
Abstract
Scavenger receptor A (Sra), also known as macrophage scavenger receptor 1 (Msr1), is a surface glycoprotein preferentially present in macrophages that plays a primary role in innate immunity. Previous studies have shown that Sra is a modifier gene for the response to bacterial LPS in mice at the level of IL-10 production, in particular. In the present study, we found that Sra(−/−) mice are more resistant to septic shock induced by cecal ligation and puncture than wild-type C57BL/6 J (B6) mice. In addition, Sra(−/−) mice displayed initial elevated high density lipoprotein (HDL) circulating levels. Naïve peritoneal macrophages (PMϕs) were isolated from Sra(−/−) mice to understand the possible protective mechanism. Incubation of these cells with LPS was found to modulate TLR4 signaling, leading to a reduction in IL-10 and IL-6 mRNA levels, but not TNF-α expression, at low concentrations of LPS in comparison with PMϕs isolated from B6 mice. No differences were found in LPS binding between PMϕs derived from Sra(−/−) or B6 mice. The lack of Sra binding to LPS was confirmed after transfection of Chinese hamster ovary (CHO) cells with the Sra gene. The contribution of Sra to the outcome of sepsis may be a combination of changes in TLR4 signaling pathway and elevated levels of HDL in circulation, but also LPS toxicity.
Introduction
The innate immune system is the first line of defense against invading pathogens, which is orchestrated by a network of various cell-types, including macrophages (Mϕs), dendritic cells (DCs), polymorphic nuclear leukocytes (PMNL), and monocytes. These cells express an array of molecules involved in the recognition of pathogens or their products that distinguish specific molecular features common to these microorganisms, broadening their scope and reaction capacity. These molecules have been coined pattern recognition receptors (PRRs) and their ligands coined pathogen associated molecule patterns (PAMPs). The recognition of PAMPs by PRRs triggers a variety of responses directed at destroying, neutralizing, and clearing pathogens in order to resolve the infection. 1 It has become clear that PRRs are also involved in the response to injury, and the clearance of apoptotic cells and cellular debris secondary to tissue necrosis. Therefore, PRRs seem to play a wide role in many pathological conditions. PRRs can be localized in the extracellular environment as soluble molecules (e.g. mannan binding lectin), associated with the plasma membrane (e.g. scavenger receptors, C-type lectins) or combined between the cell surface or subcellular compartment, as is the case with the TLR family. 1
Scavenger receptors, as parts of PRRs, comprise a large family of cell surface molecules with the common characteristic of recognizing complex macromolecules. They are classified into two subgroups: one that binds modified lipoproteins and polyanionic ligands (class A) and another that recognizes Ig-like domains (class B). They have also been grouped according to their molecular structure, such as by the presence of collagenous domains that characterize scavenger receptor A (Sra) and macrophage receptor with collagenous domain (MARCO). Sra, also known as macrophage scavenger receptor 1 (Msr1) or CD204, is a cell surface glycoprotein primarily expressed on myeloid cells.2–4 Sra is present in three isoform products of alternative splicing, all containing exons 1–8, but differing in the presence of exons 10 and 11 (SraI), exons 9 (SraII), and 10 (SraIII). SraI and II have been detected in cells, whereas SraIII is of questionable expression. 5 The typical collagenous domain, which is considered the binding site for multiple ligands, is present in all isoforms, as well as the coiled-coil region that is responsible for trimerization, whereas a cystein-rich domain is only present in SraI.2,4 These receptors are type II glycoproteins with a very short cytosolic tail at the N-terminus of the molecule. Sra is expressed in Mϕs, but not in PMNL or monocytes.3,4
Sra was initially discovered as a binding protein for modified lipoproteins [e.g. acetylated low density lipoprotein (LDL), oxidized LDL]. It is particularly present in foam cells and its role in arteriosclerosis has been well recognized.3,4,6,7 Sra has also been implicated in the development of Mϕ apoptosis under endoplasmic reticulum stress.8,9 Sra is also up-regulated during Mϕ differentiation. 10 It plays a significant role in phagocytosis of bacteria11–14 and apoptotic cells. 15 Likewise, Sra is involved in cell adhesion.16,17 Inbred mouse strains lacking Sra (Msr1) showed increased susceptibility to Listeria monocytogenes and herpes simplex virus, 18 malaria, 19 and Neisseria meningitides.13,20 Similarly, Sra(−/−) mice were reported to be more susceptible to LPS if they were primed by injection of bacillus Calmette-Guerin 21 or in absence of priming.22–24 In contrast, other studies have shown that Sra(−/−) mice are protected from a LPS challenge. 25
We identified Sra (Msr1) as a candidate gene in genetic screening for phenotypic differences in the inflammatory response between mouse strains, which could be used as a potential predictor of the incidence of sepsis. 26 Sepsis is a serious health problem with an incidence of 750,000 cases per year in the USA, a mortality rate of 30–50%, and an annual healthcare cost of more than US$17 billion. 27 Sra (Msr1) was recognized based on differences in LPS-induced plasma levels of IL-10 between C57BL/6 J (B6) and A/J mice. 28 Polymorphisms in this gene, which have been previously described, 29 were correlated with differences in LPS-induced IL-10 expression levels in various mouse strains. 28 We found that LPS-induced IL-10 levels were depressed in Sra(−/−) mice as opposed to wild-type B6 mice. 28 We extended our prior studies to further investigate the role of Sra in the inflammatory process during sepsis. In this study, we observed that Sra is a modifier of the TLR4 signal transduction pathway, which was independent of direct LPS binding to the scavenger receptor. Moreover, we found that Sra(−/−) mice were more resistant to septic shock, which is correlated to initial elevated high density lipoprotein (HDL) levels in the transgenic mouse.
Materials and methods
Mice
Sra(−/−) mice, which were backcrossed over seven generations to B6 mice, 30 were originally obtained as a gift from Dr Mason Freeman. These animals were bred in our colony at University of California San Diego and were certified to be Helicobacter-free. In addition, a colony of B6 mice was maintained in the same environment as Sra(−/−) mice and used as a wild-type control. Both strains of mice were fed with the same regular chow diet. All procedures were performed in accordance with the Guidelines for the Care and Use of Laboratory Animals of the National Institutes of Health, and were approved by the Institutional Animal Care and Use Committee of the University of California, San Diego School of Medicine.
Experimental animal models
Male mice (8–9-wks-old) were fasted for 16 h before any intervention, which was always performed during the morning between 10:00 and 11:30. Male mice were anesthetized with isoflurane via a vaporizer at 1.5–2.5 minimum alveolar concentrations (MAC). Under sterile conditions, a 2-cm incision was made in the lower abdominal region and the cecum was exposed. The distal portion of the cecum was ligated 1.5 cm from the end with a 4-0 silk suture and punctured once with either an 18- or 16-gauge needle. The cecum was replaced in the peritoneal cavity and squeezed to place a small portion of its content (bacteria and feces) into the peritoneum. Then, the peritoneal wall and skin were closed with double sutures. Mice were resuscitated with a 1-ml subcutaneous injection of sterile saline (0.9%). After the procedure, mice had access to water and food ad libitum.31,32
Isolation of peritoneal Mϕ
Resident naïve peritoneal Mϕs (PMϕs) were isolated by peritoneal cavity lavage as described previously. 33 Male mice (8–9-wks-old) were fasted for 16 h before the procedure and euthanized under anesthesia with isoflurane via a vaporizer at 1.5–2.5 MAC during the morning. Under sterile conditions, a 16-gauge catheter was inserted into the peritoneum. Cold RPMI 1640 medium supplemented with 1% heparin (20 U/ml), 1% penicillin (100 U/ml), and 1% streptomycin (100 U/ml) was introduced (5 ml) via the catheter into the peritoneum. The abdomen was gently massaged and the lavage was withdrawn through the catheter. The peritoneal lavage was centrifuged at 800 rpm (115 × g) for 10 min. The pellet containing the cells was suspended in 1 ml of RPMI 1640 medium supplemented with 10% FBS, 1% penicillin, and 1% streptomycin (approximately 1 × 106 PMϕ/mouse). Cell suspension was seeded on a 12-well plate at a concentration of 5 × 105 cells per well. Cells were allowed to attach for 1 h at 37°C in a CO2 incubator. Non-adherent cells were removed and fresh medium was added. Cells were incubated for an additional 16 h and then stimulated (or not) with LPS (1 or 100 ng/ml Escherichia coli 026:B6; Sigma-Aldrich, St Louis, MO, USA) in RPMI 1640 medium supplemented with 10% FBS for 3 h at 37°C. Cellular viability was determined by the trypan-blue exclusion test (90% viability was considered acceptable). Purity of the cell population was determined by staining with F4/80 Ab. A 90–95% population of Mϕ was considered acceptable. PMϕs isolated by this procedure did not appear to be activated by the lack of cytokine gene expression in absence of LPS stimulation.
Cloning of SraI and II and cell transfection
SraI and SraII were cloned from a cDNA library prepared using PolyA selected RNA obtained from PMϕ isolated from A/J or B6 mice. Sra genes were cloned by PCR using platinum pfx polymerase (Invitrogen, Carlsbad, CA, USA) and the following primers: 5′ UTR (CGA
Immunostaining
Transfected cells were analyzed for Sra expression by fluorescent microscopy using a rabbit polyclonal Ab against all Sra isoforms. 28 CHO-K1 cells were incubated with 10% fetal sheep serum (FSS) in PBS for 30 min at 25°C to block non-specific binding. Cells were incubated with rabbit anti-Sra Ab (1/200) in PBS-5% FSS for 1 h at 25°C. Cells were washed three times with PBS and incubated with AlexaFlour 488-conjugated anti-Rabbit Ab in PBS-5% FSS for 40 min at 25°C. Cells were then washed 3 × w/PBS, fixed with 4% paraformaldehyde for 10 min, DAPI stained, then washed again 3 × w/PBS. Alternatively, cells were incubated with AlexaFluor488-conjugated-Ac-LDL (1/50) or LPS-Biotin (visualized by AlexaFlour 594-conjugated streptavidin) following the same protocol as described above.
Measurement of mRNA levels by quantitative real time PCR
PMϕs were washed twice with PBS and homogenized in TRIzol reagent (Invitrogen) and total RNA was isolated, as recommended by the manufacturer. Total RNA was treated with DNase I (DNA-free kit, Ambion, Austin, TX, USA) and reverse transcribed to cDNA using the High Capacity Reverse Transcription Kit (Applied Biosystems, Foster City, CA, USA). Newly synthesized cDNA was stored at −20°C. Samples of cDNA were amplified by real time (RT) PCR in a 7500 Fast Real-Time PCR System (Applied Biosystems) using QuantiTect SYBR Green PCR kit (Qiagen, Valencia, CA, USA) with the following QuantiTect validated primer sets (Qiagen): TNF-α (QT00104006), IL-6 (QT00098875), IL-10 (QT00106169), CD14 (QT00246190), and TLR4 (QT00259042). Standards corresponding to each target gene were added in each PCR reaction. The results for each sample were normalized by copy number of GAPDH (QT01658692) used as a marker of cDNA inputs. All results were expressed as copy numbers of target gene per copy numbers of GAPDH.
Flow cytometry analysis
PMϕs were gently scraped, centrifuged, and re-suspended in FACS staining buffer (FSB, DPBS without Ca2+/Mg2+ supplemented with 0.5% BSA). PMϕs were then incubated with rat anti-mouse CD16/CD32 (mouse Fc block, BD Pharmingen, San Diego, CA, USA) for 15 min. In some experiments, PMϕs were incubated with APC-conjugated anti-F4/80 Abs (eBioscience, San Diego, CA, USA) and biotinylated LPS for 30 min followed by AlexaFluor488-conjugated streptavidin (BD Pharmingen). In other experiments, PMϕs were stained for 30 min with a combination of FITC-conjugated anti-CD14 (eBioscience) and Allophycocyanin (APC)-conjugated anti-TLR4/MD2 Abs (eBioscience). PMϕs were also incubated with biotin-conjugated LPS (10 µg/ml for 30 min) followed by AlexaFluor488-conjugated streptavidin. PMϕs were then washed and re-suspended in FSB. Fluorescence was acquired using a BD FACSCanto II and analyzed by FlowJo software (Tree Star, Ashland, OR, USA).
Cholesterol analysis
Male mice (8–9-wks-old) were nil per os (NPO) for 16 h before sample collection. Mice were euthanized under isofluorane anesthesia, and approximately 500 µl of blood was drawn from mice via cardiac puncture using a 16-gauge needle and placed in EDTA-coated tubes on ice. Tubes were spun down at 5000 g for 5 min and plasma was collected from the supernatant, placed into fresh tubes, and stored at −20°C until further analysis. Cholesterol levels were performed by the hematology core facility at the University of California San Diego.
Statistical analysis
All data were analyzed using GraphPad Prism software (GraphPad Prism Software, San Diego, CA, USA). Significance was analyzed using a Student’s t-test or one-way ANOVA followed by Newman-Keuls Multiple Comparison Test. A P value of <0.05 was considered statistically significant. Statistical significance was analyzed by log-rank test for comparison of mortality rates.
Results
Sra(−/−) mice are more resistant to septic shock induced by cecal ligation and puncture
Sra1 (Msr1) was mapped as a modifier gene of the inflammatory response induced by LPS.
28
In the present study, we investigated the response of Sra(−/−) mice to septic shock. We used cecal ligation and puncture (CLP), which is the gold standard model to study sepsis in rodents, because it mimics many features of the condition in humans. Male Sra(−/−) or B6 mice were subjected to CLP using a single perforation with an 18- or 16-guage needle that resulted in higher mortality rates depending on the increase in puncture size. Sra(−/−) mice displayed better survival rates (56%) than B6 mice (10%) after CLP with 18-guage perforation, which was statistically significant (P < 0.05) (Figure 1A). These results were corroborated with a more severe injury (CLP with 16-guage perforation), displaying 30% survival for Sra(−/−) mice and 14% for B6 mice (P < 0.05) (Figure 1B). B6 mice reached 50% mortality within 38 h and 48 h after 16 - or 18-guage perforation, respectively. In contrast, Sra(−/−) mice displayed 50% mortality within 54 h after CLP with 16-guage perforation and did not reach 50% mortality after CLP with 18-guage puncture (Figure 1).
Survival of Sra(−/−) and B6 mice after CLP. Male Sra(−/−) or B6 mice were NPO for 16 h before the procedure. Anesthetized mice were subjected to CLP (1.5-cm ligation) and puncture with 18 - or 16-guage needle. Survival was monitored continuously for 120 h (18-gauage) or 72 h (16-guage) after CLP. Statistical significance was analyzed by log-rank test. (A) CLP (18-guage), Sra(−/−) n = 10, B6 n = 20, *P < 0.05 Sra(−/−) vs. B6. (B) CLP (16-guage), Sra(−/−) n = 20, B6 n = 20, *P < 0.05 Sra(−/−) vs. B6.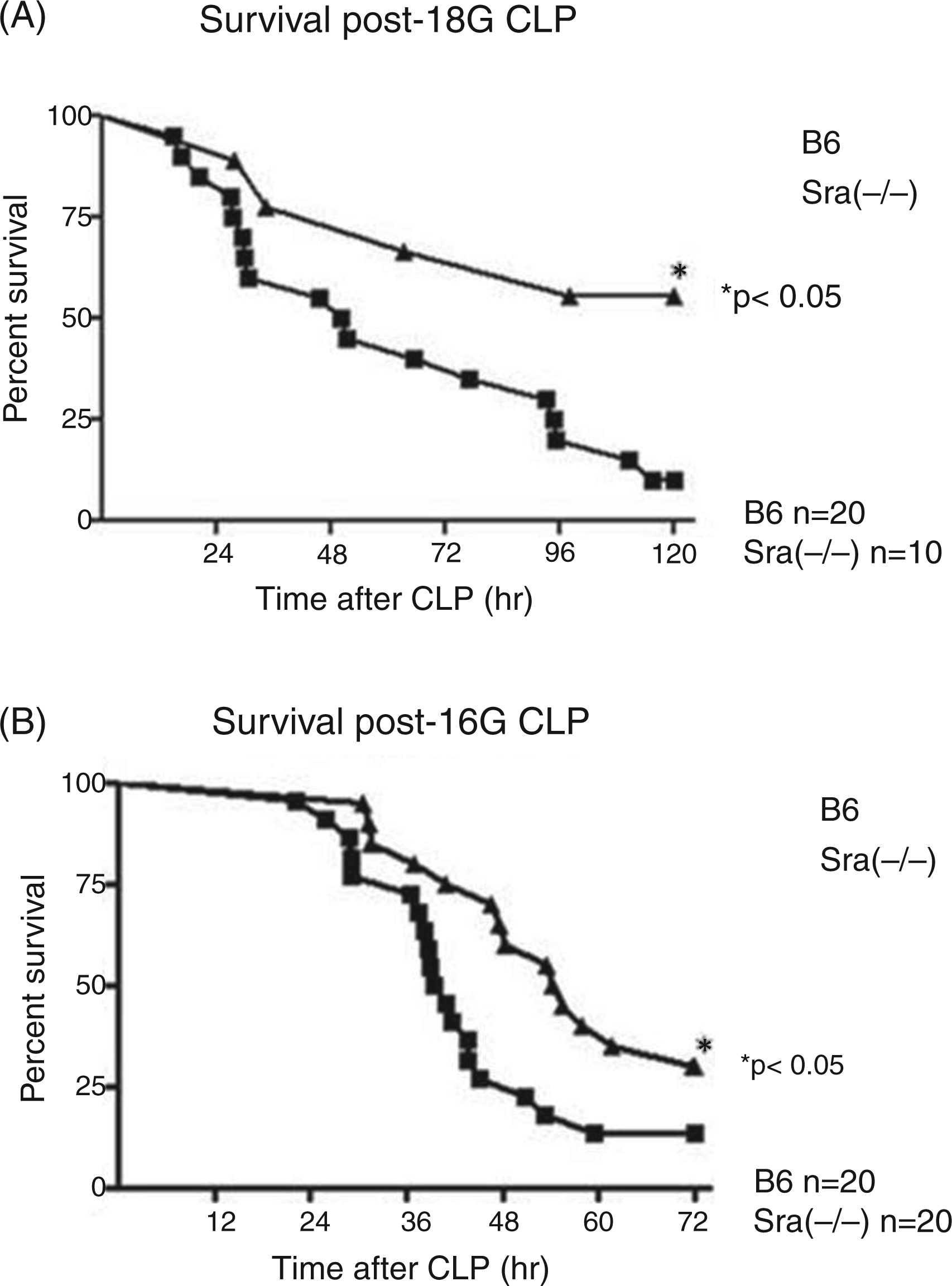
Cytokine expression is altered in PMϕs derived from Sra (−/−) mice
The potential role of Sra1 in the response to LPS was investigated in PMϕs isolated from naïve Sra(−/−) or wild-type (B6) mice. These primary cells were incubated in culture conditions with two concentrations of LPS (1 ng/ml or 100 ng/ml). The lower LPS concentration activates Mϕs exclusively via CD14, which is the major LPS binding site on Mϕs, whereas the higher concentration activates Mϕs independently of the presence of CD14.
34
LPS-induced cytokine expression (mRNA levels) was measured by quantitative RT-PCR (qRT-PCR), which provided a rapid and accurate assessment of gene expression. LPS-induced TNF-α mRNA levels were no different between PMϕs isolated from Sra(−/−) and B6 mice at both LPS concentrations (Figure 2A, B). In contrast, IL-10 expression was significantly reduced in PMϕs derived from Sra(−/−) mice in comparison with cells from B6 mice at the low LPS concentration (Figure 2C), but not at high LPS concentrations (Figure 2D). Similar observations to IL-10 were made for IL-6 expression (Figure 2E, F). These observations suggest that Sra modulates TLR4 signaling, depending on the concentration of LPS. We also investigated the response of PMϕs isolated from Sra(−/−) or B6 mice to a different ligand, zymosan, which activates Mϕs via TLR2 or dectin. Expression of both TNF-α and IL-10 was equally elevated in PMϕs isolated from Sra(−/−) and B6 mice (data not shown).
LPS-induced cytokine profile in PMϕs isolated from Sra(−/−) or B6 mice. Naïve PMϕs were isolated from Sra(−/−) or B6 mice by peritoneal lavage in the absence of recruiting agents, as described in the ‘Materials and methods’. Cells were incubated (or not) with LPS (1 ng/ml or 100 ng/ml as indicated) for 3 h at 37°C in RPMI 1640 medium containing FBS (10%). At the end of the incubation time, RNA was then isolated, DNase treated, and reverse-transcribed to cDNA. TNF-α, IL-10, and IL-6 mRNA levels were determined by qRT-PCR using the standard curve method. The housekeeping gene GAPDH was used to normalize the data. Results are expressed as average ± SE (n = 4–5). Significance was analyzed by student’s t-test between cells under each condition isolated from Sra(−/−) or B6 mice. P < 0.02 for IL-10 Sra(−/−) vs. B6. P < 0.02 for IL-6 Sra(−/−) vs. B6.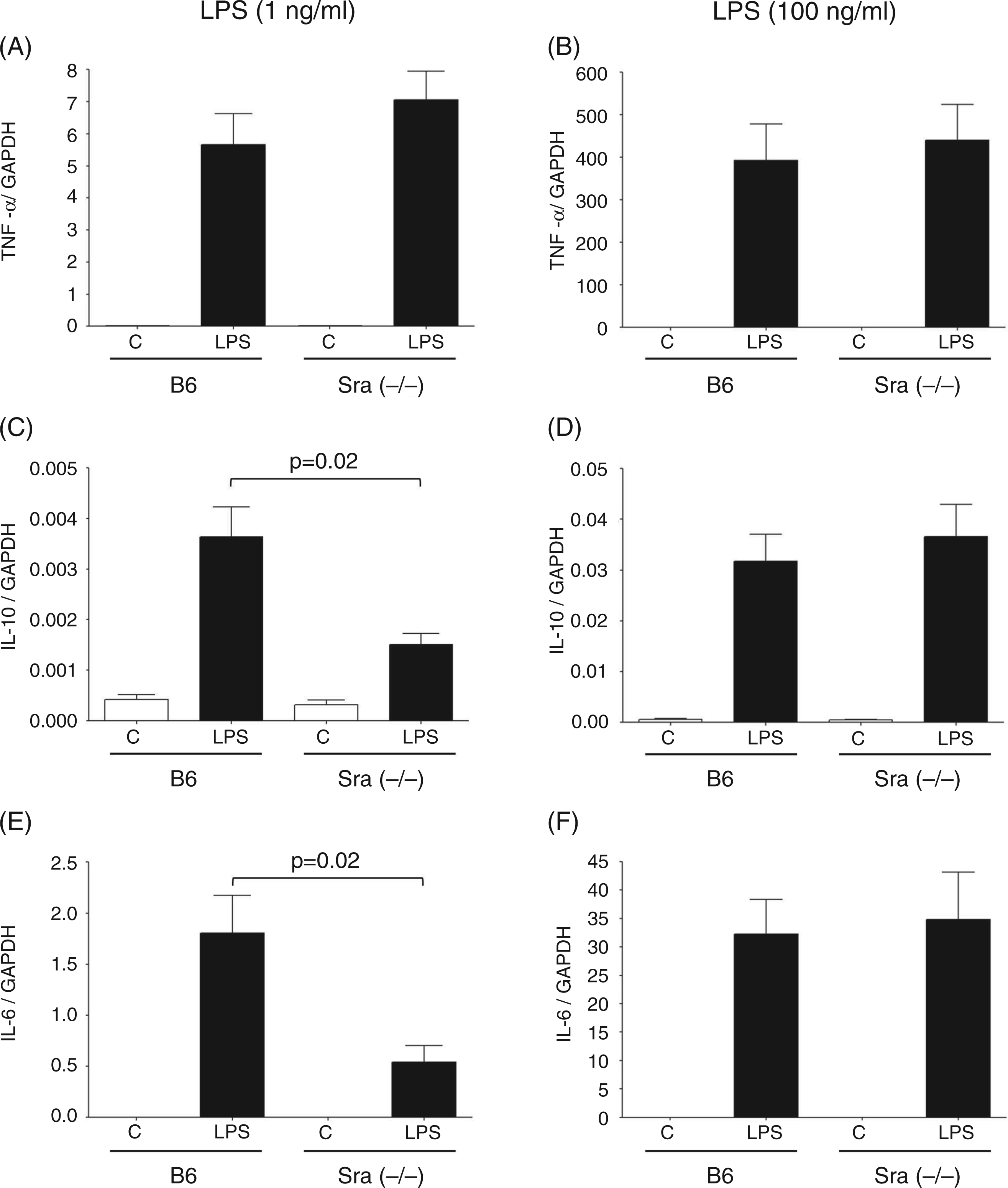
TLR4 and CD14 expression is similar between PMϕs derived from Sra (−/−) and B6 mice
As the preceding observations suggest that the TLR4 signaling pathway is altered in the absence of Sra at low concentrations of LPS, we evaluated the expression of TLR4 and CD14 in PMϕs isolated from Sra(−/−) mice or B6 mice after incubation with LPS (1 ng/ml) in culture. Surface levels for both TLR4 and CD14 in resting cells were slightly reduced in PMϕs isolated from Sra(−/−) as opposed to B6 mice (Figure 3A). The expression of both receptors (mRNA levels) was equally increased upon LPS stimulation in PMϕs isolated from Sra(−/−) and B6 mice (Figure 3B). Similarly, cell surface levels of CD14 were similar between PMϕs isolated from Sra(−/−) and B6 mice, even after LPS (1 ng/ml) stimulation in culture. A small reduction in TLR4/MD2 surface levels between PMϕs isolated from B6 or Sra(−/−) mice after incubation with LPS was observed (Figure 3C).
Comparison of CD14 and TLR4 mRNA,and surface levels on Sra(−/−) and B6-derived PMϕs before and after incubation with LPS. Naïve PMϕs were isolated from Sra(−/−) or B6 mice by peritoneal lavage in the absence of recruiting agents, as described in the ‘Materials and methods’. (A) PMϕs were stained with a combination of FITC-conjugated anti-CD14 and APC-conjugated anti-TLR4/MD2 Abs. Fluorescence was acquired using a BD FACSCanto II flow cytometer and analyzed by FlowJo software. (B) Cells were incubated (or not) with LPS (1 ng/ml) for 3 h at 37°C in RPMI 1640 medium containing FBS (10%). RNA was then isolated, DNase treated, and reverse-transcribed to cDNA. CD14 and TLR4 mRNA levels were determined by qRT-PCR using the standard curve method. The housekeeping gene GAPDH was used to normalize the data. Results are expressed as average ± SE (n = 4–5). Significance was analyzed by student’s t-test between cells under each condition isolated from Sra(−/−) or B6 mice. (C) LPS-treated PMϕs were stained with a combination of FITC-conjugated anti-CD14 and APC-conjugated anti-TLR4/MD2 Abs. Fluorescence was acquired using a BD FACSCanto II flow cytometer and analyzed by FlowJo software. Representative histogram plots are shown with filled grey histograms for control PMϕs (untreated) and open black histograms for LPS-treated PMϕs. Dashed histograms represent CD14 or TLR4 corresponding isotype control Abs.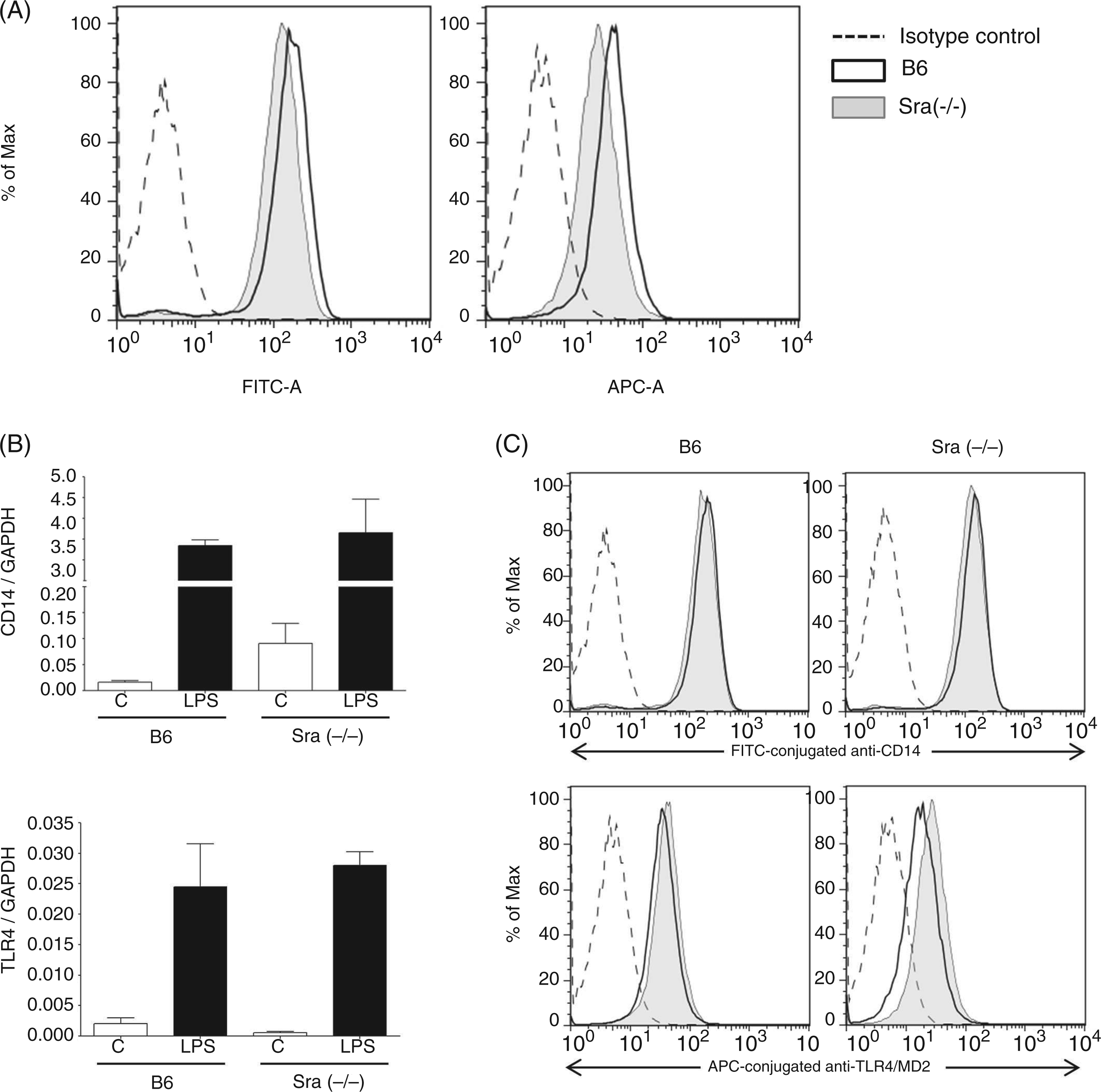
Sra (Msr1) does not contribute to LPS binding
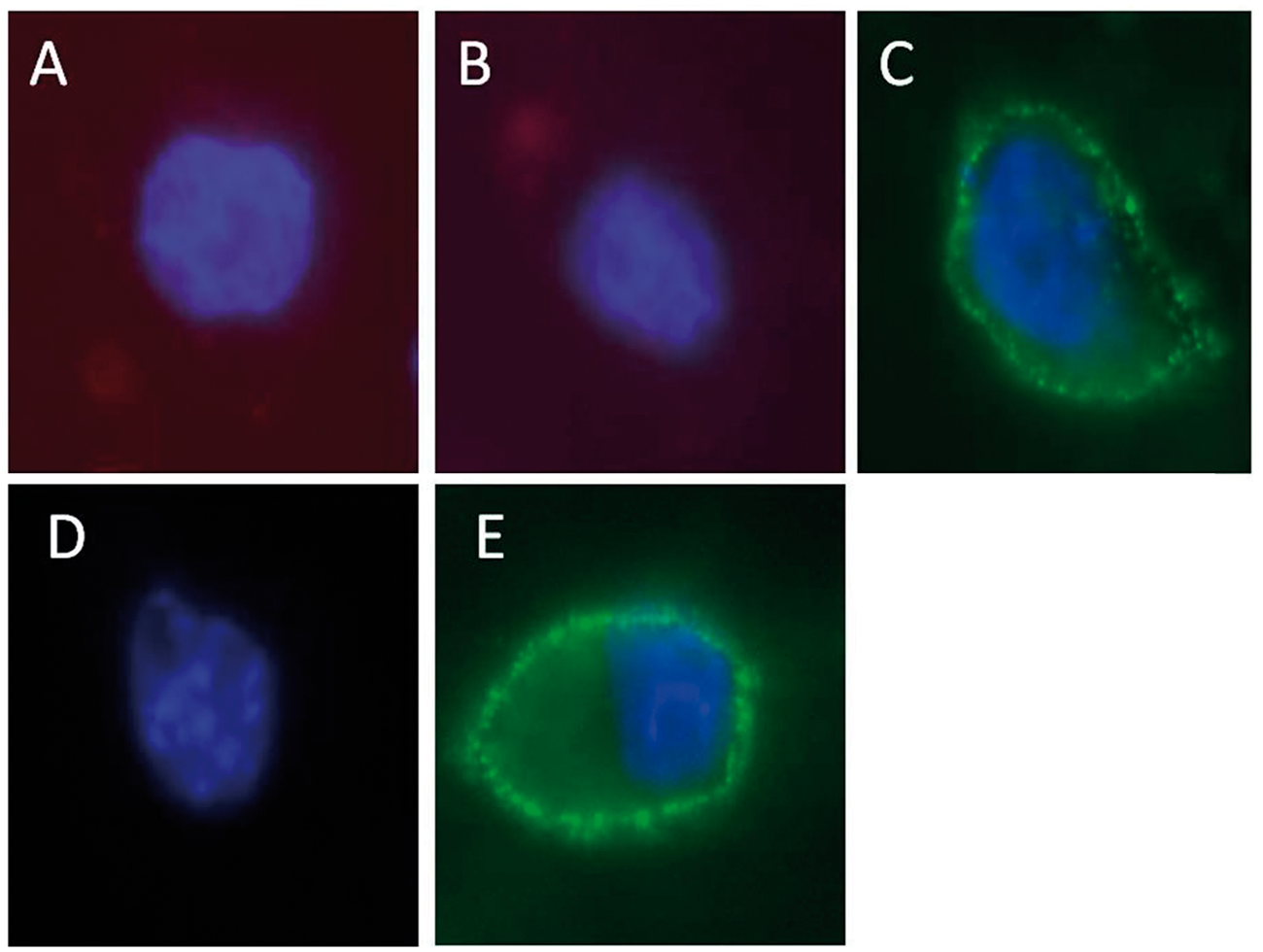
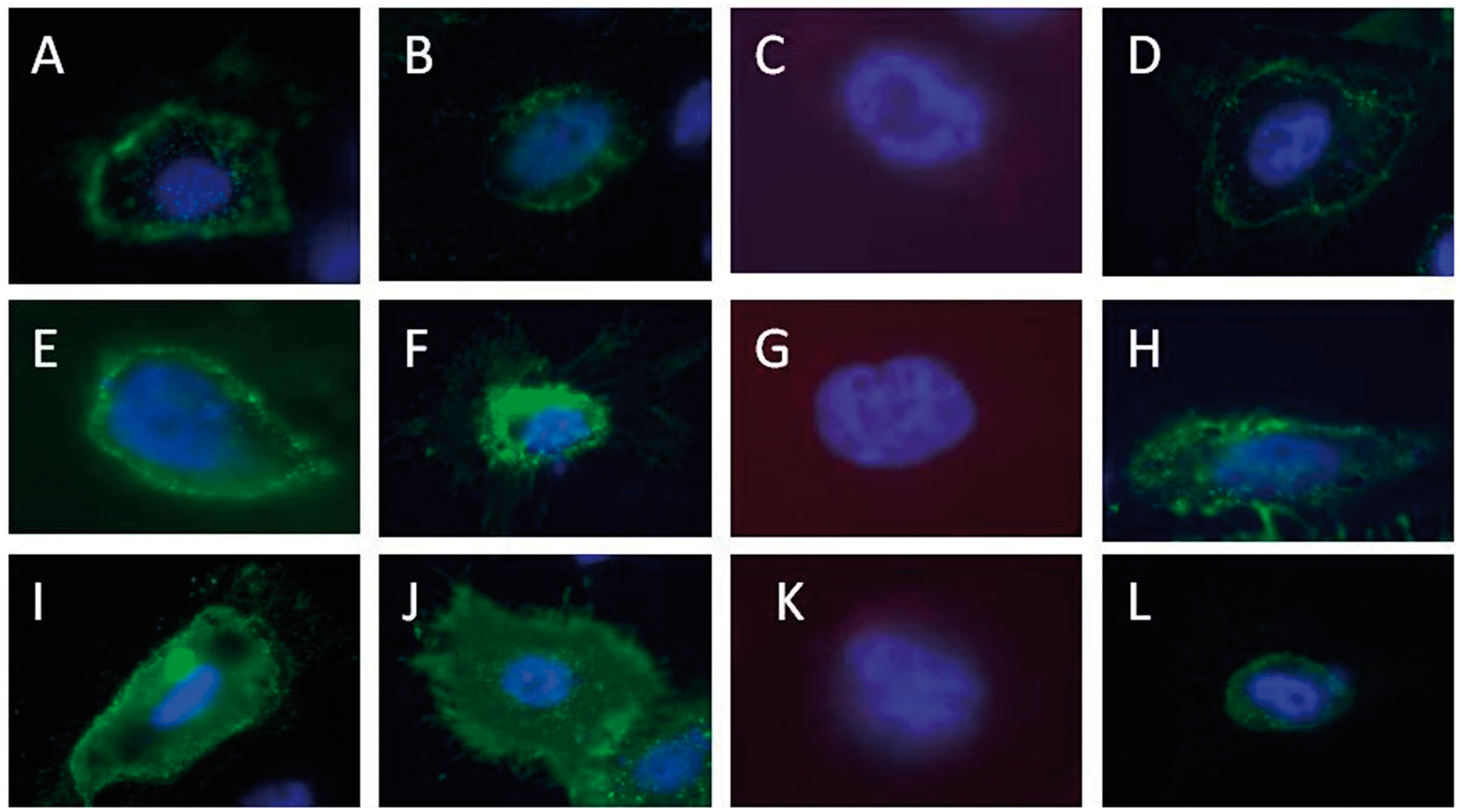
The above observations suggest that the TLR4 pathway, which is the major signal transducer for LPS, is altered in absence of the Sra gene at low concentrations of LPS, but not at higher amounts of the endotoxin. As the interaction of LPS with Sra has been proposed previously,2,35 we investigated whether or not LPS binds directly to Sra. Binding of biotin-conjugated LPS to PMϕs isolated from B6 or Sra(−/−) mice was measured by flow cytometry. We did not observe any difference in LPS binding between the knockout- and wild type-derived PMϕs (Figure 4), suggesting that if, indeed, LPS binds to Sra, it may be just a small contribution to the total binding of endotoxin. To further corroborate the preceding observations, CHO cells that did not present any LPS binding proteins were transfected with mouse Sra gene and binding of biotin-conjugated LPS was visualized by fluorescent microscopy. Mock-transfected CHO cells did not display any detectable LPS binding (Figure 5A). Similarly, CHO cells transfected with mouse Sra did not present any association with LPS (Figure 5B). The presence of Sra on the cell surface of transfected cells was demonstrated by using an Ab specific for mouse Sra (Figure 5C), but not by the secondary Ab alone (Figure 5D). Moreover, we demonstrated by positive binding of Alexafluor 488-labeled AC-LDL that Sra on the surface of transfected cells was functional (Figure 5E). The two isoforms of the Sra gene (Sra-I or Sra-II), which are present on Mϕs, could differ in LPS binding. Thus, CHO cells were transfected with either SraI or SraII, or both, and surface expression of these receptors was detected by various reagents. Indeed, both SraI and SraII were recognized by anti-Sra Ab (Figure 6A, E, I), and both isoforms bound acetylated (Ac)-LDL (Figure 6C, G, K). However, no LPS binding was observed in cells transfected with each of them nor a combination of both (Figure 6D, H, L). The preceding observations suggest that Sra (Msr1) does not bind to LPS.
PMϕs isolated from Sra(−/−) or B6 mice showed an identical LPS binding profile. Naïve PMϕs were isolated from Sra(−/−) or B6 mice by peritoneal lavage in the absence of recruiting agents, as described in the ‘Materials and methods’. (A) Isolated PMϕs were characterized by staining with APC-conjugated anti-F4/80 Abs and biotin-conjugated LPS followed by AlexaFluor488-conjugated streptavidin. An isotype control was also included. Notice that there is no difference in LPS binding between PMϕs isolated from Sra(−/−) or B6 mice. Fluorescence was acquired using a BD FACSCanto II flow cytometer and analyzed by FlowJo software. CHO cells transfected with Sra did not display any LPS binding. CHO cells were transfected with murine Sra (plasmid) using Fugene HD. After 24 h of the transfection, cells were analyzed using various reagents. (A) Mock-transfected CHO cell; (B–E) Sra transfected CHO cells. Cells were stained with biotin-conjugated LPS followed by AlexaFluor594-conjugated streptavidin (A, B), anti-Sra Ab and AlexaFluor488-conjugated anti-rabbit secondary Ab (C), AlexaFluor488-conjugated anti-rabbit secondary Ab in absence of primary Ab (D), and AlexaFluor488-conjugated-Ac-LDL (E). Notice the lack of LPS binding to Sra transfected cells. CHO cells transfected with SraI, SraII, or both did not bind LPS. CHO cells were transfected with murine SraI, SraII, or both (plasmid) using Fugene HD. After 24 h of the transfection, cells were analyzed using various reagents. SraI-transfected CHO cells (A–D), SraII (E–H), SraI and SraII (I–L). Cells were stained with anti-Sra Ab and AlexaFluor488-conjugated anti-rabbit secondary Ab (A, C, E, I); AlexaFluor488-conjugated anti-rabbit secondary Ab in absence of primary Ab (B, F, J); AlexaFluor488-conjugated-Ac-LDL (C, G, K); biotin-conjugated LPS followed by AlexaFluor594-conjugated streptavidin (D, H, L).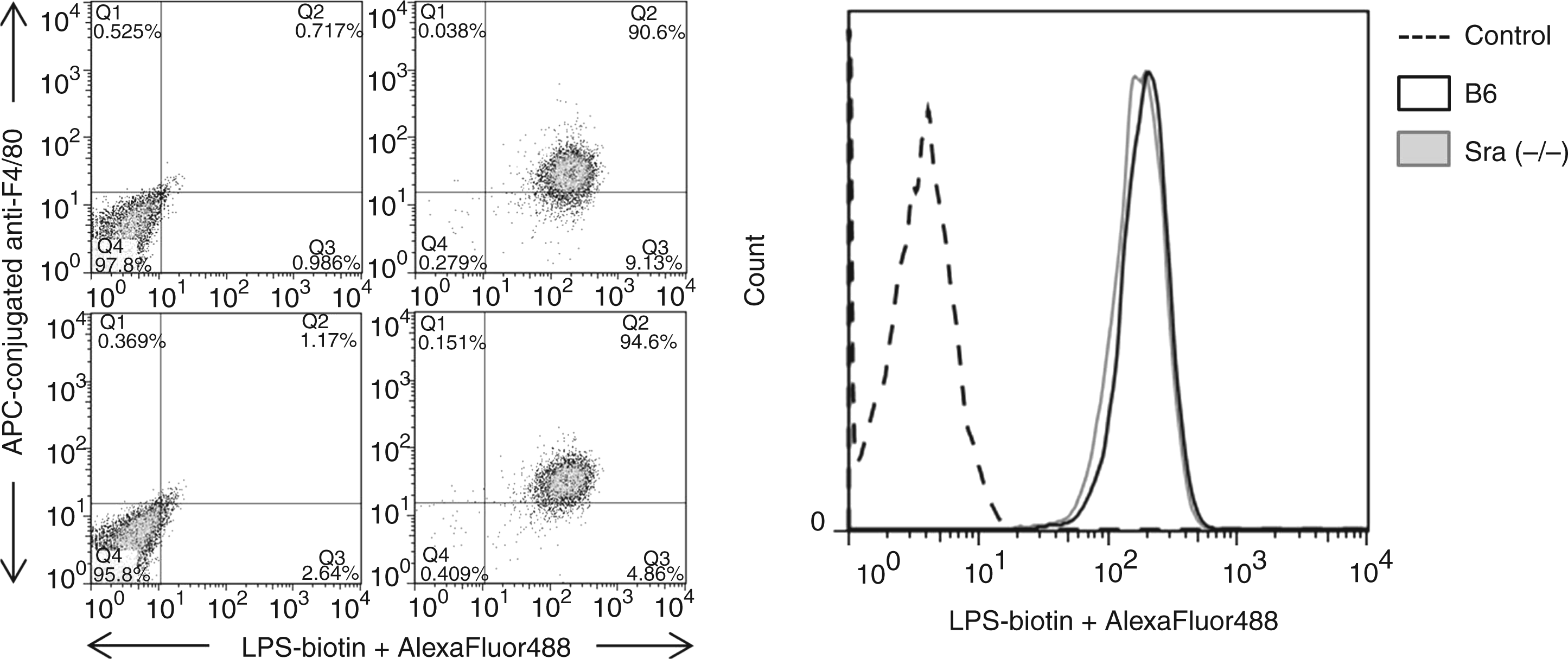
Sra(−/−) mice displayed an alteration of lipid profile in comparison with B6 mice
As protection in Sra(−/−) mice from endotoxic and septic shock is unlikely to be related to the presence of LPS, we searched for other factors that could explain the improved survival of Sra(−/−) mice from LPS and CLP. We measured the lipid profile of naïve Sra(−/−) or B6 mice in non-operated conditions or after CLP. Total cholesterol levels were elevated in Sra(−/−) mice as compared with B6 mice, and no change in these levels were observed after CLP in both strains (Figure 7A), as reported previously.
36
The elevated total cholesterol levels in Sra(−/−) mice were mainly due to high HDL levels in comparison with B6 mice (Figure 7B). No alterations in HDL levels were observed after CLP (Figure 7B). Similarly, LDL levels (calculated) were higher in Sra(−/−) than in B6 mice (Figure 7C). An increase in LDL levels was observed in B6 mice after CLP, but not in Sra(−/−) mice (Figure 7C). There was no difference in triglycerides between Sra(−/−) and B6 mice, and a decrease after CLP was detected in both cases (Figure 7D). Very low density lipoprotein (VLDL) and lipase levels were also not different between Sra(−/−) and B6 mice (Figure 7E, D). Based on these observations, it could be proposed that high levels of HDL may be responsible for the improved survival of Sra(−/−) mice during sepsis. Indeed, several reports have shown that HDL is capable of reducing the inflammatory response induced by LPS.37,38
Lipid profile of Sra(−/−) and B6 mice. Male Sra(−/−) (n = 10) or B6 (n = 12) mice were NPO for 16 h before the procedure. Anesthetized mice were subjected to CLP (1.5-cm ligation) and puncture with a 16-guage needle or were non-operated [NO; B6 n = 5; Sra(−/−) n = 3]. The lipid profile was determined in plasma samples from NO mice, 6 or 20 h after CLP. Total cholesterol (A), HDL (B) calculated LDL (C), triglyceride levels (D), calculated VLDL (E), and lipase (F). Significance was analyzed using a one-way ANOVA followed by Newman-Keuls multiple comparison test for each time point (P < 0.0001).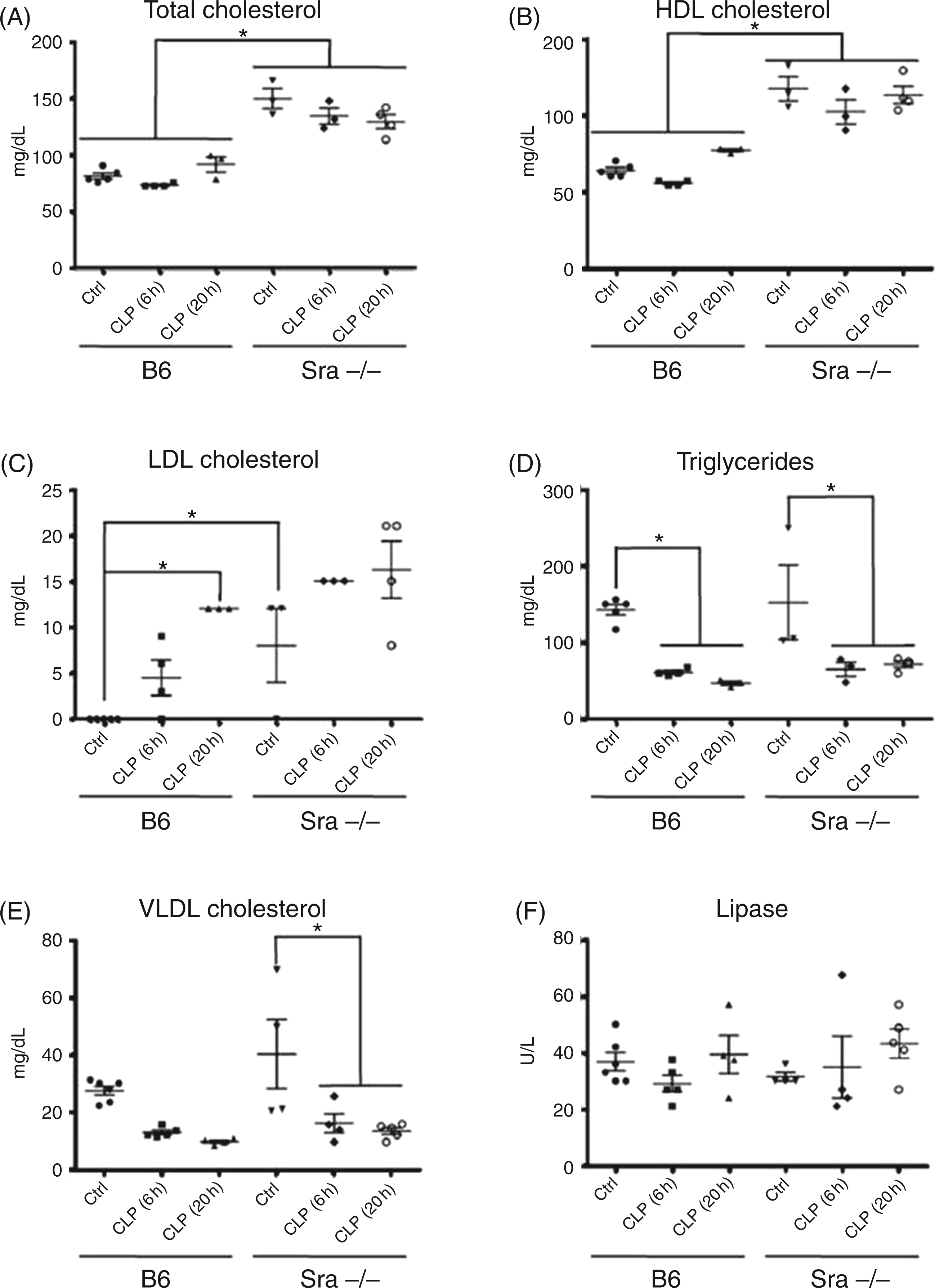
Discussion
The innate immune system is the initial and more rapid response to infections and injury. Pathogens, dead cells, and cellular debris are recognized by a group of cell surface receptors named PRRs, which direct clearance and activation of necessary pathways to resolve the insult. Scavenger receptors, which are part of PRRs, have gained increasing attention because of the versatility of the ligands that they recognize and the cellular processes in which they are involved.2,4 We have previously identified a member of the scavenger receptor family, Sra1 or Msr1, as a modifier gene of the inflammatory response induced by LPS during a genetic screening. 28 Moreover, we observed that Sra(−/−) mice presented reduced LPS-induced IL-10 plasma levels in comparison with wild type B6 mice. 28 In the present study, we evaluated whether or not the presence of Sra altered the response to a clinically relevant septic insult induced by CLP, which is the gold standard animal model for the study of sepsis. We observed that Sra1(−/−) mice displayed a better survival rate than wild-type mice during sepsis induced by two variants of the CLP model that differ in the severity of the insult. Therefore, these observations clearly displayed an advantage in the outcome by the absence of Sra. Prior studies have shown increased mortality of Sra1(−/−) mice after injection with LPS,22–24 or after a combination of LPS and priming with bacillus Calmette-Guerin. 21 In contrast, other studies have reported increased survival of Sra(−/−) mice after injection of LPS. 25 As the CLP model is composed of an area of necrosis and leakage of the gut polymicrobial flora into the peritoneum, it is likely that it does not depend on LPS toxicity alone. Thus, other factors in this insult are likely to play a role in the outcome, in which Sra may play a critical part. Indeed, we did not find any direct binding of biotin-conjugated LPS to naïve PMϕs isolated from Sra(−/−) in contrast to B6 (wild-type)-derived cells or after transfection of CHO cells with SraI or SraII genes. Previous studies have proposed that Sra1 interacts with LPS, as assessed by competition binding of lipid A, which is a component of LPS, and typical Sra ligands. 35 However, no direct LPS binding to Sra was demonstrated. Peiser et al. 13 showed that Sra recognized N. meningitidis independently of the presence of LPS.
In spite of the lack of direct binding of LPS to Sra, we observed that the response to LPS via the TLR4 signal transduction pathway was altered in PMϕs isolated from Sra(−/−) mice. The absence of Sra did not result in altered expression of TLR4 or CD14, which could have been a compensatory mechanism to increase LPS binding. We observed that deletion of the Sra gene resulted in reduced IL-10 and IL-6 mRNA levels after challenging naïve PMϕs with low concentrations of LPS (1 ng/ml). In contrast, expression of TNF-α was not affected by the presence (or not) of Sra, suggesting that the TLR4-NF-κB pathway is independent of the presence of the scavenger receptor. The reduced expression of IL-10 and IL-6 in PMϕs derived from Sra(−/−) mice was not observed at high concentrations of LPS (100 ng/ml). There is an important distinction between these two doses of LPS. The activation of Mϕs at low concentrations (1 ng/ml) is exclusively dependent on the presence of CD14, as demonstrated in CD14(−/−) mice. 34 At higher concentrations of LPS (e.g. 100 ng/ml), LPS is recognized by reduced affinity receptors, such as TLR4. 39 Based on this assumption, we propose that Sra forms a complex with CD14 and TLR4 that is involved in the expression of IL-6 and IL-10 at low LPS concentrations, which are likely similar to the concentrations of LPS in circulation during infection. Previous studies have also proposed an interaction of Sra with TLR49,39,40 and a modulation of the TLR4 signal transduction pathway.23,24,41,42 Recently, it has been reported that TLR4 can differentially activate the TIRAP-MyD88 and the TRAM-TRIF pathways depending on the subcellular localization of this receptor. Thus, the TIRAP-MyD88 pathway is activated when TLR4 is on the plasma membrane, whereas the TRAM-TRIF pathway is triggered by the internalization of TLR4, which is localized within early endosomes. 43 As the TRAM-TRIF pathway is directly involved in IL-10 expression and indirectly in IL-6 production, Sra may modulate the translocation of TLR4 from the plasma membrane to early endosomes, resulting in alteration of cytokine expression. In this regard, we observed small differences in the down-regulation of TLR4 from the cell surface in presence or absence of Sra after stimulation with LPS. Other studies have found that internalization of Sra is required for Mϕ activation via various ligands for this receptor. 44 Elegant studies by Seimon et al. 9 have shown that ligands for Sra, such as fucoidan, triggered apoptosis in ER-stressed Mϕs via association with TLR4 and blocked the pro-survival IRF-3-IFN-β pathway. In contrast, LPS triggered an alternative survival pathway via the activation of TRIF-TRAM and, subsequently, IRF3 and the production of IFN-β. These observations suggest the activation of two alternative pathways in Mϕs by typical ligands of Sra (fucoidan) or TLR4 (LPS). The independence between these two pathways may be explained by the lack of LPS binding to Sra. Similarly, Sra has been shown to affect dimerization and ubiquitination of TNF receptor-associated factor 6 (TRAF6), resulting in an alteration of the inflammatory response. 23 Other studies have also suggested that Sra attenuated the response of TLR4 and enhanced the activation by intracellular receptors, such as TLR3, NOD2, and NALP3. 40
Although the role of Sra in the septic response remains to be established, it may be related to the modulation of TLR4 signal transduction pathways. Another alternative to explain the survival of Sra(−/−) mice during sepsis may be associated with the initial elevated levels of HDL in circulation that we observed in these animals (Figure 7). HDL has been shown to neutralize LPS toxicity 45 and it has been reported to bind free LPS, as well as to remove LPS from the cell surface. 46 Elevated HDL has also been correlated with protection from endotoxic shock in humans 47 and in animal models.48–50 In addition, high levels of HDL have been described to reduce organ injury after hemorrhagic shock 51 and ischemia/reperfusion, 52 suggesting that the effect of this lipoprotein may be beyond LPS neutralization. Several reports have certainly indicated anti-inflammatory and antioxidant effects of HDL.37,38 A HDL-mediated reduction in the inflammatory response has been shown during endotoxic shock.37,48,53 Reduced total cholesterol, LDL and HDL levels were observed in circulation in patients after injury 54 and sepsis. 55 This reduction in HDL has been correlated with a negative outcome.55–57 In fact, an increase in HDL levels over time that was not detected in non-survival septic patients was observed in septic shock survival patients in an intensive care unit. 58 Therefore, it is possible that the improved survival rate endotoxic and septic shock observed in Sra(−/−) mice as opposed to wild-type may be linked to the initial elevated HDL levels observed in the knockout animals. The higher HDL levels in Sra(−/−) versus B6 mice is puzzling. High levels of HDL have been associated with reduced expression of scavenger receptor B or CD36 in human studies. 59 This observation was clearly confirmed in investigations using CD36 knockout mice. 60 However, a possible role of Sra in the clearance of HDL has not been established. Plasma HDL levels have been correlated with genetic factors. 61 Mouse genetic studies have reported a quantitative trait locus (QTL) associated with HDL within chromosome 8.62,63 We have also identified a QTL for the inflammatory response to LPS located within mouse chromosome 8, which contains Sra (Mrs1) as a candidate gene. 28 However, there was no overlap between these two regions. In contrast, other genetic studies analyzing plasma fatty acid levels in Alaskan Eskimos resulted in a QTL on chromosome 8 containing Sra (Msr1), which was also considered as a candidate gene. 64
In summary, our studies suggest that Sra is a modulator of the signal transduction pathway mediated by TLR4, altering the inflammatory process. In addition, our observations demonstrated that the absence of Sra in mice resulted in a better survival rate from septic shock. This increase in survival is not a result of the contribution of Sra to sequester LPS, but is rather associated with an increase in basal levels of HDL. The possibility that elevated HDL levels could improve survival from sepsis may open new and exciting approaches to ameliorate the detrimental consequences of sepsis.
Footnotes
Funding
These studies were supported by the National Institute of General Medical Sciences (grant no. GM073825).
Acknowledgements
We thank Professor Motohiro Takeya (Department of Cell Pathology, Kumamoto University, Kumamoto, Japan) for the generous gift of anti-Sra Ab.