
Abstract
The effect of 10-hydroxy-trans-2-decenoic acid (10H2DA), a major fatty acid component of royal jelly, was investigated on LPS-induced cytokine production in murine macrophage cell line, RAW264 cells. 10H2DA inhibited LPS-induced IL-6 production dose-dependently, but did not inhibit TNF-α production. 10H2DA inhibited LPS-induced NF-κB activation in a dose-dependent fashion. In addition, NF-κB activation induced by over-expression of either MyD88 or Toll/IL-1 receptor domain-containing adaptor inducing IFN-β (TRIF) was also inhibited by 10H2DA. Degradation of IκB-α and phosphorylation of IκB kinase-α were not inhibited by 10H2DA. On the other hand, reduction of LPS-induced IκB-ζ expression was discovered. Production of lipocalin-2 and granulocyte colony-stimulating factor (G-CSF), which is dependent on IκB-ζ, was also inhibited by 10H2DA, whereas that of IκB-ζ–independent cytokines/chemokines, such as IFN-β, murine monocyte chemotactic protein-1 (JE), macrophage inflammatory protein (MIP)-1α and MIP-2, was not. Together, 10H2DA specifically inhibited LPS-induced IκB-ζ expression, followed by inhibition of IκB-ζ-dependent gene production. These results suggest that 10H2DA is one of the components of royal jelly to show anti-inflammatory effects and could be a therapeutic drug candidate for inflammatory and autoimmune diseases associated with IκB-ζ and IL-6 production.
Introduction
TLRs are innate immune receptors recognizing microbial components such as bacterial LPS. 1 The TLR4/MD-2 complex recognizes LPS and its signaling activates NF-κB in two independent pathways, such as MyD88-dependent and Toll/IL-1 receptor domain-containing adaptor inducing IFN-β (TRIF)-dependent pathways. 2
As many inflammatory cytokines and chemokines are produced under the regulation of NF-κB, activation of NF-κB is strictly regulated in several ways.3,4 Nuclear factor-κB proteins are homodimers or heterodimers of the NF-κB family including p65 and p50. IκB-ζ, a member of IκB family, can be a component of an active NF-κB complex, especially in activating IL-6 promoter induced by LPS. 5 NF-κB p65 is sequestered in the cytoplasm through association with IκB in unstimulated cells. During LPS stimulation, p65 is released from IκB-α, a typical member of IκB, followed by nuclear translocation of the p50/p65 heterodimer.
Royal jelly is produced by the hypopharyngeal and mandibular glands of worker honeybees as a food essential for the growth of the queen honeybee. It has various pharmacological activities, such as vasodilative hypotensive activities, decreasing activity of serum cholesterol levels, anti-tumor activities, protective activity against hemopoietic dysfunction in X-irradiated mice, anti-allergic activities of reducing IgE production and anti-inflammatory activities in inhibiting pro-inflammatory cytokine production. 6 – 10 Royal jelly contains 10-hydroxy-trans-2-decenoic acid (10H2DA) as major lipid component, 11 which is a natural ingredient almost uniquely found in royal jelly. Therefore, it is possible that 10H2DA is an active component of royal jelly’s pharmacological activities. Indeed, anti-tumor and antibiotic activities have been reported for 10H2DA.8,12 However, the molecular mechanisms for the pharmacological activities of 10H2DA are not well known.
In this study, we investigated the effect of 10H2DA on LPS-induced cytokine production.
Materials and methods
Reagents
10H2DA with a purity of above 98% (analyzed by high performance liquid chromatography), was kindly gifted by Api Co. (Gifu, Japan). It was dissolved in ethanol at 500 m
Plasmids
Murine Toll/IL-1 receptor domain-containing adaptor protein (TIRAP) gene was amplified from mRNA of RAW264 by RT-PCR and cloned into pENTR/TEV/D-TOPO (Invitrogen, Carlsbad, CA, USA). It was then transferred into pDEST26 (Invitrogen), as recommended by the manufacturer. The MyD88 expression vector 13 was kindly gifted by Dr Takaoka (University of Tokyo), whereas TRIF 14 was from Dr Komatsu (Aichi Medical University). Promoter regions of TNF-α, IL-6, and IκB-ζ gene were amplified from the genomic DNA of RAW264 by PCR and cloned into a luciferase reporter plasmid, pGL4.26 (Promega, Madison, WI, USA). An NF-κB reporter gene plasmid, pNF-κB-TA-luc, was purchased from Clontech (Palo Alto, CA, USA). A transfection control reporter plasmid, pRL-TK, was obtained from Promega. Plasmids were extracted with GenElute HP endotoxin-free plasmid Midiprep kit (Sigma-Aldrich) for transfection into cultured cell lines.
Cell culture, 10H2DA treatment and stimulation
The RAW264 murine macrophage cell line was obtained from RIKEN Bioresource Center (Tokyo, Japan) and maintained in Roswell Park Memorial Institute (RPMI) 1640 medium containing 5% heat-inactivated fetal bovine serum (FBS; Invitrogen). The cells were seeded in a 96-well plate at 2 × 105 cells/well for cytokine assays or 2.5 × 104 cells/well for reporter gene assays. The cellswere pre-incubated with or without the indicatedconcentration of 10H2DA for 30 min then stimulated by adding LPS at the final concentration of 100 ng/ml. After 24 h incubation, the culture supernatants wereused for cytokine assays. The human embryonic kidney (HEK) cell line HEK293 was maintained in minimal essential medium containing 5% FBS and 1% nonessential amino acid (Sigma-Aldrich). The HEK293 cells were seeded in a 96-well plate at a cell density of2.5 × 104 cells/well for reporter gene assays. At theend of each assay, cell viability was assayed by AlamarBlue (Trek Diagnostic Systems, West Sussex, UK), as recommended by the manufacturer. In all experimental conditions performed in this study, cell viability was >95% compared with untreated cells.
Analysis for cytokines/chemokines expression by RT-PCR
Primers used for RT-PCR analysis
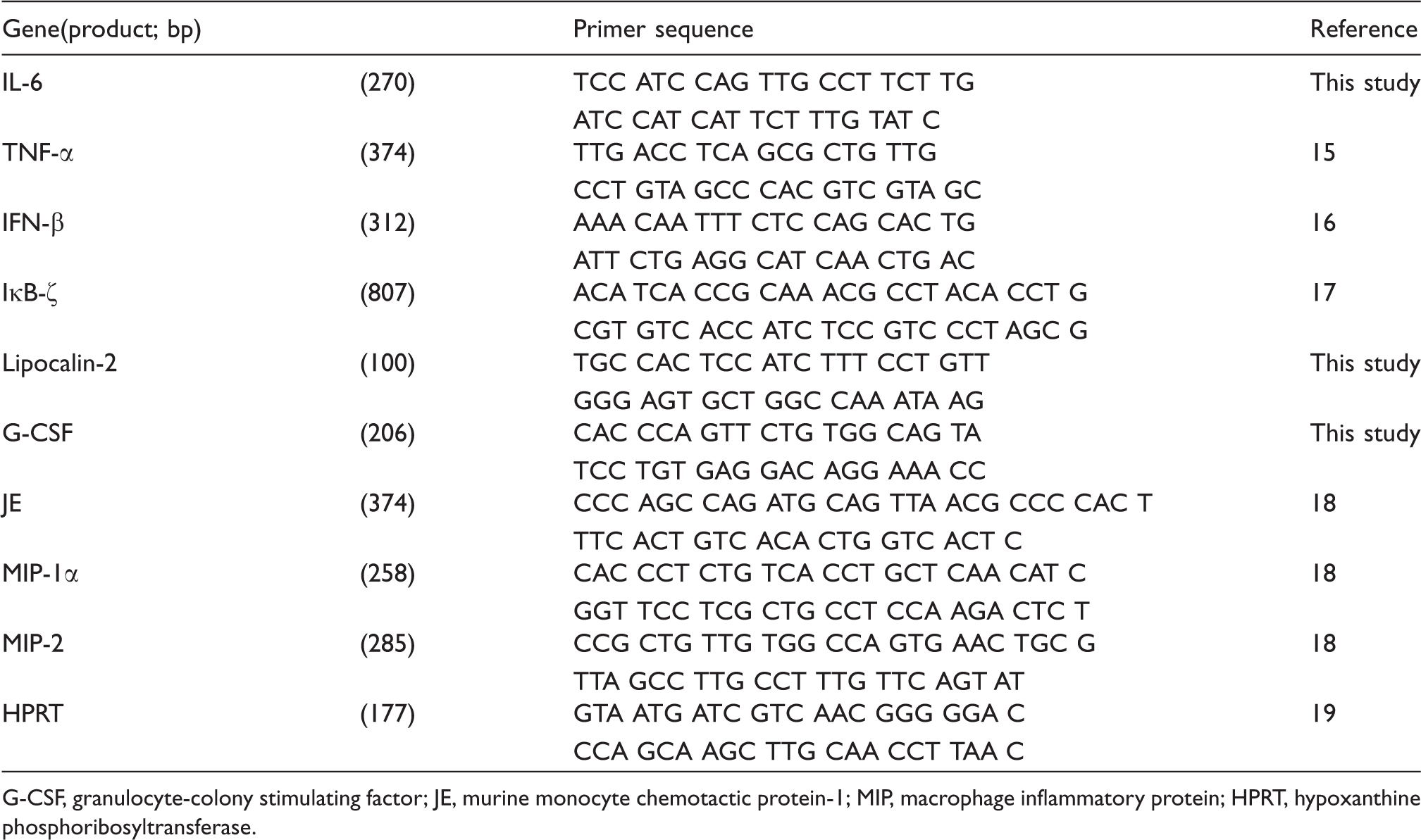
G-CSF, granulocyte-colony stimulating factor; JE, murine monocyte chemotactic protein-1; MIP, macrophage inflammatory protein; HPRT, hypoxanthine phosphoribosyltransferase.
Cytokine analysis
The TNF-α and IL-6 concentrations in the culture supernatant were analyzed by conventional sandwich ELISA. Antibodies and recombinant cytokines for ELISA were purchased from e-Bioscience (San Diego, CA, USA).
Reporter gene assay
Cells seeded in wells of a 96-well plate were transfected with a luciferase reporter construct and pRL-TK plasmid using FuGENE-HD (Roche, Indianapolis, IN, USA) for RAW264 or Lipofectamine 2000 (Invitrogen) for HEK293, as recommended by the manufacturer. After indicated treatment, the cells were lysed with Passive Lysis Buffer (Promega). Luciferase activity in the cell lysate was determined by Dual-Luciferase Reporter assay system (Promega).
Immunoblotting
The RAW264 cells were pretreated with 10H2DA (4 m
Statistical analysis
Results are expressed as means ± SD. Statistical analyses were performed using GraphPad PRISM software (GraphPad, San Diego, CA, USA). Significant differences between control and 10H2DA treatment were evaluated by Dunnett’s multiple comparison test after one-way ANOVA and by Student’s t-test. A P-value of <0.05 was considered statistically significant.
Results
10H2DA inhibited LPS-induced IL-6 but not TNF-α production
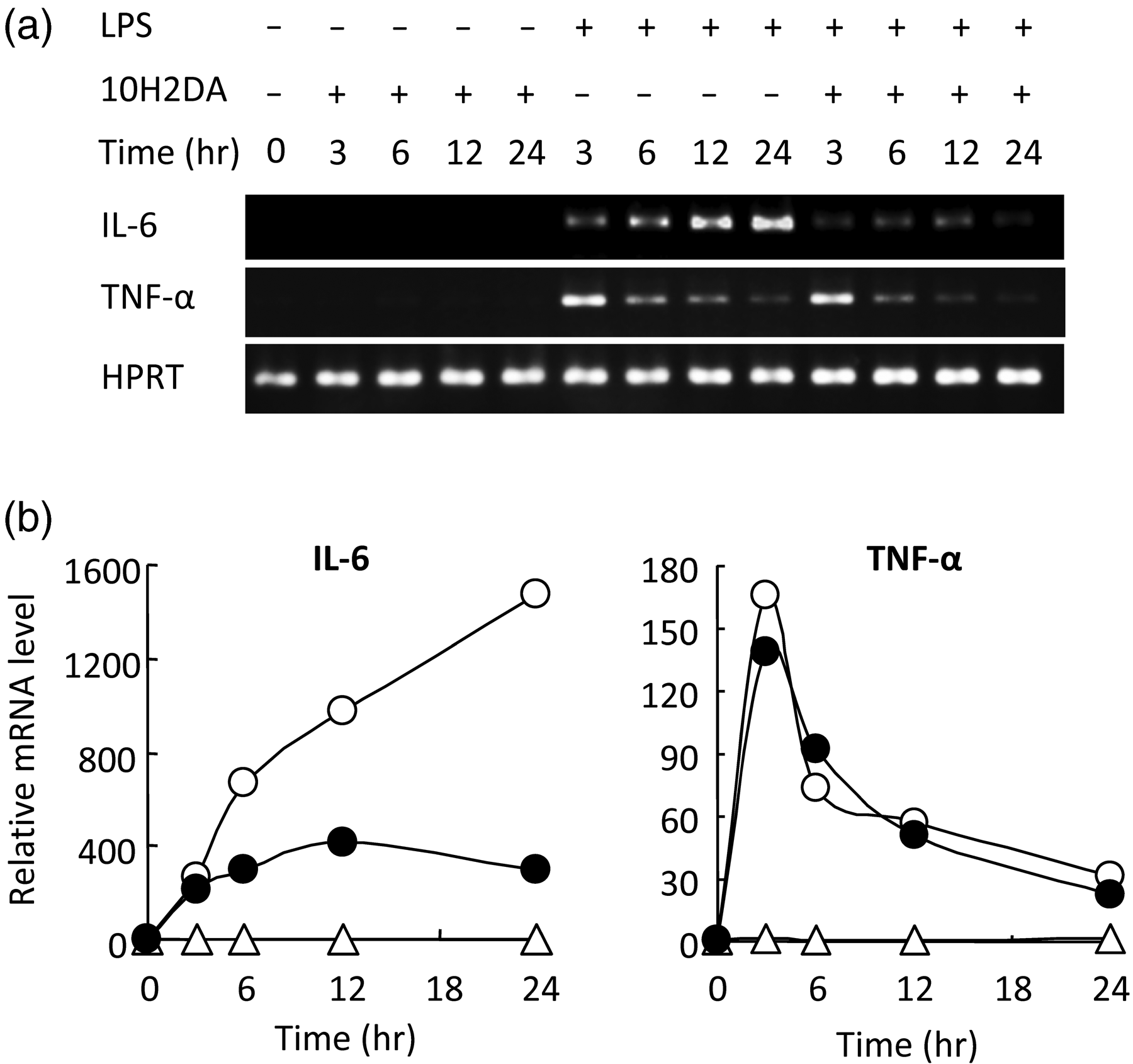
10H2DA significantly inhibited IL-6 but not TNF-α production at concentrations 2–4 m Effect of 10H2DA on LPS-induced IL-6 and TNF-α production. RAW264 cells were incubated with 0, 1, 2, or 4 m Effect of 10H2DA on LPS-induced IL-6 and TNF-α mRNA expression. RAW264 cells were incubated with or without 4 m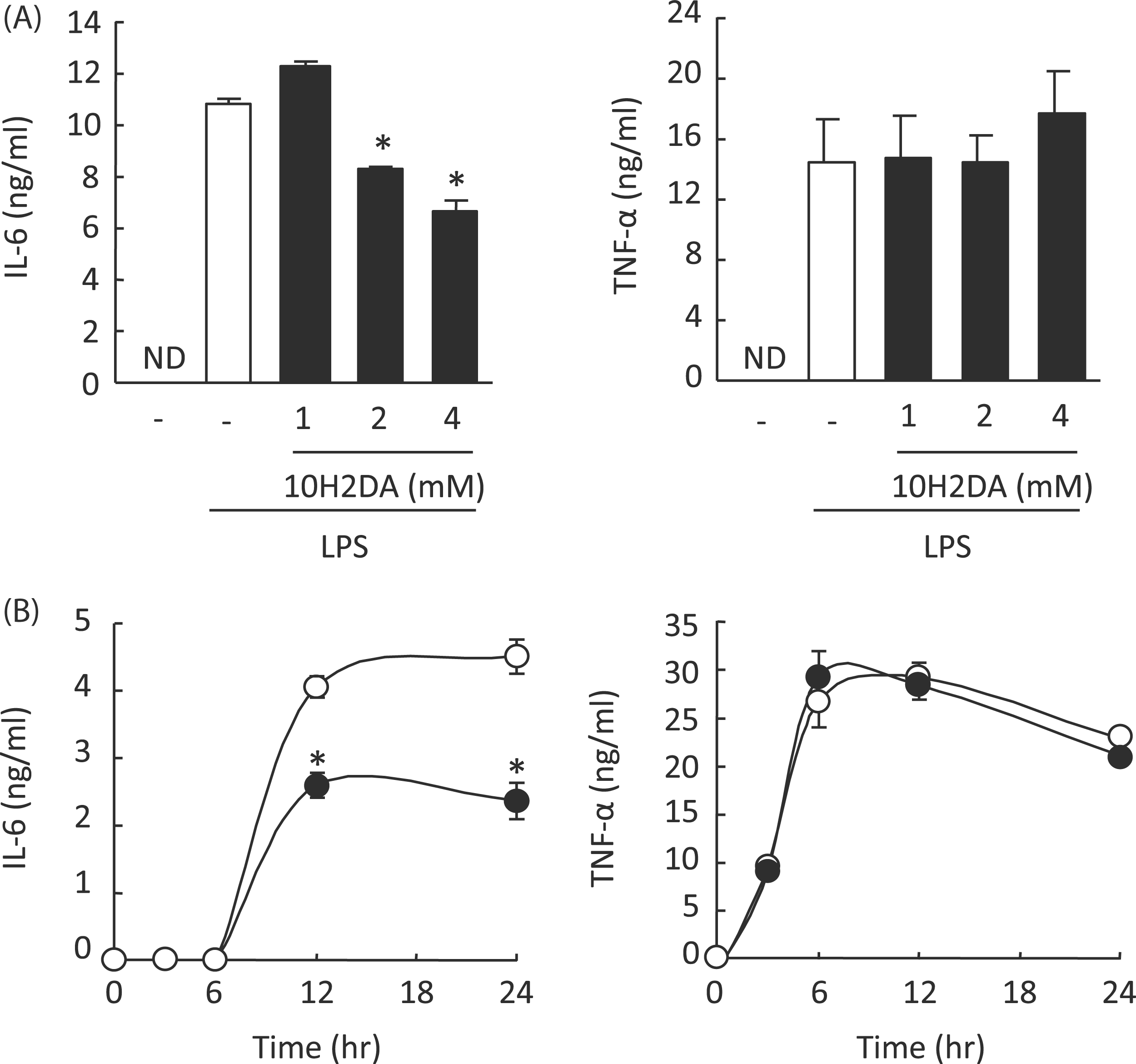
10H2DA inhibited NF-κB activation
In RAW264 cells NF-κB activation with LPS was dose-dependently inhibited by 1–4 m Effect of 10H2DA on NF-κB activation in LPS signaling. (A) RAW264 cells transfected with pNF-κB-TA-luc (100 ng/well) and pRL-TK (10 ng/well) were incubated with (filled column) or without (open column) the indicated concentration of 10H2DA for 30 min followed by stimulation with 100 ng/ml LPS for 6 h. Luciferase activities in cell lysates were measured, normalized and expressed as fold induction compared with unstimulated control. Data are expressed as mean ± SD (N = 3). *P < 0.05 compared with no 10H2DA. (B) HEK293 cells were transfected with 10 ng/well of control vector pEF (mock) or expression plasmid for MyD88, murine TIRAP, or TRIF. At the same time, pNF-κB-TA-luc (10 ng/well) and pRL-TK (1 ng/well) were co-transfected. Five h after transfection, culture media were exchanged for media containing the indicated concentration of 10H2DA (filled column) or PBS (open column) and the cells were further incubated for 24 h. Luciferase activities in cell lysates were determined. NF-κB activation was expressed as fold induction of luciferase activity compared with that of mock transfectant. Data are expressed as mean ± SD (N = 3). **P < 0.01; ***P < 0.001 compared with no 10H2DA.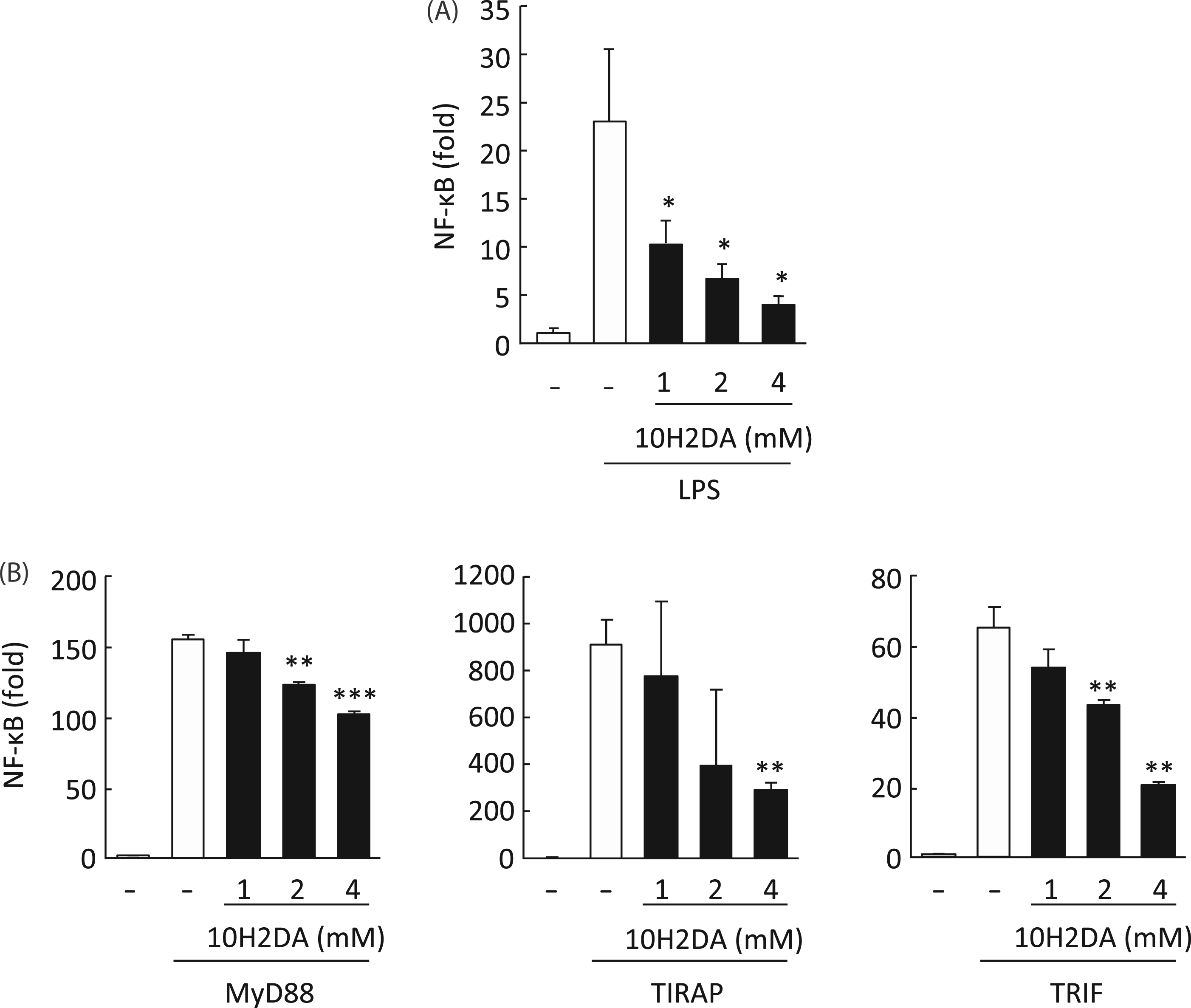
Immunoblotting revealed that 10H2DA did not inhibit LPS-induced IKK-α phosphorylation and IκB-α degradation (Figure 4).
Effect of 10H2DA on LPS-induced IKK-α phosphorylation and IκB-α degradation. RAW264 cells were incubated with 4 m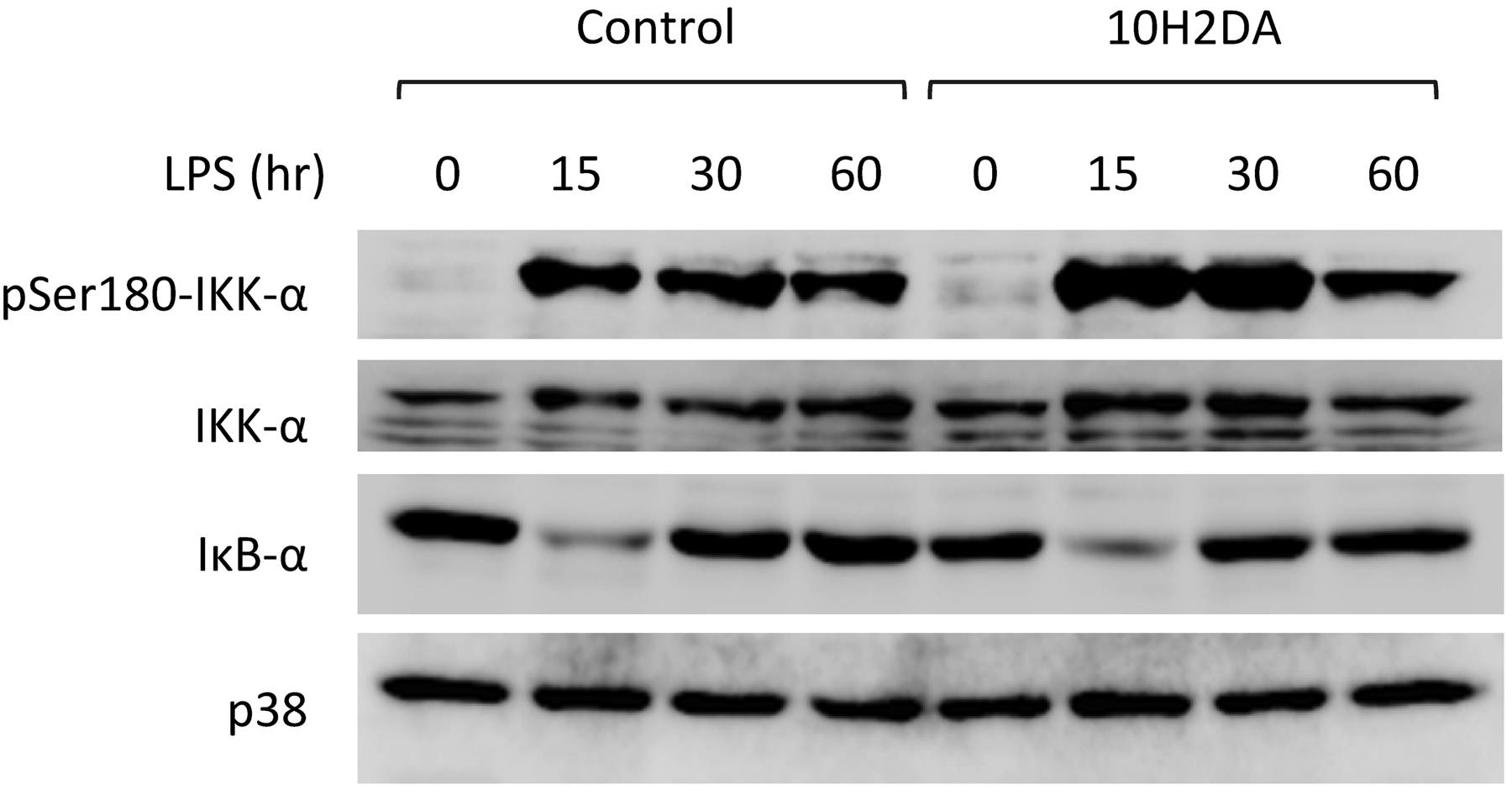
10H2DA inhibited (IκB)-ζ mRNA expression
Analyses by RT-PCR revealed that, under LPS stimulation, expression of IκB-ζ mRNA was increased at 3 h (Figure 5). 10H2DA, on the other hand, inhibited expression of IκB-ζ mRNA.
Effect of 10H2DA on LPS-induced IκB-ζ mRNA transcription. RAW264 cells were incubated with or without 4 m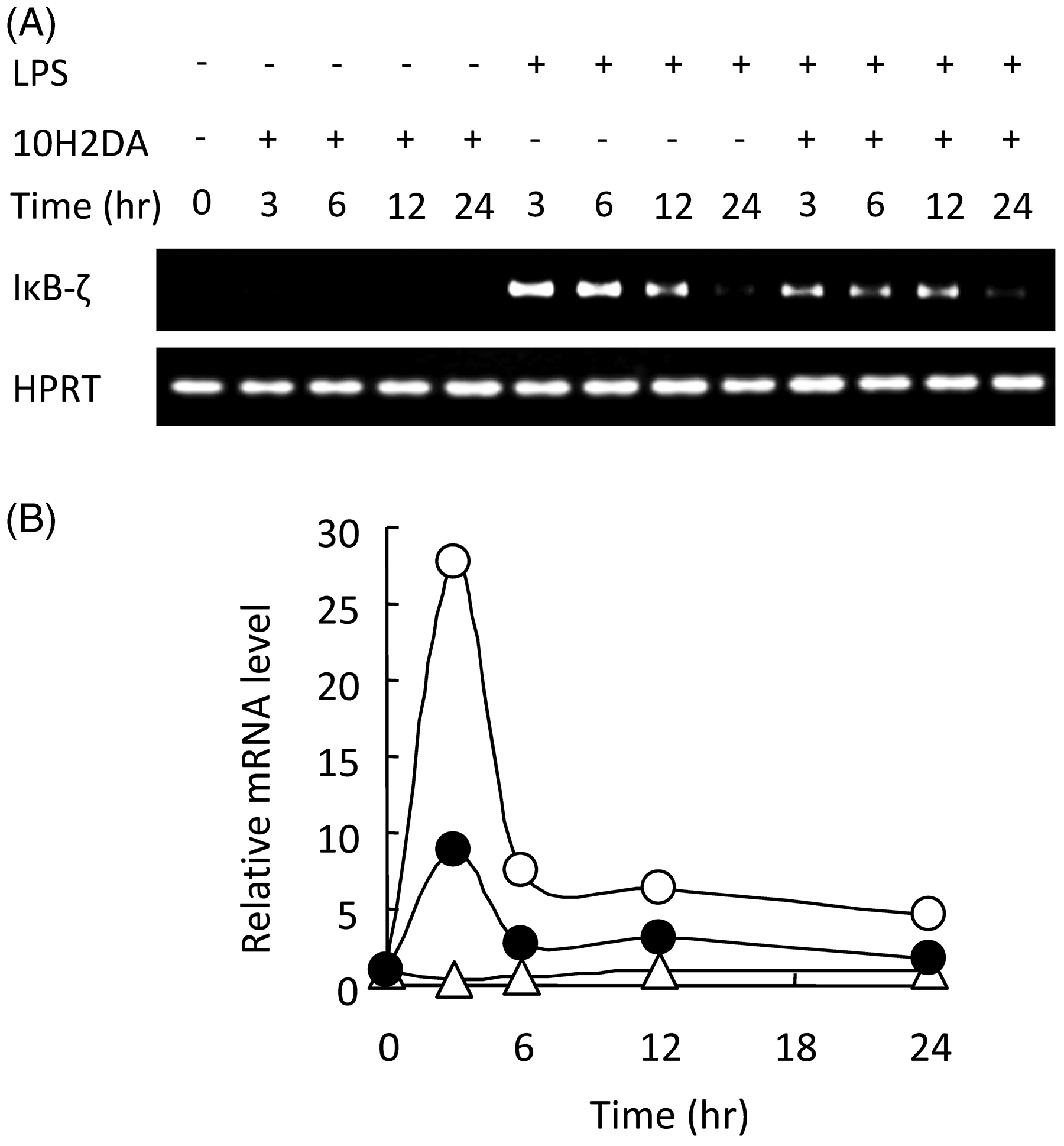
10H2DA inhibited IL-6 and IκB-ζ but not TNF-α transcription induced by LPS and TLR signaling molecules
According to luciferase reporter gene assay, in RAW264 cells LPS induced activation of TNF-α, IL-6 and IκB-ζ promoters (Figure 6). As expected, LPS-induced promoter activity of IL-6 and IκB-ζ, but not TNF-α, was dose-dependently inhibited by 10H2DA (Figure 6A). Furthermore, IκB-ζ promoter activation induced by over-expression of MyD88 was also significantly inhibited by 10H2DA (Figure 6B).
Effect of 10H2DA on LPS-induced TNF-α, IL-6, and IκB-ζ promoter activation. (A) Luciferase reporter construct of TNF-α, IL-6, or IκB-ζ promoter and pRL-TK was transfected into RAW264 cells. The cells were incubated with (filled column) or without (open column) the indicated concentration of 10H2DA for 30 min followed by stimulation with 100 ng/ml of LPS for 6 h (TNF-α andIκB-ζ) or 12 h (IL-6). Luciferase activity in cell lysates was measured, normalized and expressed as fold induction, compared withunstimulated control. Data are expressed as mean ± SD (N = 3). *P < 0.05; **P < 0.01 compared with no 10H2DA. (B) Luciferase reporter construct of IκB-ζ promoter was co-transfected with MyD88-expressing plasmid into HEK293 cells. Five h after transfection,10H2DA was added at the indicated concentration. After 6 h incubation, luciferase activity in cell lysates were measured,normalized and expressed as fold induction, compared with mock-transfectant without 10H2DA. Data are expressed as mean ± SD (N = 3). *P < 0.05; **P < 0.01; ***P < 0.001 compared with no 10H2DA. Open column, without 10H2DA; filled column, with 10H2DA.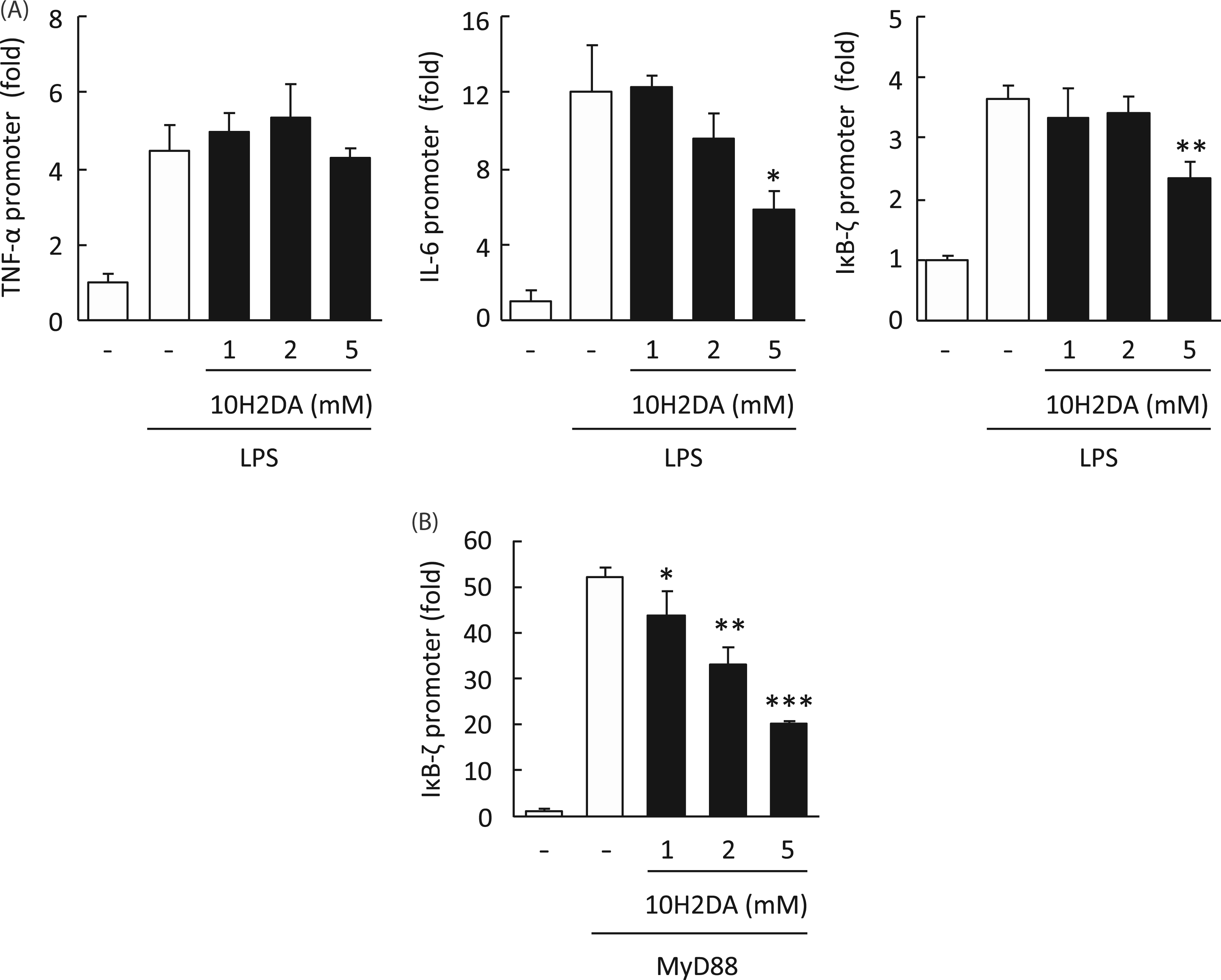
10H2DA inhibited IκB-ζ–dependent gene expression
Expression of lipocalin-2 and G-CSF was dependent on IκB-ζ, in comparison with independent genes IFN-β, murine monocyte chemotactic protein-1 (JE), MIP-1 and MIP-2. The LPS-induced expression of IκB-ζ–dependent genes lipocalin-2 and G-CSF was significantly inhibited by 10H2DA, whereas that of IFN-β, JE, MIP-1 and MIP-2 was not (Figure 7).
Effect of 10H2DA on cytokines/chemokines mRNA expression. The RAW264 cells were incubated with or without 4 m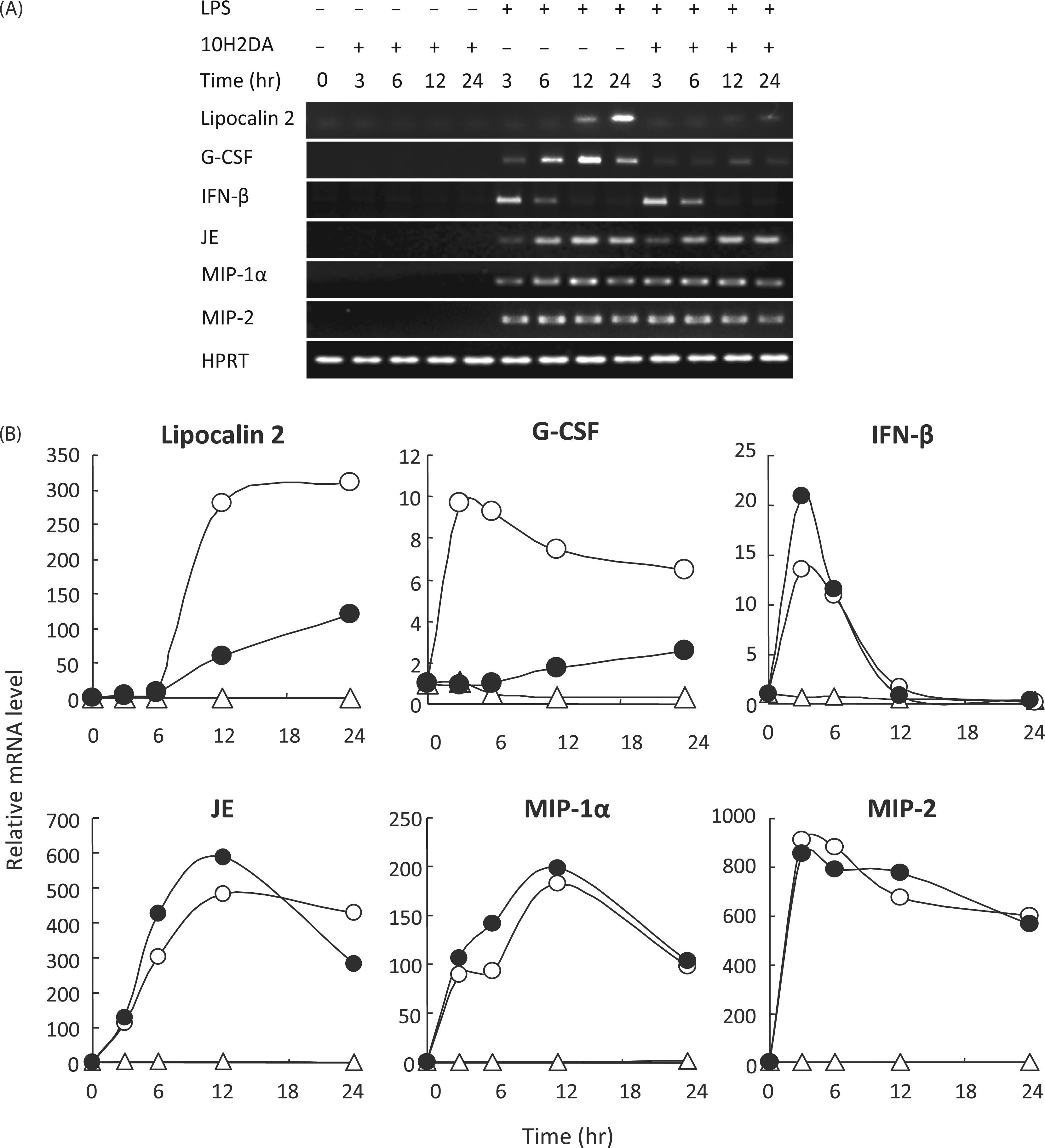
Discussion
De novo synthesis of IκB-ζ has been reported to be critical for IL-6 production in LPS stimulation. 5 After stimulation of IκB-ζ production with LPS through the MyD88-dependent pathway, IκB-ζ interacts with the p50 subunit of NF-κB, following which the active NF-κB complex upregulates IL-6 production.
We revealed specific inhibitory effects of 10H2DA on LPS-induced IκB-ζ production followed by inhibition of IκB-ζ–dependent gene expression, including IL-6 (Figure 8). IL-6 has been reported to play critical roles in colonic inflammation and colon cancer.
20
Recently, IL-6 was found to be a key contributor in the development of Th17 helper T cells, which critically contribute to autoimmune and inflammatory diseases.21,22 Lipocalin-2 and G-CSF genes have also been reported to be IκB-ζ–dependent genes, as well as IL-6.
23
We detected significant inhibition of expression of these genes by 10H2DA, whereas no significant reduction was seen in the expression of IκB-ζ-independent genes, such as TNF-α, IFN-β, JE, MIP-1α and MIP-2, suggesting that a reduction in IκB-ζ expression results in the inhibition of IL-6 production. Lipocalin-2 has been suggested to promote insulin resistance.
24
Furthermore, IκB-ζ is involved in IFN-γ production by TNF-α stimulation.
25
Recently, IκB-ζ has been reported to positively regulate Th17 development in co-operation with ROR nuclear receptors.
26
IκB-ζ-deficient mice showed resistance to experimental autoimmune encephalomyelitis. IκB-ζ expression is supposed to contribute to these various pathological conditions. Therefore, 10H2DA may improve these pathological consequences via its activity in reducing IκB-ζ-induction and IκB-ζ–dependent gene-expression.
A schematic of the effect of 10H2DA on LPS-stimulated gene expression. LPS induces expression of NF-κB-dependent genes, such as TNF-α and JE, through MyD88/NF-κB signaling pathway. IκB-ζ is one of the NF-κB-dependent genes and induces the IκB-ζ-dependent genes including IL-6, lipocalin-2 and G-CSF. 10H2DA acid inhibits only a subset of NF-κB, which activates IκB-ζ promoter and is suspected to be in a different state of phosphorylation or other post-translational modifications.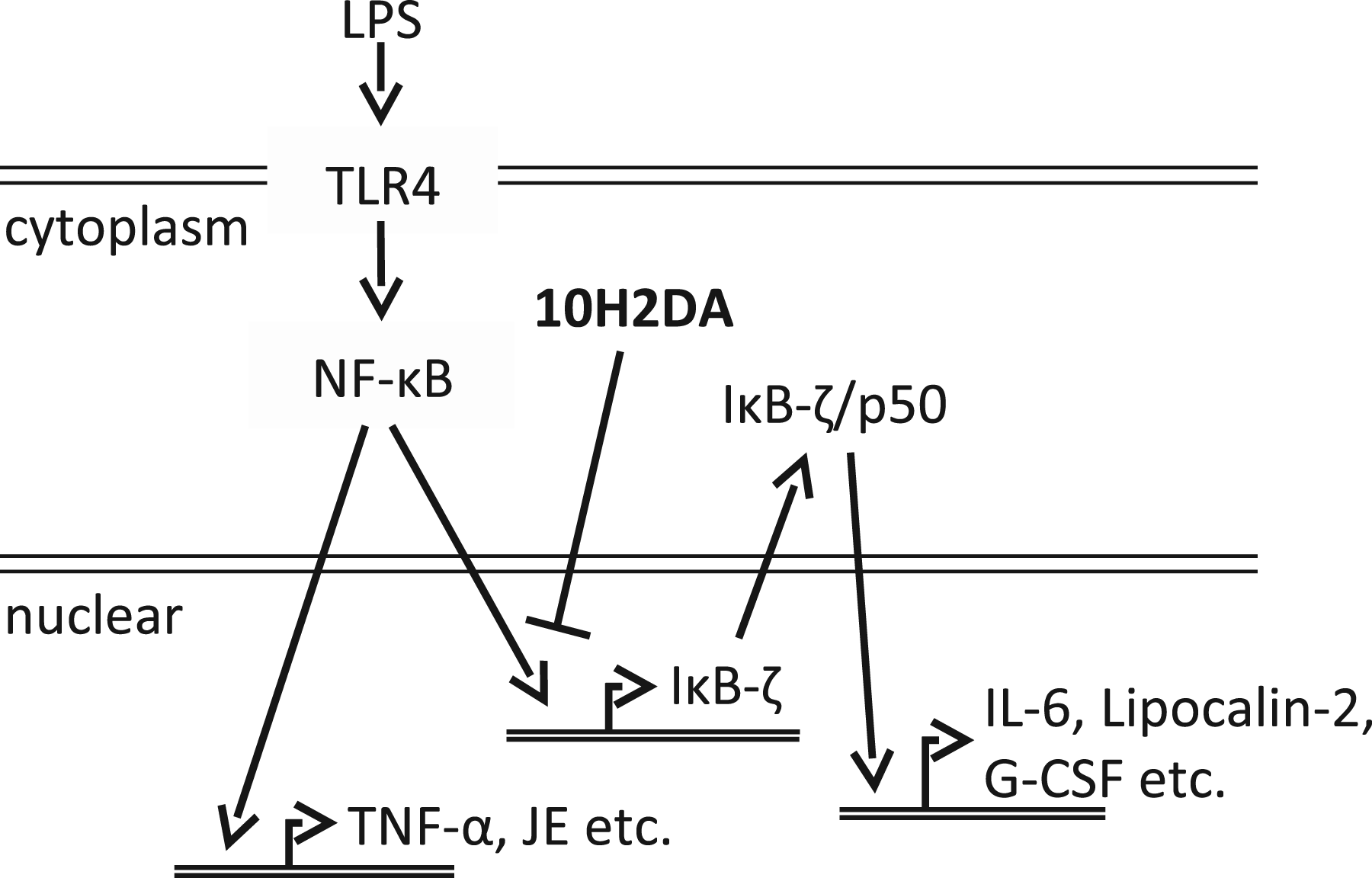
TNF-α and IL-6 are known to be major pro-inflammatory cytokines induced by MyD88-dependent LPS signaling. 27 NF-κB is a well known transcription factor responsible for IL-6 production. 28 In LPS stimulation, MyD88-dependent NF-κB activation is thought to be responsible for IκB-ζ gene expression. 5 Using a reporter gene assay, 10H2DA inhibited NF-κB activation under LPS and MyD88 induction. In RT-PCR analysis, however, 10H2DA did not affect the production of cytokines or chemokines, including TNF-α and JE, whose expression is supposed to be dependent on NF-κB. Furthermore, promoter activation of IκB-ζ, but not the TNF-α gene, was inhibited by 10H2DA in reporter gene assay. The NF-κB reporter construct that we used has four repeats of the consensus κB sequence, which is identical to that in the promoter region of the IκB-ζ gene. The κB sequence-specific binding of a subset of NF-κB complexes may be the reason that expression of IκB-ζ, but not the other NF-κB-dependent genes, was inhibited by 10H2DA. In fact, we could not detect the inhibitory effect of 10H2DA on the degradation of IκB-α. LPS-stimulated IKK-α phosphorylation was even increased by adding 10H2DA. We could also not detect the inhibition of NF-κB p65 nuclear translocation by 10H2DA (data not shown). These data suggest that at least part of NF-κB was released from the IκBs and translocated into the nucleus in LPS stimulation. Therefore, some NF-κB-dependent genes could be transcribed.
NF-κB activation is controlled by IKKs which are required for phosphorylation-induced degradation of IκBs. 3 Following ligation of TLRs, IKKs are activated by phosphorylation, resulting in IκB phosphorylation and degradation. 3 We could not detect significant inhibition of IκB degradation by 10H2DA, suggesting that NF-κB was at least released from the IκB–NF-κB complex. Full activation of NF-κB needs several post-translational modifications, including phosphorylation. 4 Activation of p65, a subunit of NF-κB activated in LPS stimulation, is believed to be regulated by phosphorylation of at least four Ser residues. Phosphorylation at Ser 276 is critical for interaction with the transcriptional co-activators cAMP response element-binding protein (CREB)-binding protein and p300, 29 and is therefore necessary for NF-κB–dependent gene transcription. The other phosphorylation sites may affect specific gene expression. Anrather et al. 30 reported that phosphorylation of Ser residues at 205, 276 and 281 is necessary for the expression of MIP-2 and ICAM-1, whereas IL-6 and vascular cell adhesion molecule (VCAM)-1 are expressed in a phosphorylation-independent manner. On the otherhand, Ser 536 needs to be phosphorylated to produce IL-6 in angiotensin II-stimulated vascular smooth muscle cells. 31 They also reported significant amounts of NF-κB were present in the nucleus; however, unphosphorylated NF-κB does not activate gene transcription, even in the nucleus. Although we could not detect an inhibitory effect of 10H2DA on LPS-induced phosphorylation of p65 at Ser 536 (data not shown), regulation of p65 phosphorylation at specific Ser residues may allow modulation of particular gene expression, including the IκB-ζ gene.
Kohno et al. 10 showed that royal jelly inhibited LPS- and IFN-γ–induced cytokine production, and identified one of the major protein components, MRJP3, as an anti-inflammatory factor derived from honeybee royal jelly. 10 They also suggested that royal jelly contains another anti-inflammatory factor with low molecular mass that inhibits TNF-α and IL-6 production. Although we could not detect an inhibitory effect on LPS-induced TNF-α production, 10H2DA may be one of the low molecular weight anti-inflammatory factors in royal jelly.
We showed that 2–4 m
In conclusion, we demonstrated that 10H2DA inhibited LPS-induce IκB-ζ production followed by gene expression of IκB-ζ–dependent genes, including IL-6. This is, as far as we know, the first report of a specific inhibitor for IκB-ζ induction. Accumulating evidence that IκB-ζ and IL-6 contribute to inflammatory and autoimmune diseases suggests that 10H2DA may be a therapeutic drug candidate for IκB-ζ– and IL-6–dependent disorders.
Conflicts of interest
None declared.
Footnotes
Acknowledgements
We thank Dr Takaoka and Dr Komatsu for providing the plasmids. This work was supported by a grant from the Japan Royal Jelly Fair Trade Council (Tokyo, Japan) and partly by Grant-in-Aid for Challenging Exploratory Research from the Japan Society for the Promotion of Science [21659110 to TS].