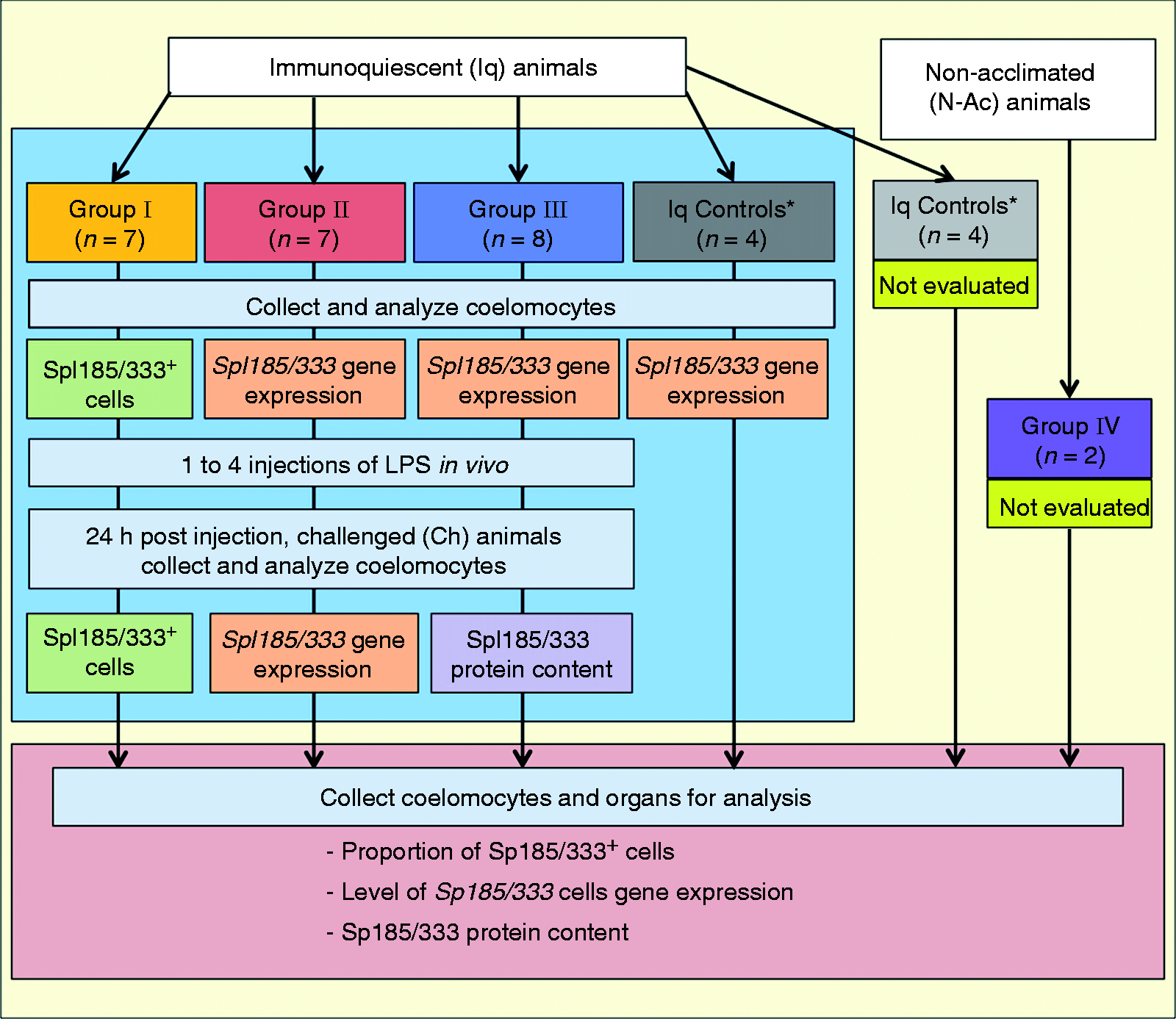
Preliminary analysis of coelomocytes from animals in group I was based on IF assessment of coelomocytes to determine the percentage of cells that were Sp185/333+ before immune challenge and 24 h after a second injection of LPS (Figure 1). Animals showing approximately a twofold increase in the percentage of Sp185/333+ coelomocytes after challenge compared with the pre-challenge state of immunoquiescence (3 of 7 animals; Table 1, animals #5–7) were chosen for further analysis. Coelomocytes from animals in group II were evaluated for Sp185/333 transcript level before and 24 h after immune challenge using RT-PCR (Figure 1). Gel images of the amplicons were used to estimate the change in Sp185/333 transcript level in coelomocytes induced to Ch from Iq animals. Those animals that showed a substantial increase in Sp185/333 transcript level in response to immune challenge were chosen for further study (5 of 7 animals; Table 1, animals #12–16). Coelomocytes from animals in group III were collected 24 h after 2 or more injections of LPS and evaluated using 2 methods (Figure 1). RT-PCR was used to estimate the Sp185/333 transcript level prior to immune challenge and ELISA was used to evaluate the Sp185/333 protein content in coelomocytes after immune challenge. Animals in group III that had a lower level of Sp185/333 transcripts prior to immune challenge were assumed to have a lower level of Sp185/333 protein content compared to post-challenge. Immune-challenged animals that had a twofold higher Sp185/333 protein content in the coelomocytes over the internal control background levels were chosen for dissection and analysis of the organs (4 of 8 animals; Table 1, animals #17–20). Group IV animals were N-Ac animals and assumed to be immune activated upon delivery to the laboratory. Consequently, immune activation through LPS injection was not administered and samples for preliminary evaluation were not collected (Figure 1, Table 1, animals #21–22). Coelomocytes from a subgroup of the controls were analyzed for Sp185/333 transcripts using RT-PCR to demonstrate no expression prior to organ collection (Figure 1; Table 1, animals #8–11). The remaining control animals were assumed to be Iq and no preliminary analysis was done on coelomocytes prior to organ collection (Figure 1; Table 1, animals #1–4).
Sp185/333 transcripts increase in intestine, axial organ and coelomocytes after immune challenge with LPS
Increases in Sp185/333 gene expression has been demonstrated previously by
non-quantitative methods;3,7,11 consequently, we repeated the evaluation of gene
expression in coelomocytes using quantitative methods, and found that Ch animals in groups I–III
showed a significant increase in the relative Sp185/333 transcript level after
immune challenge compared with the same animals before challenge (P < 0.05;
Figure 2). However, results for group IV did
not show increased transcript level, which indicated that although these N-Ac animals were assumed
to be immune activated, the coelomocytes did not contain an elevated level of
Sp185/333 transcripts compared with coelomocytes from animals in groups I–III that
had received LPS. When the relative Sp185/333 mRNA levels in coelomocytes collected
prior to immune challenge in groups I–III were compared with coelomocytes from the control group, no
differences in the relative transcript levels were observed (data not shown). These results
demonstrated that qPCR could be used to evaluate Sp185/333 transcript content in
coelomocytes and that the results were in agreement with previous reports.3,7,11
Sp185/333 gene expression increases in coelomocytes in sea urchins after immune
challenge. qPCR was used to determine the Sp185/333 gene expression in coelomocyte
samples relative to SpL8 expression. Animals are grouped by treatment, which
includes post-immune-challenged animals (Ch; bars to the left of the vertical dotted line),
non-acclimated animals (N-Ac; bar to the right of the vertical solid line) and control animals. The
combined results for coelomocytes collected from Iq plus control animals (light and dark gray as in
Figure 1) were set to 1 and are represented
by the horizontal dotted line. Ch animals are separated into Groups I–III, and N-Ac animals are in
Group IV (see Table 1; group colors as
in Figure 1). Significant differences
(*P < 0.05 or **P < 0.0001) are in comparison to
Sp185/333 expression in coelomocytes from respective Iq or control animals. SEM are
shown.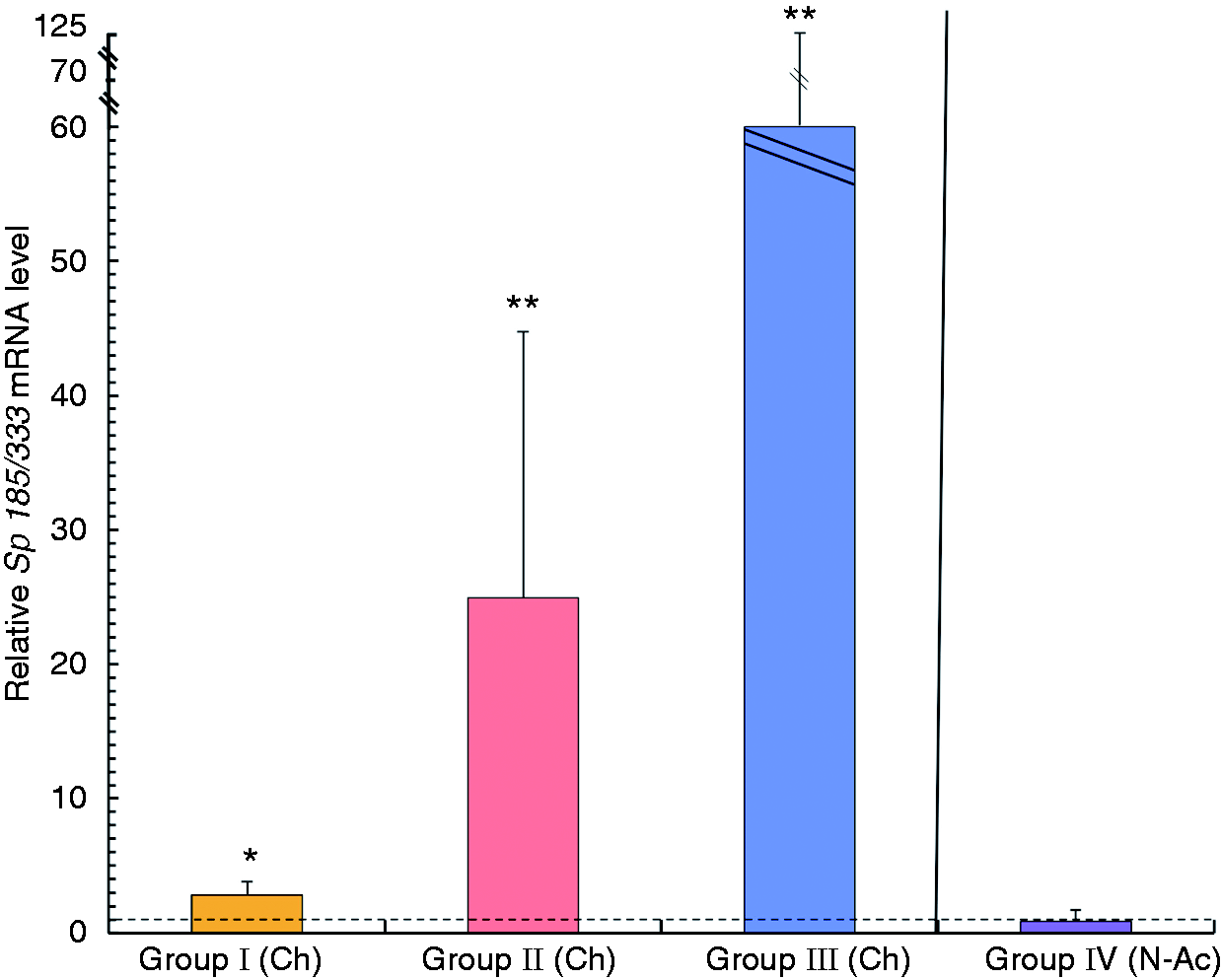
Although Sp185/333 gene expression in coelomocytes has been well documented
(reviewed in Ghosh et al.
23
and Smith
24
) it is not known
whether these genes are expressed by cells in organs. Therefore, we undertook a preliminary analysis
of organ samples from S. purpuratus and found that every organ showed
Sp185/333 gene expression (Figure
3). Because Sp185/333 transcripts increase in coelomocytes after exposure to
bacteria or to several different pathogen-associated molecular patterns,3,7,9,11 we measured whether Sp185/333
transcript level changed in organs after LPS injections compared with organs from control animals.
Quantitative analyses of the relative Sp185/333 transcript level in organ samples
for Ch animals in groups I–III, as well as N-Ac animals in group IV (Table 1), were compared with the control samples. Results
showed an increase in the relative Sp185/333 transcript levels in the intestine for
groups III and IV animals after challenge (P < 0.05; Figure 4). There was also an increase in the relative
Sp185/333 transcript level in the axial organ for group III compared with controls
(P < 0.05; Figure 4),
but elevated transcripts were not observed for samples from other groups. No significant changes in
the relative Sp185/333 transcript level after immune challenge were observed in
gonad, esophagus or pharynx for any experimental group. Overall, these results provided initial
evidence that Sp185/333 expression in intestine and axial organ in Ch and N-Ac
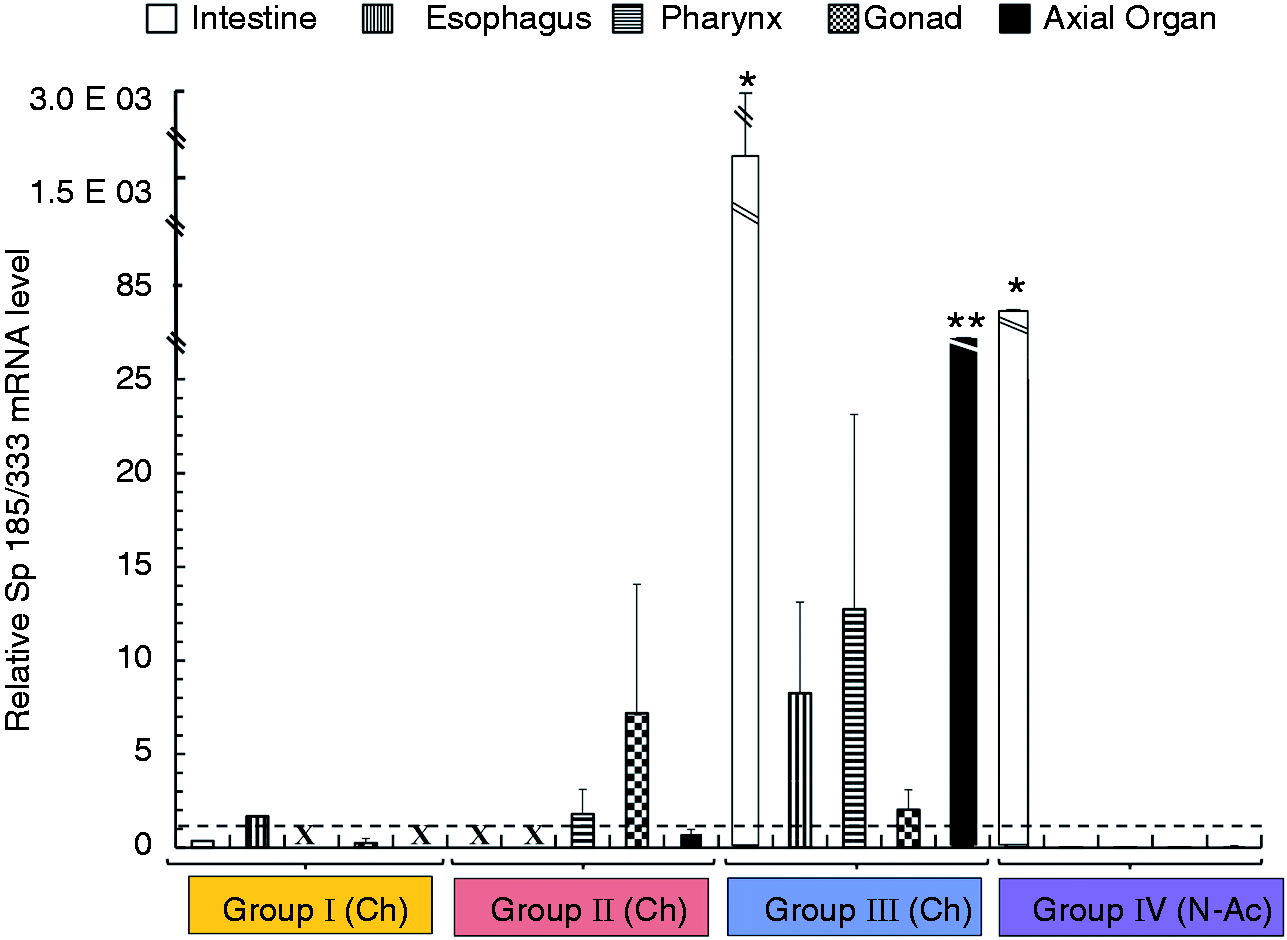
animals was responsive to immune challenge. Sp185/333 genes are expressed in organs. RT-PCR was used to analyze gene
expression in organs collected 24 h after LPS injection. cDNA samples were amplified with a pair of
primers (F5 and 3′UTR) that target a portion of the full-length Sp185/333
transcript, and a different pair of primers that target the SpL8 transcript
(encodes a homologue of the human ribosomal L8 protein), which was used to assess sample quality. C:
coelomocyte; Es: esophagus; Ph: pharynx; Go: gonad; In: intestine; AO: axial organ; +: Ch animal
that was challenged with LPS; −: Iq animal; n/a: not applicable. Sp185/333 gene expression increases in organs of sea urchins after immune
challenge. qPCR was used to determine the relative Sp185/333 gene expression in
coelomocytes and organ samples relative to SpL8 expression. Animals are grouped by
treatment, which includes Ch and N-Ac animals (Groups I–IV, colors as in Figure 1) and control animals (light and dark gray as in Figure 1; see also Table 1) which were set to 1 and represented by the horizontal
dotted line. Significant differences in Sp185/333 gene expression
(*P < 0.05 or **P < 0.0001) are relative to the expression
in the same organs from the control animals. X indicates missing data. SEM are shown.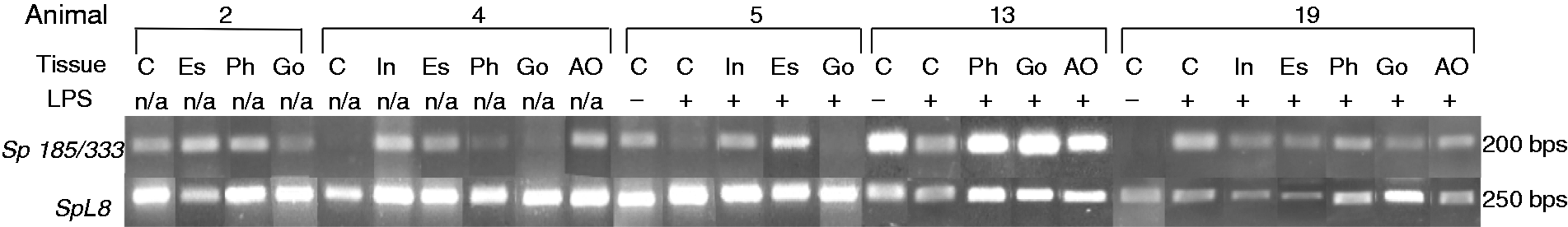
Sp185/333 proteins increase in coelomocytes, intestine, esophagus, pharynx and axial organ after immune challenge with LPS
Previous work demonstrates that Sp185/333 proteins in coelomocytes increase significantly with a
∼twofold increase in protein content between 24 and 48 h after LPS injection compared with control
animals that were sham injected.
9
Because Sp185/333 gene expression was observed in all major organs,
and the level of mRNA significantly increased in some of these organs after immune challenge (see
above), we assumed that there would also be an increase in the level of Sp185/333 proteins in organs
from Ch animals. Therefore, we analyzed the Sp185/333 protein content in organ samples from animals
that received LPS injections compared with samples from control animals. We also compared the
Sp185/333 protein content in coelomocytes from the Ch animals before and after challenge to confirm
that the immune response was activated in these animals. Results for coelomocytes from animals in
group III (Figure 1; Table 1) showed an increase in the level of Sp185/333 proteins
in response to LPS challenge compared with the pre-challenge time point
(P < 0.01); however, group II animals did not show a similar increase (Figure 5). We also evaluated the level of
Sp185/333 proteins in organ samples for groups II and III after injections and compared them to the
protein level in organ samples from control animals. Results indicated that group III had an overall
increase in Sp185/333 proteins in the pharynx (P < 0.05), esophagus, intestine
and axial organ (P < 0.01); however, there was no change in the level in testes
for Ch animals compared with control animals for this group (Figure 5). Animals in group II showed no significant change in
Sp185/333 protein content in any organ after immune challenge compared with the controls, similar to
the results from coelomocytes for that group. Overall results from immune-challenged animals for
coelomocytes and organs for group III animals showed an increase in the level of Sp185/333 proteins
for both coelomocytes and cells associated with the organs, while the level of Sp185/333 proteins
from group II animals did not change after immune challenge. These results reflect the preliminary
analyses of coelomocytes in response to immune challenge that were done prior to organ collection.
Coelomocytes from group II were elevated for Sp185/333 mRNA level, while
coelomocytes from group III were evaluated for Sp185/333 protein content (Figure 1; Table 1). Preliminary evaluation of coelomocytes in group III showed an increase in
Sp185/333 protein content and these changes were also observed in the organs in the subsequent
experimental analysis (Figure 5). However,
it was assumed that because the level of Sp185/333 transcripts had increased in
coelomocytes from group II after immune challenge, there would be a correlated increase in protein
content for the organs. However, results for the organs from group II animals did not reflect these
changes. Furthermore, all sea urchins in group II received two injections of LPS, while most of the
animals in group III received three or more injections prior to organ collection (Table 1). The preliminary evaluation of
coelomocytes for the level of Sp185/333 mRNA may not be a good predictor of protein
levels in the organs. However, three injections of LPS that were administered to animals in group
III may have been required to increase the level of Sp185/333 proteins above the pre-injection
controls. This is consistent with the level Sp185/333 proteins in group II animals that did not
change in response to two injections of LPS. Sp185/333 protein content increases in coelomocytes, intestine, esophagus, pharynx and axial
organ in sea urchins after immune challenge. Treatment groups include animals prior to challenge
with LPS (Iq, white bars), post-immune challenge (Ch, group colors as in Figure 1), and controls (gray bars as in Figure 1). Significant differences (brackets with asterisk,
P < 0.05) are compared to protein content from control animals. SEM are
shown.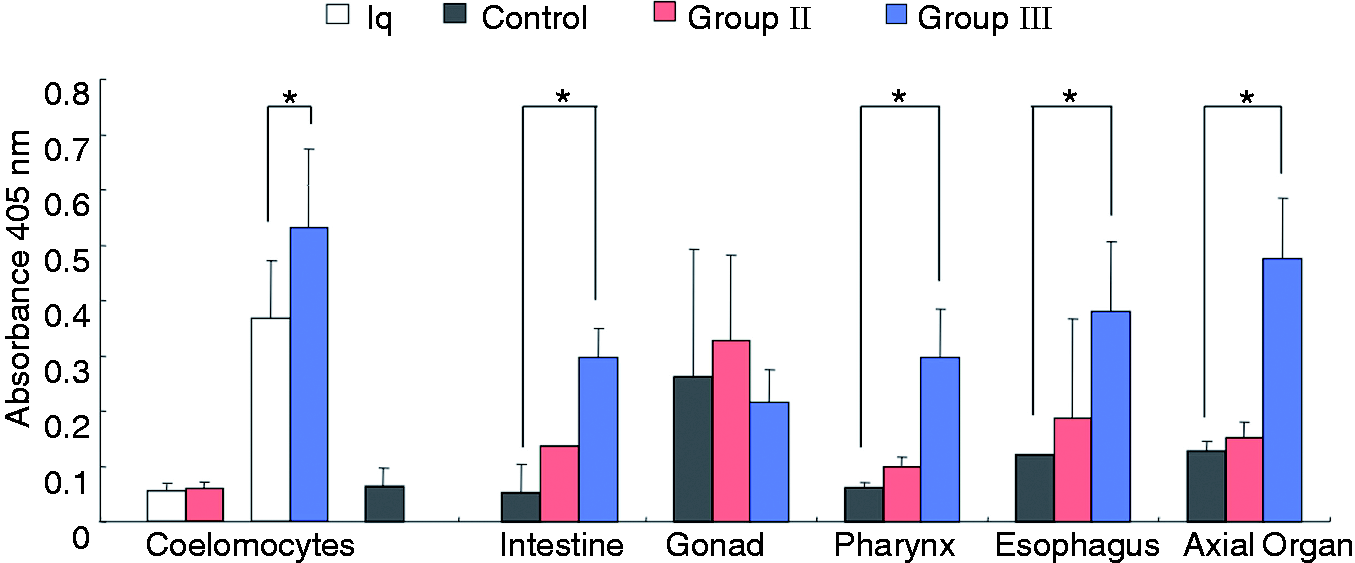
Sp185/333+ cells in the axial organ and Sp185/333+ coelomocytes increase after immune challenge with LPS
Because the Sp185/333 proteins were detected in all of the organs, we next evaluated the location
of the Sp185/333+ cells in organ samples using IF (Table 1). Sp185/333+ cells within the axial organ
(Figure 6A) were dispersed throughout the
organ, lining various canaliculi or lacunae, and, in some cases, were located within these spaces
(Figure 6B–D). In the intestine (Figure 7A) and esophagus (Figure 7B), Sp185/333+ cells were present within the
columnar epithelium lining the lumen of the intestine and along the coelomic epithelium covering the
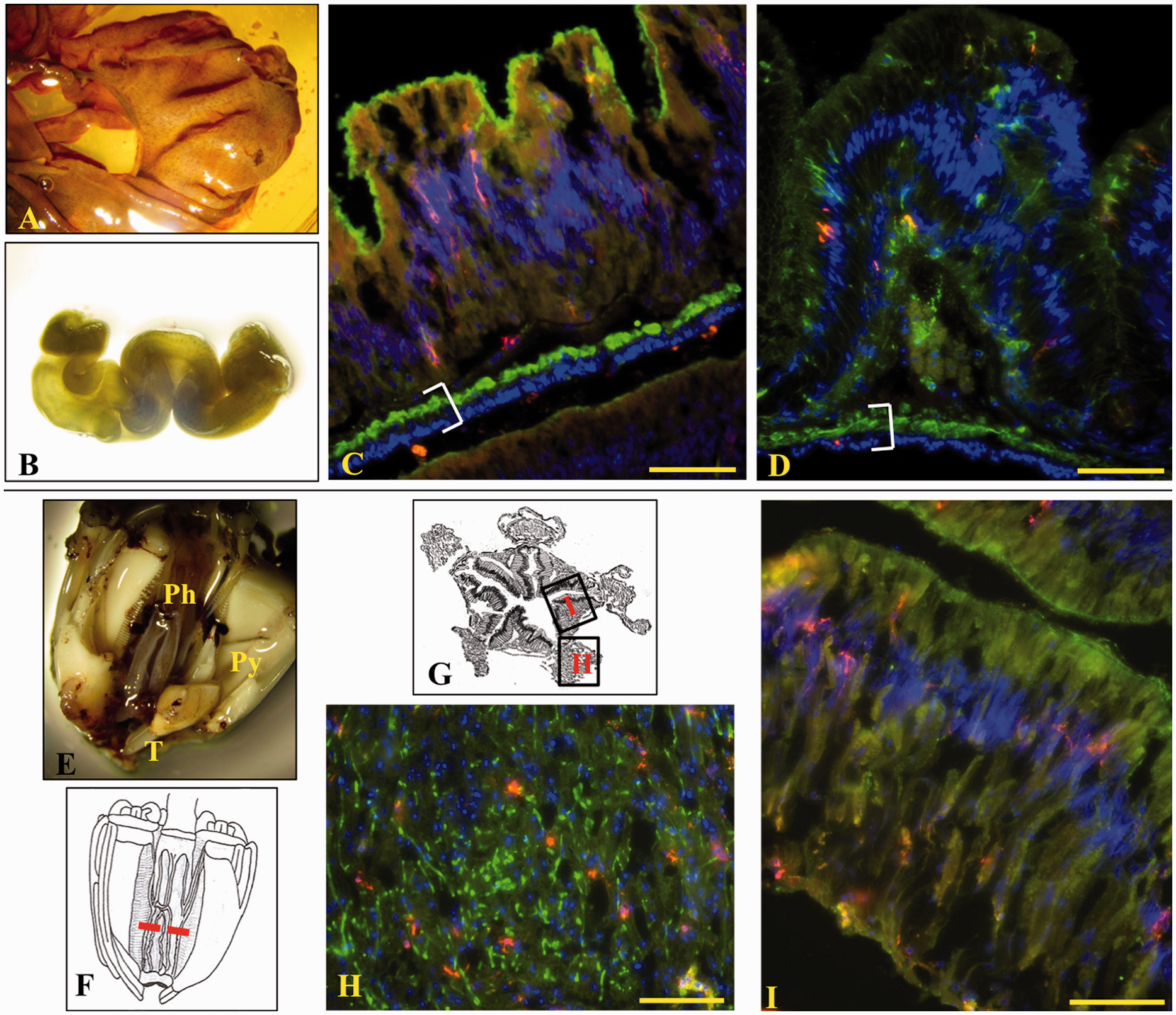
gastro-intestinal (GI) tract and in contact with the coelomic fluid (Figure 7C, D). Sp185/333+ cells were not observed in the lumen of the GI tract (pharynx,
esophagus and intestine). The pharynx (Figure
7E–G) is a complex structure with
columnar epithelium lining the lumen of the mouth cavity and connective tissue connecting the
pharynx to the lantern structure.
25
Here, Sp185/333+ cells were dispersed throughout the pharynx in both the
peripheral connective tissue (Figure 7H) and
the columnar epithelium (Figure 7I).
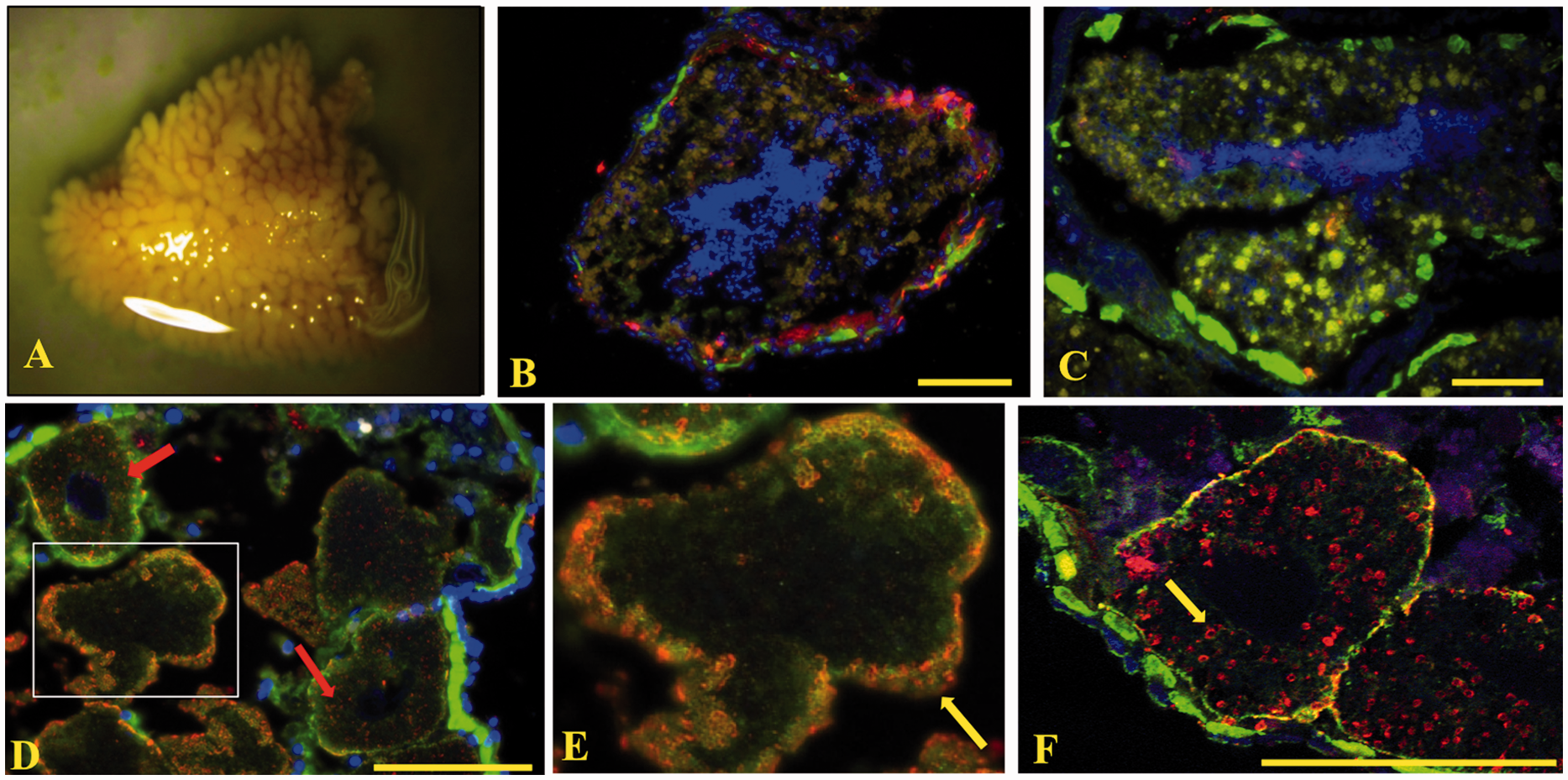
Sp185/333+cells were scattered throughout the testes (Figure 8A shows a mature gonad), both in the center near mature
sperm and along the coelomic epithelium (Figure
8B, C). Sp185/333 proteins were found
within the ovaries; however, the patterns of the Sp185/333 proteins were associated with ring
structures localized to the periphery along the outer rim of mature eggs (Figure 8D, E, yellow arrow). In the oocytes, the Sp185/333 proteins were present as ring structures
scattered about the cytoplasm (Figure 8D,
red arrows; Figure 8F, yellow arrow) in a
pattern consistent with cortical granules. The Sp185/333 proteins may also be present in follicular
cells that surround the eggs and oocytes.26–28
Sp185/333+ cells are present in axial organ. (A) A whole fixed axial organ. (B–D)
Sp185/333+ cells are present within the various spaces and dispersed throughout the axial
organ. Sp185/333+ cells are red, actin is green and DNA is blue. Image A was recorded on
a Wild M5 dissecting microscope. Image B was recorded on an Axioplan fluorescence microscope
(Zeiss). (C), (D) Paired images recorded on a spectral CLSM (LSM 510 META, Zeiss). Images B–D are
transverse sections of the organ. Bar is 100 µm. Sp185/333+ cells are present in the pharynx, esophagus and intestine. (A) A fixed
portion of the intestine. (B) Whole fixed esophagus. Longitudinal sections are shown for the
intestine (C) and the esophagus (D). Sp185/333+ cells (red) are present throughout
columnar epithelium (green, actin; blue, DNA). White brackets enclose the region containing two
layers of muscle fibers and the coelomic epithelium. (E) Whole fixed Aristotle’s Lantern (Py:
pyramid; T: tooth) with a portion cut away to show the tubular pharynx (Ph). (F) Drawing of the
Aristotle’s Lantern in which the red horizontal dotted line indicates the location of transverse
sections that were evaluated by IF. (G) Drawing of a transverse section through the pharynx. (H) and
(I) correspond to images H and I. Sp185/333+ cells are located throughout the connective
tissue (H) and columnar epithelium (I) of the pharynx. Images A, B and E were recorded a Wild M5
dissecting microscope. Images C and D and H–I were recorded on an Axioplan fluorescence microscope
(Zeiss). Bar is 100 µm. Sp185/333+ cells are present in the gonad. (A) A segment of a gonad. (B, C) Testes.
Sp185/333+ cells (red) are present in association with the coelomic epithelium (B)
surrounding the testes and within the testes with the mature sperm (C) (green, actin; blue, DNA).
(D–F) Ovary. Sp185/333 proteins appear in a scattered pattern throughout the oocyte (red arrows),
but localized to the outer portion of the egg (E; yellow arrow), perhaps in cortical granules and
follicular cells. An enlarged portion of image D is shown in E. (F) In the oocytes (yellow arrow),
some of the Sp185/333 staining pattern is crescent-shaped and similar in shape to cortical granules.
Image A was recorded with a Wild M5 dissecting microscope. Images B–E were recorded with an Axioplan
fluorescence microscope (Zeiss). Image F was recorded with a spectral CLSM (LSM 510 META, Zeiss).
Longitudinal sections of IF images are shown. Bar is 100 µm.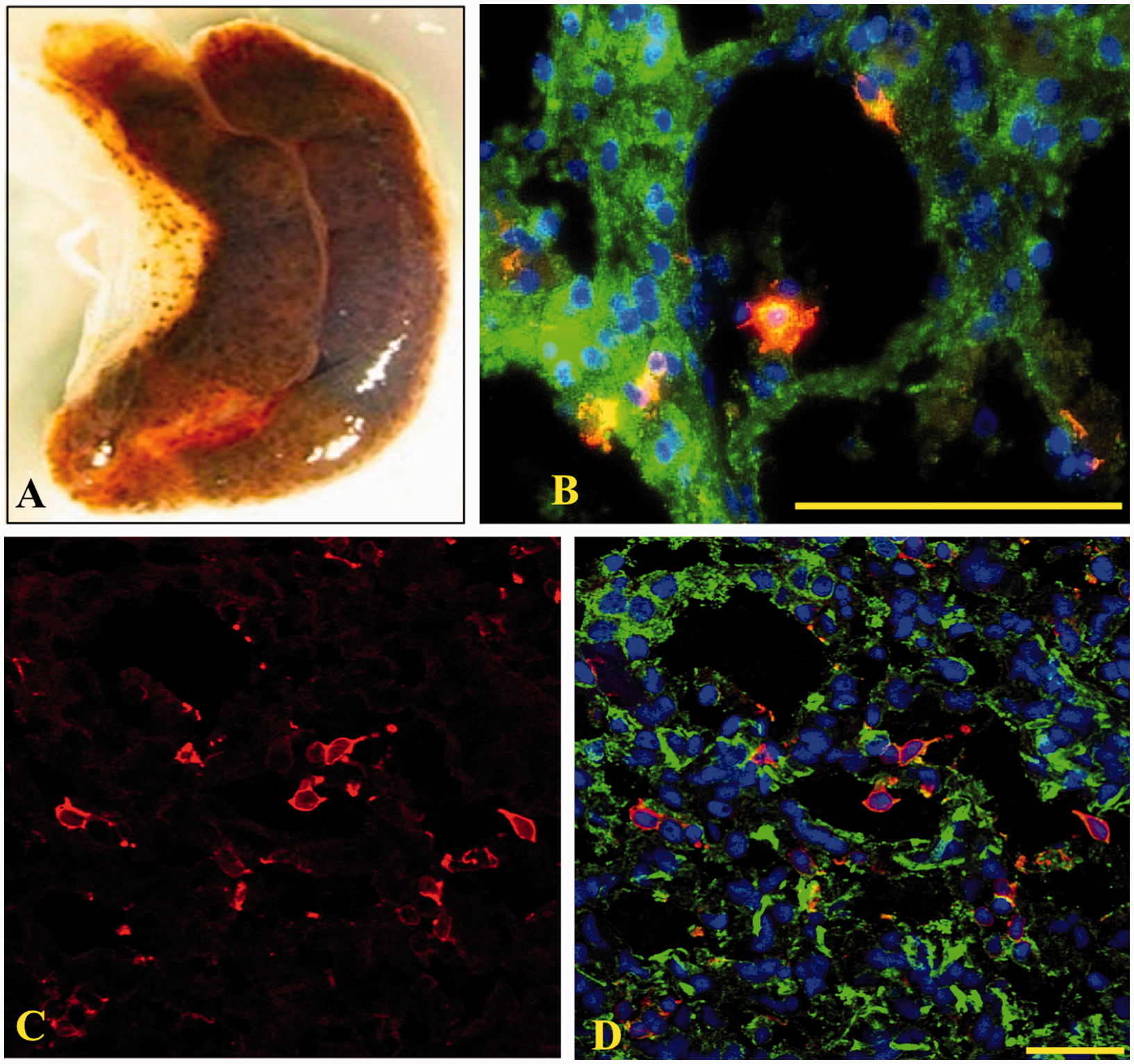
Because Sp185/333+ cells were present in all organs that were examined, we next
evaluated whether there were changes in the percentages of Sp185/333+ cells in the organs
in response to immune challenge (Table
1). Results for group I Ch animals compared with controls showed a significant increase in
the percentages of Sp185/333+ cells in the axial organ after challenge
(P < 0.0001); however, there were no detectable changes in pharynx, esophagus,
intestine and testes compared with the controls (Figure 9). Similar results were obtained for the group IV N-Ac animals, which had a higher
percentage of Sp185/333+ cells in the axial organ, pharynx and esophagus compared with
controls (P < 0.05; data not shown). There was no change in the percentage of
Sp185/333+ cells in the intestine and testes from animal 21 (group IV). The locations of
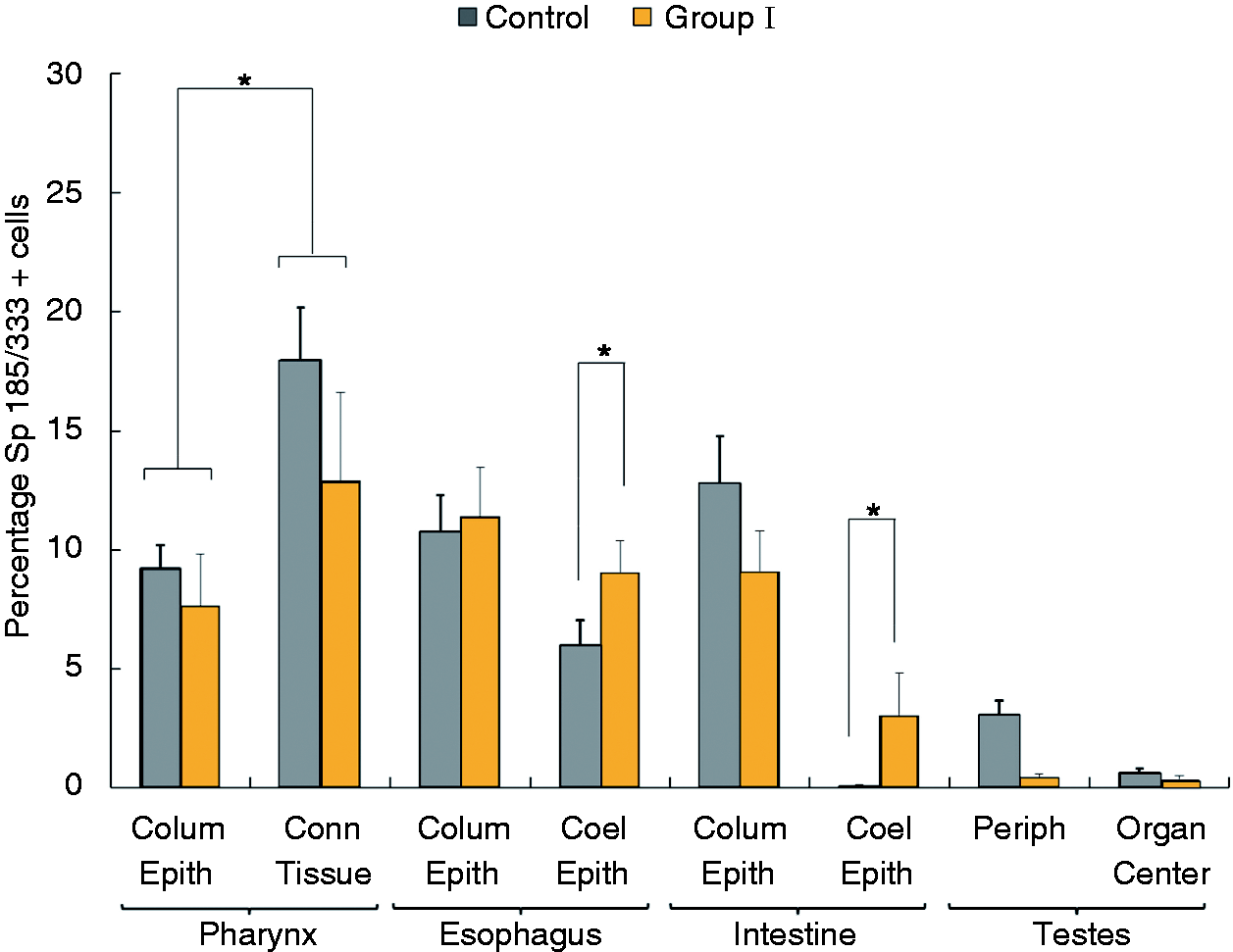
Sp185/333+ cells within some of the organs were also evaluated. Within the esophagus and
intestine there was a higher percentage of Sp185/333+ cells in the coelomic epithelium in
Ch animals compared with controls (P < 0.05), whereas there was no difference in
the percentage of Sp185/333+ cells in the columnar epithelium from Ch animals compared
with controls (Figure 10). When the
location of the Sp185/333+ cells within pharyngeal tissues was evaluated without regard
to the treatment (i.e. Ch animals and controls were analyzed together given that there was no
difference between them), the percentage of Sp185/333+ cells was higher in the connective
tissue at the periphery of the pharynx compared to the columnar epithelium that lines the mouth
(P < 0.05; Figure 10).
There were no differences in the location of Sp185/333+ cells in the testes (center vs
periphery), which did not change with respect to immune challenge. The Sp185/333+ cells
were distributed uniformly throughout the axial organ, including sections from the end regions and
the middle of the organ. In summary, Sp185/333+ cells were observed in all organs. There
was as an overall increase in the percentage of Sp185/333+ cells in axial organ in Ch
animals, whereas the increase in Sp185/333+ cells was variable among the other organs
from Ch animals. Furthermore, the percentages of Sp185/333+ cells within the esophagus
and intestine in Ch animals responding to LPS showed an increase in the percentage of
Sp185/333+ cells within the coelomic epithelium, which was not observed for cells within
the columnar epithelium. The percentage of Sp185/333+coelomocytes and Sp185/333+ cells in the axial
organ increases after immune challenge. The percentages of Sp185/333+ cells versus the
total number of cells were collected for individual animals using IF and grouped by similar
treatment and organ. Coelomocytes (coel) were collected from animals in group I
(n = 3) 0 h before (white bar; Iq) and 24 h after (yellow bar, Ch) LPS injection.
Tissues were collected from group I animals (yellow bars as in Figure 1) after challenge, and were the same animals that donated
coelomocytes (n = 3; n = 2 for testes). Additional organs were
collected from Iq control animals (n = 4; n = 2 for testes; dark
gray bars as in Figure 1). The percentages
of Sp185/333+ coelomocytes increase in challenged animals compared with pre-injection
samples (Iq), and the percentages of Sp185/333+ cells in axial organ (axial o) are also
higher in Ch animals compared with controls (bars with asterisk; P < 0.0001). No
differences in the percentage of Sp185/333+ cells in intestine, testes, pharynx or
esophagus were observed between Ch and control animals. SEM are shown. The percentage of Sp185/333+cells in the coelomic epithelium increases in esophagus
and intestine after immune challenge compared with the controls. The percentages of
Sp185/333+ cells versus the total number of cells were evaluated for individual animals
using IF and grouped by treatment and tissue. Tissues were collected from controls animals (dark
gray bars as in Figure 1;
n = 4; n = 2 for testes) and Ch animals in group I (yellow bars as
in Figure 1; n = 3;
n = 2 for testes) 24 h after LPS injection. Percentages of
Sp185/333+cells in the coelomic epithelium of the esophagus and intestine are higher in
group I Ch animals compared with the same samples in controls (brackets with asterisk;
P < 0.05). The percentage of Sp185/333+cells in the connective tissue
is higher than in the columnar epithelium of the pharynx, when treatment (i.e. Ch vs control) is not
taken into consideration (brackets with asterisks; P < 0.05). SEM are shown.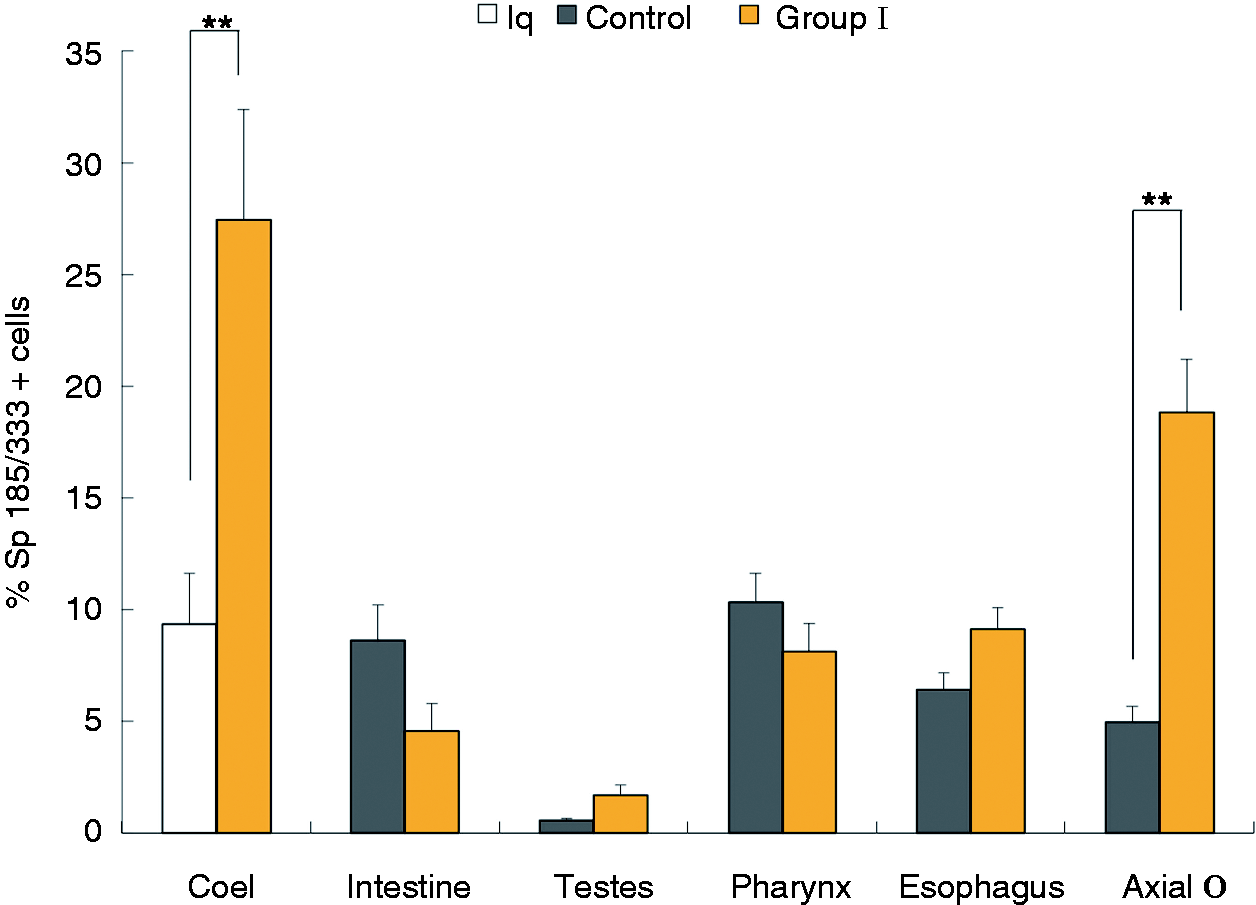
Sp185/333 protein repertoires vary in coelomocytes and organ samples within and among sea urchins
Sp185/333 protein repertoire in coelomocytes is known to be highly variable showing a broad range
of size and pI among different animals.9,15 However, the protein
repertoire has not been investigated in sea urchin organs. Western blot analyses for 12 animals that
included samples from Ch animals (n = 9) and controls (n = 4)
showed that the repertoires of Sp185/333 proteins from organs varied among animals and ranged in
size from < 5 kDa to > 250 kDa (a representative blot is shown in Figure 11). The most common small bands, which may be monomers,
were ∼40 kDa in organs and 52 kDa in coelomocytes. The major bands in coelomocytes ranged from
∼90 kDa to < 250 kDa, which was similar to previous reports.9,15 In immune-challenged animals, additional bands were observed compared with the
Sp185/333 protein bands in the same animals before immune challenge (arrows in Figure 11). The Sp185/333+ banding patterns are different in coelomocytes and organs. Western blot
results are shown for coelomocytes (coel), pharynx, esophagus, intestine, gonad (unknown gender) and
axial organ for animal #15. Coelomocytes were collected pre- and post-immune challenge with LPS, and
organ samples were collected post-immune challenge. Standards are shown on the left. Units are in
kDa.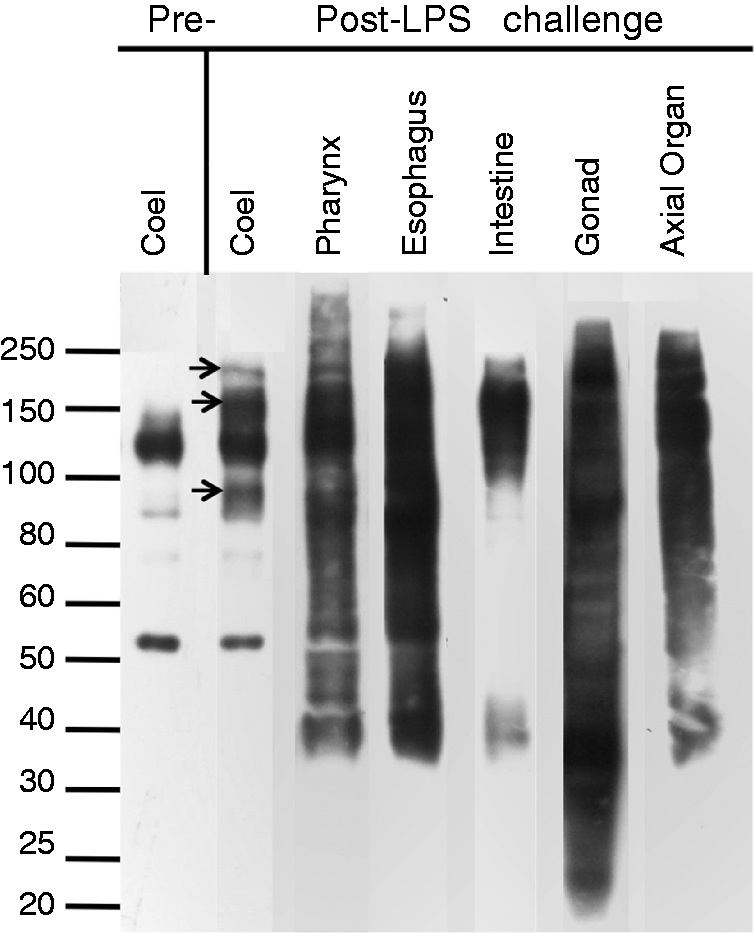
The size range for the major Sp185/333 bands was expanded in the organs compared with the size range in coelomocytes. There was one major pattern of bands in the gonads, which was common among the animals (Figure 11; Table 1).The patterns of Sp185/333 bands in pharynx and esophagus were very similar within and among different animals; however, the patterns in the intestine varied in the number and size of bands among different animals. The Sp185/333 band sizes in the axial organ were markedly similar to both pharynx and esophagus both within and among animals. However, in general, there were fewer bands in the axial organ, which were similar to the overall pattern of bands in coelomocytes after activation with LPS. There were no obvious changes in patterns of Sp185/333 bands in organs from Ch animals after LPS challenge compared with the same organs from controls (not shown). Overall, there was diversity among Sp185/333 protein repertoires among the different organs and coelomocytes within and among animals. In immune-challenged animals there was an expansion in the sizes of the Sp185/333 bands in coelomocytes compared with coelomocytes from unchallenged animals, but there were no major shifts in the repertoire of these proteins in organs from Ch versus controls.
Discussion
We show here that the diverse family of Sp185/333 immune genes and the proteins
that they encode are expressed by cells dispersed in the major organs of S.
purpuratus, in addition to expression by the coelomocytes that has been reported
previously.9,15 The Sp185/333 transcript level and
Sp185/333 protein content in coelomocytes increases after immune challenge, and these changes are
also observed in the intestine and the axial organ (Figure 12). While the Sp185/333 protein content increases
significantly after LPS injection in pharynx and esophagus, there is no change in the level of
Sp185/333 transcripts or the proportion of Sp185/333+ cells in these
organs (Figure 12). This suggests that
rather than an influx of Sp185/333+cells to the pharynx and esophagus, there may be an
increase in protein production from pre-existing Sp185/333 transcripts. Finally, an
increase in the proportion of Sp185/333+ cells was only observed in the axial organ after
immune challenge (Figure 12). The increase
in Sp185/333 transcript level, protein content and the proportion of
Sp185/333+ cells after LPS injection in the axial organ suggests that either existing
Sp185/333-negative (Sp185/333−) cells may be induced to produce the proteins and/or there
may be an influx of Sp185/333+ cells to the axial organ from elsewhere. In addition to
the assessment of Sp185/333 transcript expression and Sp185/333 protein content,
the range of commonly observed Sp185/333 protein sizes in the organs (< 5 kDa to >250 kDa) is
broader than that detected for the most common Sp185/333 proteins in coelomocytes (∼40–250 kDa).
This suggests that these proteins may either be expressed by non-coelomocytes or that the proteins
are multimerized and/or processed differently in organs irrespective of the cell type that expresses
them. Post-translational processing may be the major mechanism for generating variations in the
Sp185/333 proteins because the size range of mRNAs is very similar among organs and coelomocytes
(Supplementary File 1). In general, the Sp185/333 proteins are present in the major organs of the
sea urchin and this, perhaps, extends throughout all of the tissues. The axial organ and intestine respond to LPS challenge. The internal anatomy of an adult sea
urchin that has been cut in half and opened is shown on the left. The table to the right indicates
the responses to LPS in Ch animals. The axial organ shows increases in Sp185/333
mRNA, Sp185/333 protein and Sp185/333+ cells after challenge with LPS. The intestine
shows increases in Sp185/333mRNA and Sp185/333 protein. Responses by the pharynx
and esophagus are consistent with the production of additional Sp185/333 protein, perhaps from
existing mRNA. Neither testis nor ovary show changes in response to LPS. +: samples that show
significantly increased levels of Sp185/333 mRNA, Sp185/333 protein or
Sp185/333+ cells compared with controls; −: samples that did not show differences from
the controls.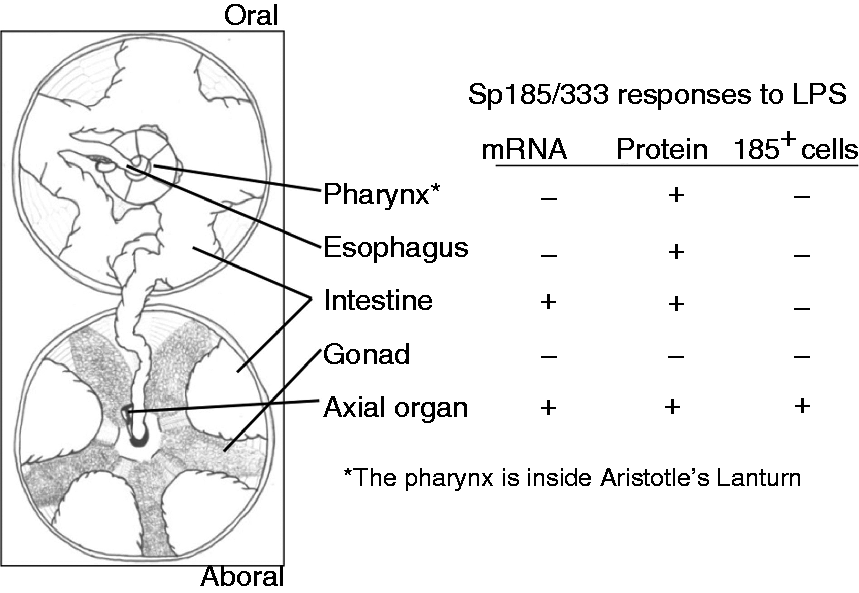
Sp185/333 in coelomocytes
In response to immune challenge, almost twice as many coelomocytes appear in the coelomic cavity compared with pre-challenge for Iq sea urchins. 15 This includes a significant increase in the total number of Sp185/333+ cells, which are composed of small phagocytes and polygonal phagocytes. The additional small phagocytes that appear post-challenge include cells that are Sp185/333− and Sp185/333+, suggesting that both appear in the coelomic fluid as a result of either proliferation or release of pre-existing marginated cells. 15 However, there is no overall change in the number of polygonal phagocytes in the coelomic fluid after immune challenge, although there is an increase in the number of this cell type that is Sp185/333+. This indicates that some of the existing Sp185/333− polygonal phagocytes may be induced to produce Sp185/333 proteins. Our results for coelomocytes are in agreement with these previously published data. 15
Sp185/333 in the intestine
He185/333 proteins have been isolated from in another sea urchin species, Heliocidaris erythrogramma, where they are present in cells associated with the intestine and are localized to the coelomic epithelium in contact with the coelomic fluid. 16 This is the same location as reported here for Sp185/333+ cells in S. purpuratus, which are also associated with the coelomic epithelium that covers the outer surface of the intestine. However, Sp185/333+ cells present in the columnar epithelium of the intestine in S. purpuratus were not reported in H. erythrogramma. 16 The intestinal localization of the Sp185/333+ cells within the columnar epithelium is strikingly similar to the variable region-containing chitin-binding proteins (VCBPs) expressed in intestine-associated cells in amphioxus, Branchiostoma floridae, and in the solitary tunicate, Ciona intestinalis.29,30 It is noteworthy that both the Sp185/333 and the VCBP gene families have been described as highly diverse,12,13,29–35 that the VCBPs bind to bacteria and augment phagocytosis by hemocytes in C. intestinalis 30 and that a recombinant Sp185/333 protein binds to marine Gram-negative bacteria (Lun, Schrankel, Sacchi and Smith, unpublished). The Sp185/333+ cells in the columnar epithelium of the intestine may be intestine cells, while the cells associated with the coelomic epithelium surrounding the intestine in contact with the coelomic fluid may be coelomocytes. The increased Sp185/333 transcript level and elevated Sp185/333 protein content in the intestine in response to immune challenge may be an indication that these cells are involved in immune functions or specialized intestine-associated immunity and that they may all be coelomocytes. Because the GI tract contains ingested and resident/commensal microbes, an active mucosal immune response as predicted from the large numbers of NLR gene models in the genome would be required to control or combat these microbes.2,36 This mucosal immune system would likely include cells expressing proteins with antimicrobial function. Overall, the Sp185/333 protein expression pattern in the intestine is similar to the He185/333 proteins in the coelomic epithelium, as well as to the staining pattern of VCBPs in the GI tract of a urochordate and a cephalochordate. This suggests that the Sp185/333 proteins may have immune effector function(s) in organs, either through parenchymal cells of the organs or wandering coelomocytes.
Sp185/333 in the axial organ
The axial organ lies at the central axis of the spheroid echinoid body, and is positioned at the conjunction of the perivisceral coelom, and the water vascular and haemal systems.25,37 It is suspended in these cavities through attachment to the stone canal near the esophagus, anus and madreporite. 38 It has been described as a porous complex with sinuses or haemal spaces called lacunae and canaliculi. 37 Coelomocytes are described as being present in the haemal spaces of the axial organ,25,37 but this has not been verified using modern molecular or cellular methods owing to the unavailability of markers that are specific for coelomocytes. Proposed functions of the axial organ include that of an immune organ,39–42 or the site of coelomocyte and cell production 43 or cell degradation. 44 It has also been described as an excretory organ,45,46 a contractile vessel, 37 a primitive heart47,48 and a secretory gland.48,49 However, owing to the lack of convincing experimental evidence, the functions of this organ remain unknown.
The axial organ is the only organ that shows significant increases in all parameters that were measured: Sp185/333 transcript levels, Sp185/333 protein content, and the proportion of Sp185/333+ cells in response to immune challenge (Figure 12). Although, we could not determine definitively which cell type(s) express the Sp185/333 proteins in any of the organs, there are several possible explanations for the increased proportion of Sp185/333+ cells in the axial organ after immune challenge: (i) coelomocytes and/or parenchymal cells present in the axial organ may respond to immune challenge by inducing Sp185/333 gene expression and protein production; (ii) Sp185/333+ cells in the axial organ may proliferate; (iii) Sp185/333+ cells may enter the organ from the coelomic fluid. Any combination of these possibilities may also occur. The axial organ may be a site of coelomocyte proliferation, including the subsets of phagocytes that express the Sp185/333 genes and produce the encoded proteins. This hypothesis is consistent with the expression of SpTie1/2 in the axial organ, which increases in response to immune challenge and is a homologue of vertebrate Tie genes that are involved in hematopoiesis. 50 Recent evidence suggests that cells proliferate in the axial organ, coelomic epithelium and Tiedemann bodies of the sea star, Asterias rubens, in response to immune challenge. 51 If axial organ parenchymal cells express the Sp185/333 proteins, this may indicate that the axial organ plays a role in immunity for the animal, perhaps similar to that of the fat body in insects. 52 However, if the accumulation of Sp185/333+ cells is a result of cells migrating to the organ, these may be senescent cells that accumulate in the axial organ to be removed from the body either via the stone canal and madreporite, by transfer to the rectal tissues and removal through the anus, and/or otherwise undergo apoptosis. While we present here the first characterization of Sp185/333 protein expression in the organs of the purple sea urchin, the identity of the Sp185/333+ cells in tissues remains speculative. However, the morphology of Sp185/333+ cells in the axial organ is very similar to small phagocytes, and we speculate that this organ may be a source of Sp185/333+ cells and coelomocytes in general.
Sp185/333 in the ovaries
Apart from the axial organ, the identification of the Sp185/333+ cells associated with pharynx, esophagus, intestine and testes could not be established with certainty based on cell morphology. However, within the ovary, the pattern of the Sp185/333 proteins in oocytes and mature eggs is very similar to the pattern of proteins associated with cortical granules (Gary Wessel, personal communication).26,28 Cortical granules in oocytes are scattered throughout the cytoplasm, and in the mature egg they are localized to the cortex near the plasma membrane. The fertilization reaction induces the granules to undergo exocytosis and release their contents into the perivitteline space, which results in the proteolysis of the sperm receptor, 53 transformation of the outer soft vitelline envelope layer into a thickened fertilization envelope 54 and blocks polyspermy. It is noteworthy that a sea urchin integrin is also localized to cortical granules with the same pattern as described here for Sp185/333 proteins. 55 Upon fertilization, the integrin is integrated into the fertilization envelope and putative interaction with the Sp185/333 proteins, perhaps via the RGD or other conserved sequence for integrin binding, would position the Sp185/333 proteins on the surface of the fertilized egg. It is therefore intriguing to speculate that the Sp185/333 proteins may function to protect the egg from bacterial colonization or other foreign invasion.
There are a number of reports and reviews of echinoid diseases (e.g. Jangoux 56 ), of which most focus on skin infections, their outcomes and their effect on sea urchin aquaculture. Spotted gonad disease, reported for Strongylocentrotus intermedius, shows an accumulation of red spherule cells (and likely other types of coelomocytes) in the gonad, which surround matacercaria of parasitic digenetic trematode worms.57,58 Red spherule cells are sometimes released with eggs during spawning (LCS, personal observation) and these cells may infiltrate gonadal tissues in response to an infection. Although there is no description of how microbes and parasites might enter the gonads of otherwise healthy animals, migration down the gonopore into the gonadal tissue is likely. The expression of Sp185/333 proteins by ovarian follicular cells and perhaps by phagocytes in ovary and testicular tissue may be important for deterring or blocking potential infections to these organs.
Conclusions
The evidence presented in this study suggests that cells expressing Sp185/333 proteins may not be confined to coelomocytes in the coelomic fluid of the purple sea urchin. The Sp185/333 transcripts and proteins are present in cells in the major organs and tissues throughout the body, including the axial organ, pharynx, esophagus, intestine and gonads. It is feasible that the Sp185/333+ cells in the organs are wandering phagocytes and that they may have a variety of functions in the organs, including immune defense. Highly diverse arrays of 185/333 proteins are expressed by two species of sea urchins (S. purpuratus and H. erythrogramma), and the genes have been identified in the genomes of three additional species, Strongylocentrotus franciscanus, Allocentrotus fragilis 23 and Lytechinus pictus (Buckley and Smith, unpublished). The extraordinary sequence diversity of the encoded proteins is likely a reflection of the arms race between host and pathogen.59,60 Sea urchins are in constant contact with microbes that share their seawater and ocean substrate habitat. 61 Infection from this constant exposure to microbes is likely held in check by a variety of immune mechanisms, including the activities of the coelomocytes and the sequence diversity that is present a variety of the sea urchin immune gene families,1,2,4,5 including the Sp185/333 genes. 24
Footnotes
Funding
This work was supported by the US National Science Foundation grants MCB 04-24235 and MCB 07-44999 to LCS. Travel funding and educational support to AJM was provided by the Department of Biological Sciences and Columbian College of Arts and Sciences, George Washington University.
Acknowledgements
The authors wish to thank Dr Katherine Buckley, Rebecca Easley, Dr Young-Ok Kim, Elizabeth Hoye and Praveen Savalgi for assistance with tissue and coelomocyte collections. Drs David Raftos, John Henson, Robert Donaldson, Ioannis Eleftherianos, Ken Brown and two anonymous reviewers provided very helpful improvements to the manuscript. The authors thank Drs Ken Brown and Gary Wessel for suggestions on the morphology and staining pattern of the Sp185/333 proteins in ovaries. The authors also thank Dr Anastas Popratiloff for assistance with confocal imaging.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.