
Abstract
It has already been shown that tetanus toxoid (TTd) hyper-immunization is a suitable experimental method for creating the animal model of antiphospholipid syndrome (APS) in BALB/c mice. The severity of APS pathology in BALB/c mice mainly correlates to the affinity of anti-β2 glycoprotein I (β2GPI) antibodies. In this study we have investigated reproductive pathology induced in C57BL/6 mice by TTd hyper-immunization using a combination of different pretreatments (complete Freund's adjuvant or glycerol) and adjuvants (alhydrogel or glycerol). A decrease in fecundity was recorded in only C57BL/6 mice immunized with alhydrogel adjuvant, irrespective of the kind of applied pretreatment; it was associated with an increase in abundance of low affinity anti-β2GPI IgG antibodies and Th1 prevalence.
Introduction
Antiphospholipid syndrome (APS), also known as Hughes' syndrome, is an autoimmune disease with clinical manifestations of thrombocytopenia, recurrent fetal loss and repeated thromboembolic phenomena. 1 In addition to these clinical manifestations, it can include deep vein thrombosis, livedo reticularis, stroke and obstetrical complications, recorded as early or late miscarriages. Manifestations of APS 2 in the mouse model include a decrease in fecundity and/or fertility, together with delivery of fewer pups of lower body weight. It is considered that this disease appears as a result of an increase in anti-phospholipid (aPL), anti-β2 glycoprotein I (β2GPI) Abs and/or β2GPI-dependent aPL Abs. β2 glycoprotein I, also designated as apolipoprotein H, is a plasma glycoprotein that binds various kinds of negatively charged phospholipids (PLs), lipoproteins and activated platelets, and is used as a cofactor in aPL Abs binding. Criteria for APS 3 diagnosis have been formulated and comprise aPL Abs, but further investigations showed that anti-β2GPI Abs also represent a valid criterion for APS diagnosis. 4
The etiology of APS is multifactorial, with microbial antigens recorded as its frequent triggers. 5 The most cited mechanisms connecting microbial antigen stimulation and pathogenic aPL Abs secretion are molecular mimicry and polyclonal cell activation. Strict distinction between these two in the generation of pathogenic aPL Abs is hardly to be sustained. Molecular mimicry, as the most likely mechanism, is characterized by cross-reactivity owing to structural similarity between pathogen and host epitopes. In addition to innate immunity activation, host-like epitopes of pathogens must induce an adaptive immune response with T cell involvement and consequent epitope spread as well. On the other hand, polyclonal cell activation, which is mostly dependent on innate immune receptor activation can, if persistent, generate molecular mimicry related consequences. 6
Tetanus toxoid (TTd) hyper-immunization has been shown to be a suitable experimental method for the induction of APS in BALB/c mice. 7 It was convincingly demonstrated that molecular mimicry between TTd and β2GPI is among the causes of APS induction. 8 However, not all individuals or mice develop APS equally, mostly owing to differences in genetic susceptibility to autoimmune diseases. 9 This was the main reason we wished to examine the induction of APS by TTd hyper-immunization in C57BL/6 mice, which are genetically different from the BALB/c mice strain. The main difference between BALB/c and C57BL/6 mice is represented by the Th response type, i.e. BALB/c mice are Th2-prone, while C57BL/6 mice are Th1-prone.
In addition to antigen characteristics and genetic background, activation of innate immunity also participates in APS induction in animals. Many of the adjuvants exert their effects through innate immunity receptors (e.g. TLRs).
10
Adjuvant stimulation of innate receptors very often results in the generation of anti-PL Abs, which, by themselves, can further stimulate these receptors (e.g. TLR411) or prevent their stimulation (e.g. prevention of oxidized PL binding to TLR212). By understanding the adjuvant effects
13
in different systems various combinations of pretreatment and adjuvants were used to define the optimal conditions for the induction of APS in C57BL/6 mice in this study. Complete Freund's adjuvant (CFA) is a mixture of TLR agonists (lipid A, N-acetylmuramyl-
In this article we present results of the experiments performed in order to reveal possible mechanisms involved in the induction of reproductive pathology in C57BL/6 mice by TTd hyper-immunization. We evaluated biological activity of anti-β2GPI Abs by determining their capacity to produce reproductive pathology. Here we describe that both high- and low-affinity anti-β2GPI IgG Ab populations are present in C57BL/6 mice hyper-immunized with TTd, the latter being responsible for lowering fecundity.
Materials and methods
Immunization and bleeding schedules
Ten-week-old C57BL/6 female mice were used in the experiments. Immunization and bleeding schedules were as previously described (Figure 1).
6
Briefly, mice were immunized with TTd (Institute of Virology, Vaccines and Sera – Torlak, Belgrade, Serbia) mixed with either 2.5 Experimental design. Applied immunization, bleeding schedules and mating time-point are indicated with arrows. Single CFA or Glyc dose (200 µl/mouse) was applied as pretreatment (doted black arrow) one week before TTd immunization start (solid black arrow). Mice were bled before any manipulation (day – 1), one week following pretreatment and later on in two-week intervals (gray solid arrow). The TTd hyper-immunized female mice were mated one day after the third TTd dose (doted gray arrow).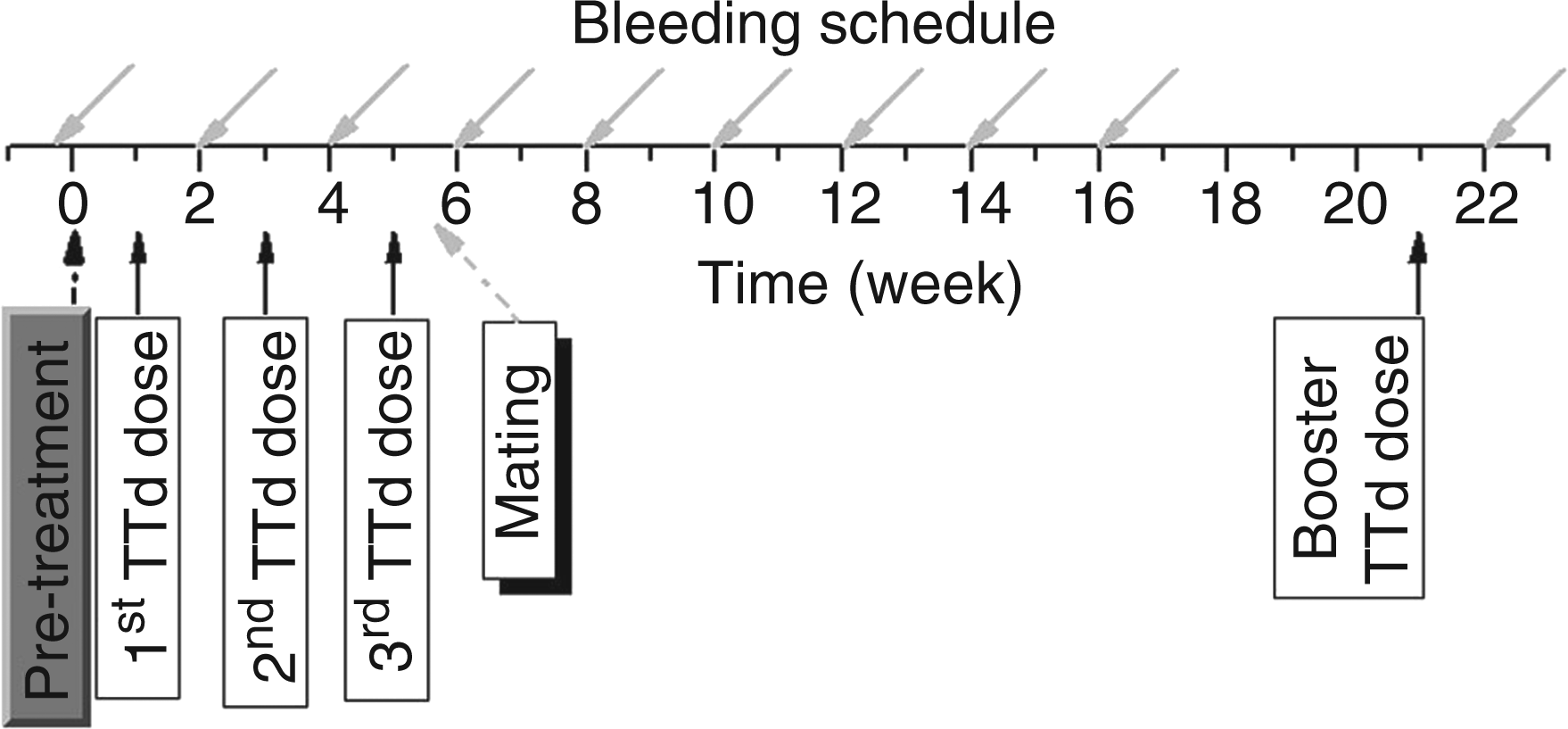
All animal experimentation was conducted in accordance with the local ‘Guiding Principles for the Care and Use of Laboratory Animals’, in agreement with the provisions of the Declaration of Helsinki, and was approved by the Animal Institutional Care and Use Committee at the Institute of Virology, Vaccines and Sera – Torlak.
Isolation of β2-GPI from human plasma
β2 glycoprotein I, which was used for serological tests, was isolated from human plasma as already described. 6 Commercially available mouse anti-human β2-GPI IgG1 (clone 5F7; ICN Biomedicals, Aurora, IL, USA) was used for identification of the isolated protein.
Indirect ELISA for protein antigens
Serum Abs specific for TTd (1 µg/ml TTd in PBS, 50 μl/well) or β2GPI (10 µg/ml β2GPI in PBS, 50 μl/well) were determined by ELISA. Plates (MaxiSorp, NUNC, Roskilde, Denmark) were saturated with 1% BSA. Sera samples were diluted 1:800 for detection of anti-TTd, and 1:500 for detection of anti-β2GPI Abs (prepared in 1% BSA/0.05% Tween). The binding of mouse sera Abs was detected using biotin- conjugated anti-mouse IgM (Cat. No. B-9265; Sigma, St. Louis) IgG (Cat. No. B-7401; Sigma, Steinheim, Germany), anti-IgG1 (Cat. No. 63-581, ICN), anti-IgG2b (Cat. No. 63-583, ICN), anti-IgG2c (115-065-208, Jackson ImmunoResearch, West Grove, PA, USA) and anti-IgG3 (MCA423B, Serotec, Kidlington, Oxford, UK), according to the manufacturer's instructions. In all cases, the ExtrAvidin-peroxidase/o-phenylenediamine (OPD) system (Sigma) was used to visualize Ag–Ab interactions. Absorbance was monitored at 492 and 620 nm (OD492/620). Serum samples were diluted in 1% BSA/0.05% Tween 20/PBS.
ELISA for high affinity anti-β2GPI Abs
The ELISA was performed as described by Pullen et al.
15
using several modifications. The procedure was similar to that given for anti-β2GPI specific ELISA with the difference that two microtiter plates precoated with β2GPI were used. After incubation with mouse sera (diluted 1:500 in 1% BSA/PBS) and subsequent washing with 0.05% Tween 20/PBS, one plate was incubated with 1
Anti-cardiolipin ELISA
Wells of 96-well ELISA plates (PolySorp, NUNC, Roskilde, Denmark) were coated with 50 µl of cardiolipin (CL) [Sigma; 30 µg/ml in ethanol)] by overnight evaporation. 6 For determination of β2GPI- dependent anti-CL Abs, we performed an additional incubation with β2GPI (50 µl/well, 10 µg/ml) for 1 h at room temperature. A 1% BSA/PBS solution was used for saturation for 2 h at room temperature. The saturation, and each subsequent ELISA step, was followed by washing with 0.05% Tween 20/PBS (4 × 200 µl/well). Diluted sera (1:800 in 1% BSA) were added to the plate and incubated for 1 h at room temperature. All other details remained identical to the ELISA described above.
Mice pregnancy models
The main reason for choosing the C57BL/6 mouse strain was that it is not lupus-prone, i.e. these mice are not genetically predisposed to autoimmune diseases.
One day after the third TTd dose, immunized and control females were mated with previously isolated males. The presence of a vaginal plug indicated successful breeding. After that, females were housed separately from males and pregnancy was allowed to proceed until term, when the number of live pups (fertility) and fecundity were recorded.
Statistical analysis
Student's t test for independent groups was used to compare fecundity, fetal resorption rates and number of live pups following TTd and PBS application. The group treated with PBS was used as the reference. A P-value of 0.05 was considered as the limit of statistical significance.
Results
Mouse reproductive pathology
Fecundity was defined as the percentage of pregnant females. In our system, fecundity was found to be mainly dependent on the adjuvant used for antigen application (Figure 2A). Thus, TTd immunization with alhydrogel induced a significant decrease of fecundity, while immunization with glycerol had no influence. Fecundity was not affected by the type of pretreatment.
Fecundity (A) and fertility (B) of C57BL/6 mice mated 1 d after the third tetanus toxoid (TTd) dose. Depending on the applied immunization protocol, female mice were divided into six groups, each containing 10–12 mice. Non-treated female mice (n = 11) were used as a normal age-matching control. Fecundity is expressed as mean percentage ± SE of pregnant mice within assigned group calculated from the results of two separately performed experiments. In both experiments, number of pups has been recorded and fertility was expressed as a mean number of pups per pregnant female of the corresponding group ± SE. The statistical significance of the observed differences was determined by t-test using the normal age-matched control group as reference (P < 0.05*, P < 0.005**). Al, alhydrogel; Glyc,2.5 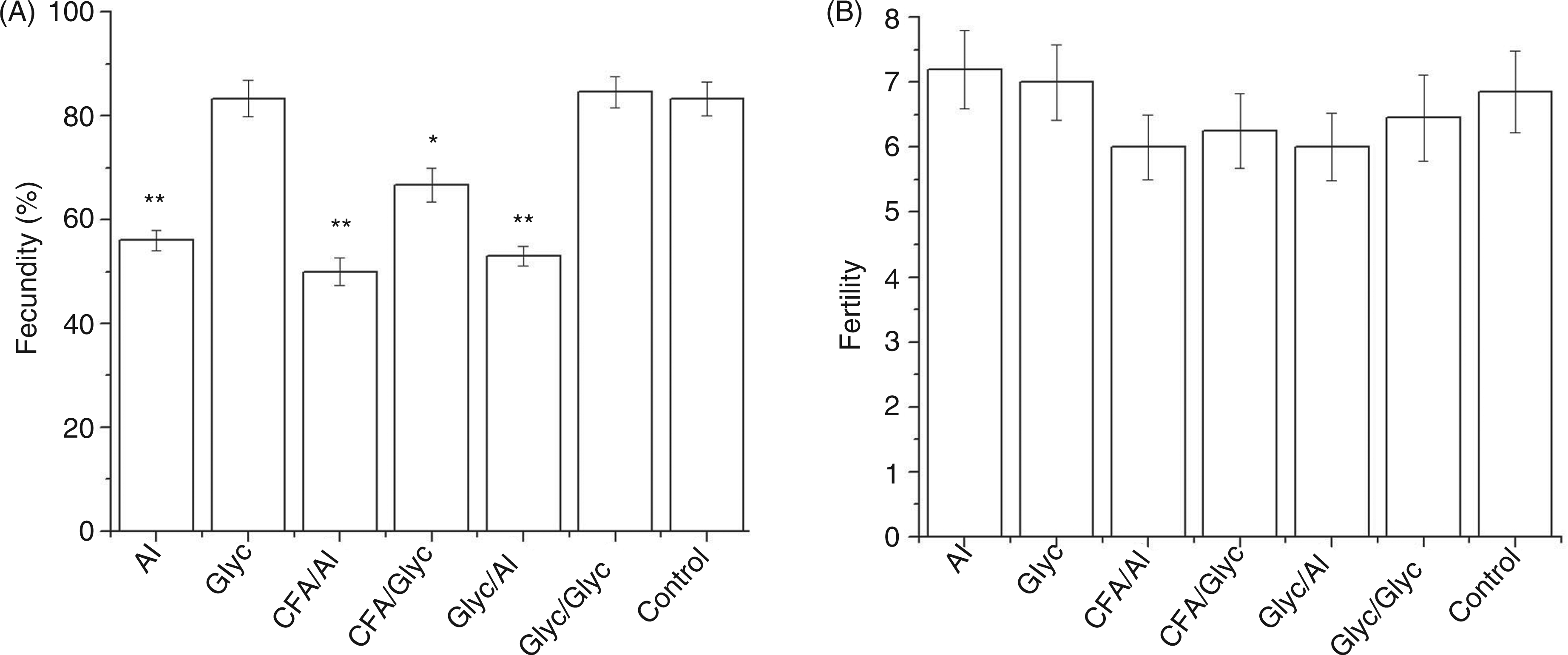
Fertility was defined as the average number of pups per pregnant female. We ascertained that TTd hyper-immunization generally had no impact on the fertility of C57BL/6 mice (Figure 2B).
Anti-CL IgG after tetanus toxoid (TTd) hyper-immunization
Anti-CL Abs are the golden diagnostic standard for APS.
16
Immunized mice were mated after the third TTd dose, i.e. 4–5 wk after the start of the procedure. This was the reason why we tested sera for the presence of anti-CL Abs of the IgG isotype. We have analyzed dynamics of specific antibodies induced during immunization course by following their frequencies and amplitude (levels), which represent simultaneously measured absorbance of corresponding sera, each diluted 1:800, at different time points, and for different groups. The highest levels of anti-CL Abs were recorded in CFA pretreated groups; levels were lower in groups without pretreatment (Figure 3) and the lowest in glycerol pretreated mice (Figure 3). Briefly, we have shown that anti-CL IgG fluctuations depend on the type of pretreatment used. This type of dependence cannot define anti-CL Abs as pathological, as the recorded reproductive pathology was adjuvant dependent (Figure 2).
Fluctuation in the level of anti-CL IgG antibodies in the sera of C57BL/6 mice during immunization course (until the sixth week), post-immunization follow-up period (until the sixteenth week) and after booster TTd dose application (twenty-second week). Non-pretreated (A), CFA-pretreated (B) and glycerol (Glyc)-pretreated (C) mice were immunized with TTd mixed with Al (black line) or 2.5 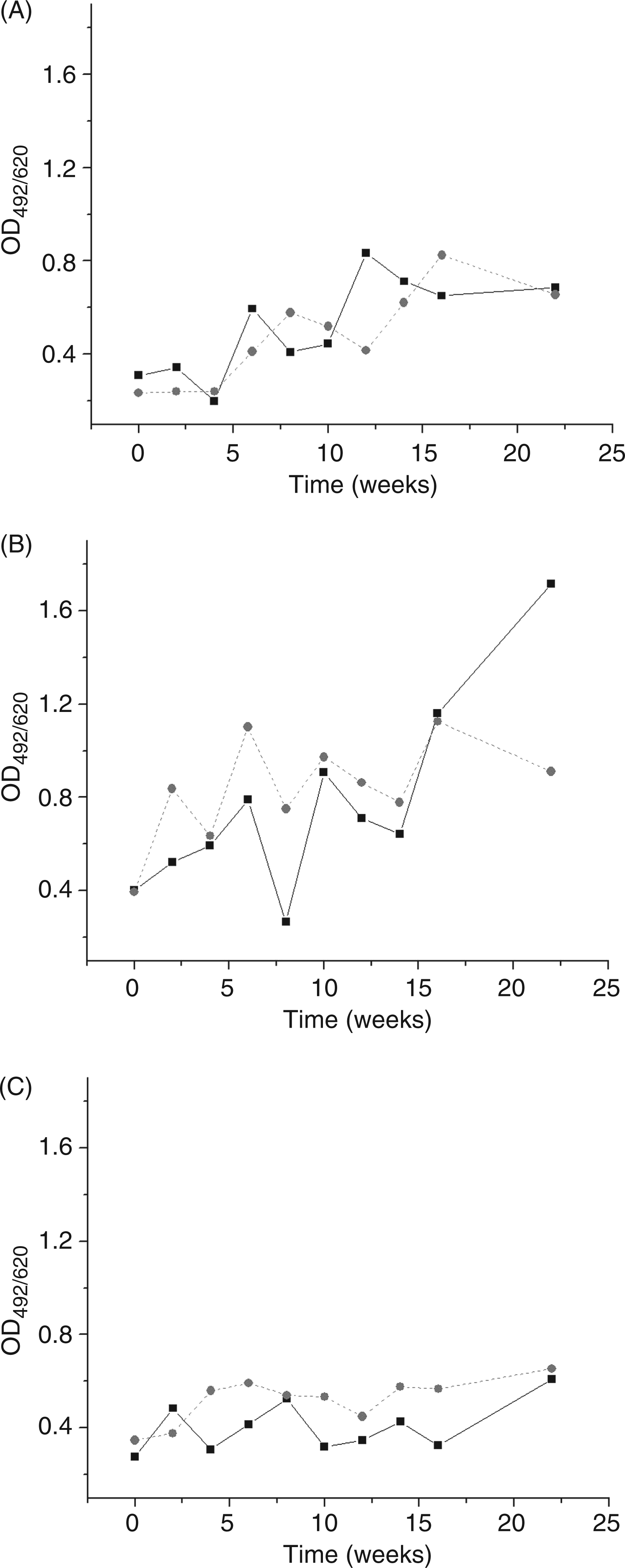
Anti-β2GPI IgG after tetanus toxoid (TTd) hyper-immunization
In parallel with anti-CL Abs, we analyzed anti-β2GPI IgG Abs. The TTd hyper-immunization led to a rise of anti-β2GPI IgG level for all immunization groups. This was observed after the first TTd dose; however, the extent differed between the experimental groups.
Titers of anti-β2GPI IgG in sera collected from TTd immunized C57BL/6 mice one week after the third TTd dose application. Titer was defined as the lowest sera dilution in which concentration of anti-β2GPI IgG was the same as in sera collected before any treatment (normal mice sera).

Al, alhydrogel; Glyc, glycerol; CFA, Complete Freund's adjuvant.
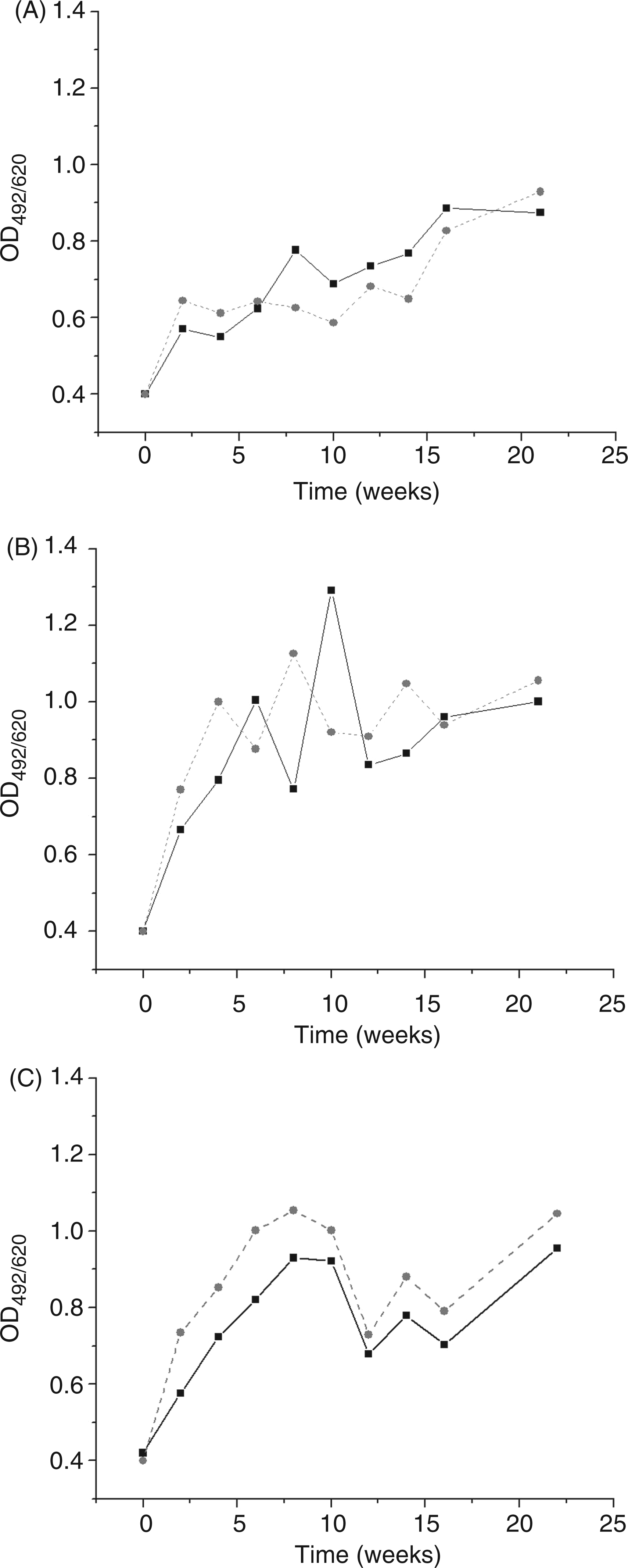
Fluctuation in the level of anti-β2GPI IgG Abs in the sera of C57BL/6 mice during immunization course (until the sixth week), post- immunization follow-up period (until the sixteenth week) and after booster TTd dose application (twenty-second week). Non-pretreated (A), CFA-pretreated (B) and Glyc-pretreated (C) mice were immunized with TTd mixed with Al (black line) or 2.5
Anti-CL/β2GPI IgG after tetanus toxoid (TTd) hyper-immunization
According to the available literature data, the serological picture of APS can include anti-β2GPI Abs reactive with epitopes present in native β2GPI, as well as against the cryptic epitope available upon binding of β2GPI to anionic surfaces such as CL.
17
Therefore, we tested anti-β2GPI, anti-CL and anti-CL/β2GPI-dependent IgG Abs concomitantly (Figure 5). In order to simplify presentation of the results, only those results obtained from CFA/Al treated mice are shown. The findings for other experimental groups are in accordance with them. The data are presented as a comparison of the results obtained at the beginning of the procedure, and 1 day after the third TTd dose (mating time). In our system only anti-β2GPI and anti-CL IgG Abs exhibited statistically significant increases when compared with control sera. There were no significant changes in the amounts of IgM Abs with either tested specificity (Figure 5).
Levels of IgM and IgG antibodies specific for CL (A), β2GPI/CL (B) or β2GPI (C) in the sera of CFA/Al TTd-immunized C57BL/6 mice (n = 10) collected before any intervention (0 wk, white columns) and after immunization completion (sixth week, gray columns). All sera samples were assayed in triplicate for specific reactivity using ELISA, and mean OD492/620 ± SE are presented. The statistical significance of difference between levels of specific sera antibodies before and after CFA/Al TTd-immunization was determined by t-test (P < 0.05*, P < 0.005**).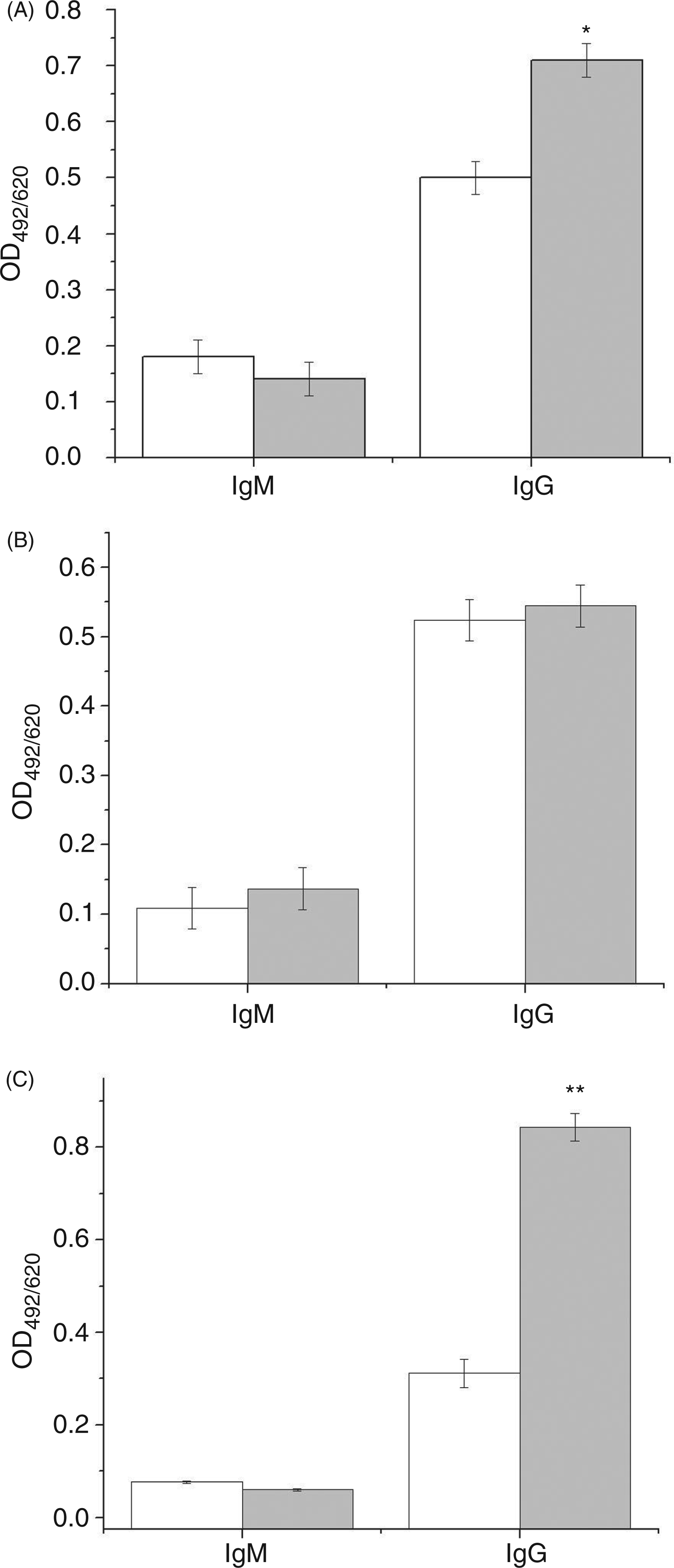
Affinity of anti-β2GPI antibodies
Low- and high-affinity anti-β2GPI Abs were analyzed by desorption of low affinity Abs with KSCN. The relative amounts of these Abs were calculated as the difference between the total quantity of anti-β2GPI Abs (detected after incubation with PBS) and the amount of high affinity anti-β2GPI Abs (detected after incubation with KSCN), obtained in ELISA. The results indicate fluctuations of low-affinity anti-β2GPI Abs in the sera of treated animals (Figure 6). It is noticeable that the levels and fluctuations of low affinity anti-β2GPI Abs are adjuvant dependent. Sera from all animals treated with Al had raised relative amounts of low-affinity anti-β2GPI Abs during the period of mating, when compared to values at the beginning of the immunization procedure.
Time-dependent fluctuations in abundance of low affinity anti-β2GPI IgG estimated as the difference between the absorbance registered for wells incubated with PBS (OD492/620(PBS)) and the absorbance registered for wells incubated with 1 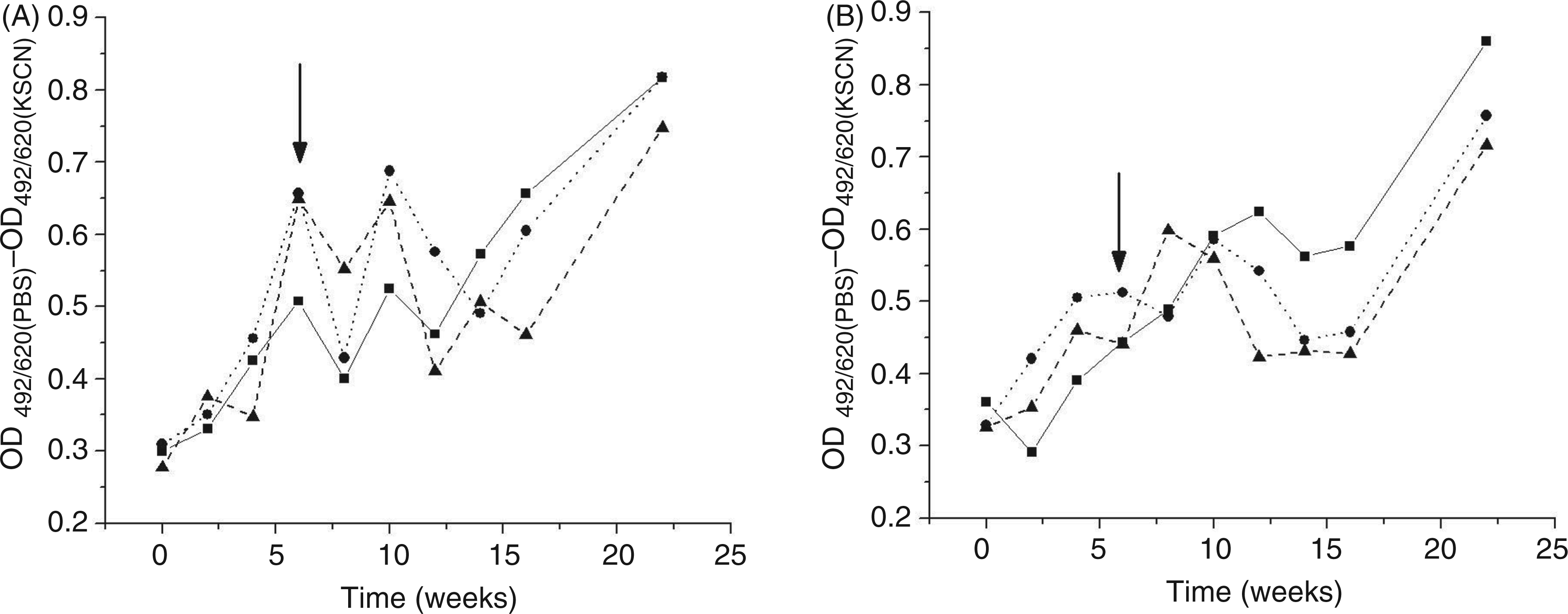
Anti-tetanus toxoid IgG
Hyper-immunization with TTd led to the production of TTd specific Abs in all experimental groups, with intensity dependent on the applied adjuvant (Figure 7). We also found that Al is a more potent adjuvant in TTd Ab production than glycerol, which confirms findings in previous studies.
18
Time-dependent fluctuations in the level of sera anti-TTd IgG in C57BL/6 mice immunized with TTd. Panels A and B show results obtained for non- (solid line), CFA- (dotted line) or Glyc-(dashed line) pretreated mice immunized with TTd in Al and Glyc adjuvant, respectively. Pools of sera obtained at the defined time-point from mice immunized according to the same protocol (n = 10) were used as samples. All samples were analyzed simultaneously in triplicate using ELISA.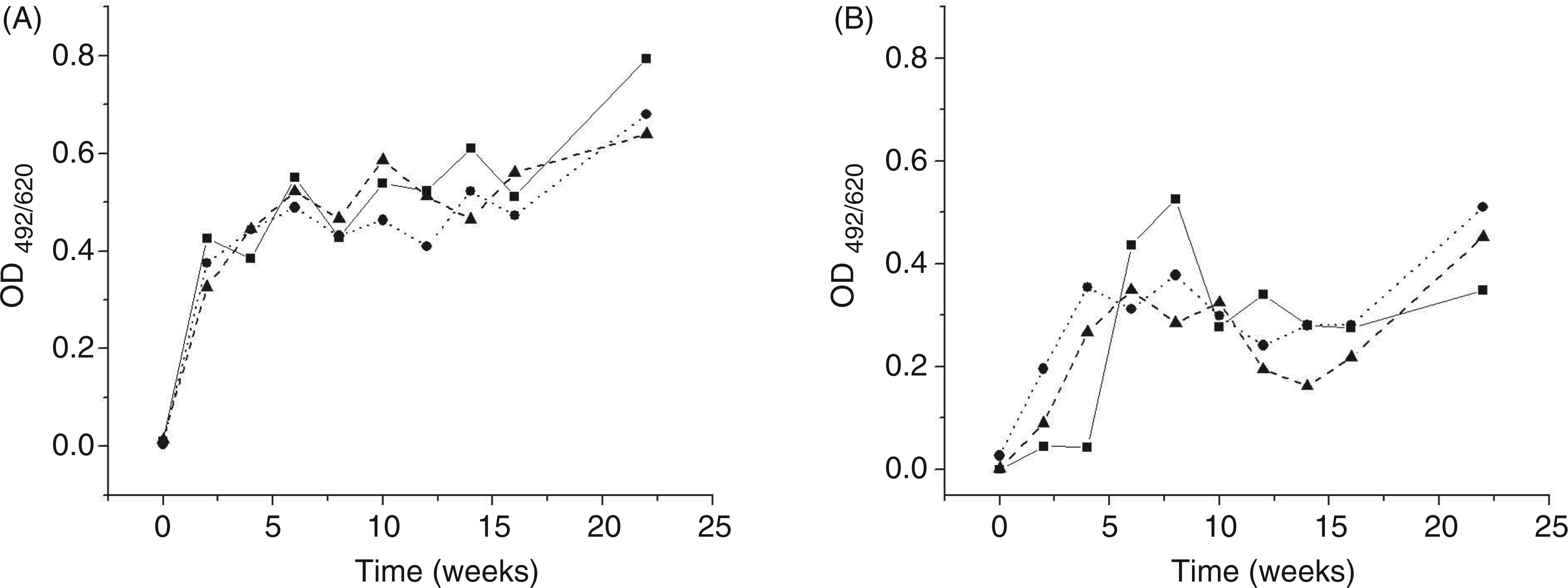
Th prevalence
We determined the presence of different TTd specific IgG subclasses in the sera of mice from each experimental group. The pattern of subclasses within the IgG pool indirectly represents skewing of the Th1/Th2 immune response. Secretion of IgG1 Abs is primarily induced by Th2-type cytokines, whereas production of IgG2c in C57BL/6 mice reflects the presence of Th1-type cytokines. The IgG1/IgG2c ratios within the specific IgG pool from sera of TTd hyper-immunized mice are shown in Figure 8. At the time of mating, the Th2 response was much lower in Al treated animals, resulting in predomination of the Th1 type response.
Discussion
In this study, we investigated the relationship between pretreatments and adjuvants used in TTd hyper-immunization, and the reproductive pathology induced in C57BL/6 mice. It has been shown that TTd-hyper-immunization procedures comprising Al as adjuvant,lead to the reproductive pathology, actually decreasing fecundity. Our results also show the influence of applied immunization protocols on characteristics of Abs implicated in APS pathology.
Ratio of IgG1/IgG2c within TTd-specific IgG pool (used for evaluation of Th2/Th1 prevalence) in sera of TTD-immunized C57BL/6 mice collected at the defined time points. Results are presented as ratio of TTd-specific IgG1 and IgG2c OD492/620 recorded by ELISA. Panels A and B show results obtained for non- (soild line), CFA- (dotted line) or Glyc-(dashed line) pretreated mice immunized with TTd in Al and Glyc adjuvant, respectively. Pools of sera obtained at the defined time-point from mice immunized according to the same protocol (n = 10) were used as samples. Arrows indicate time point of mating.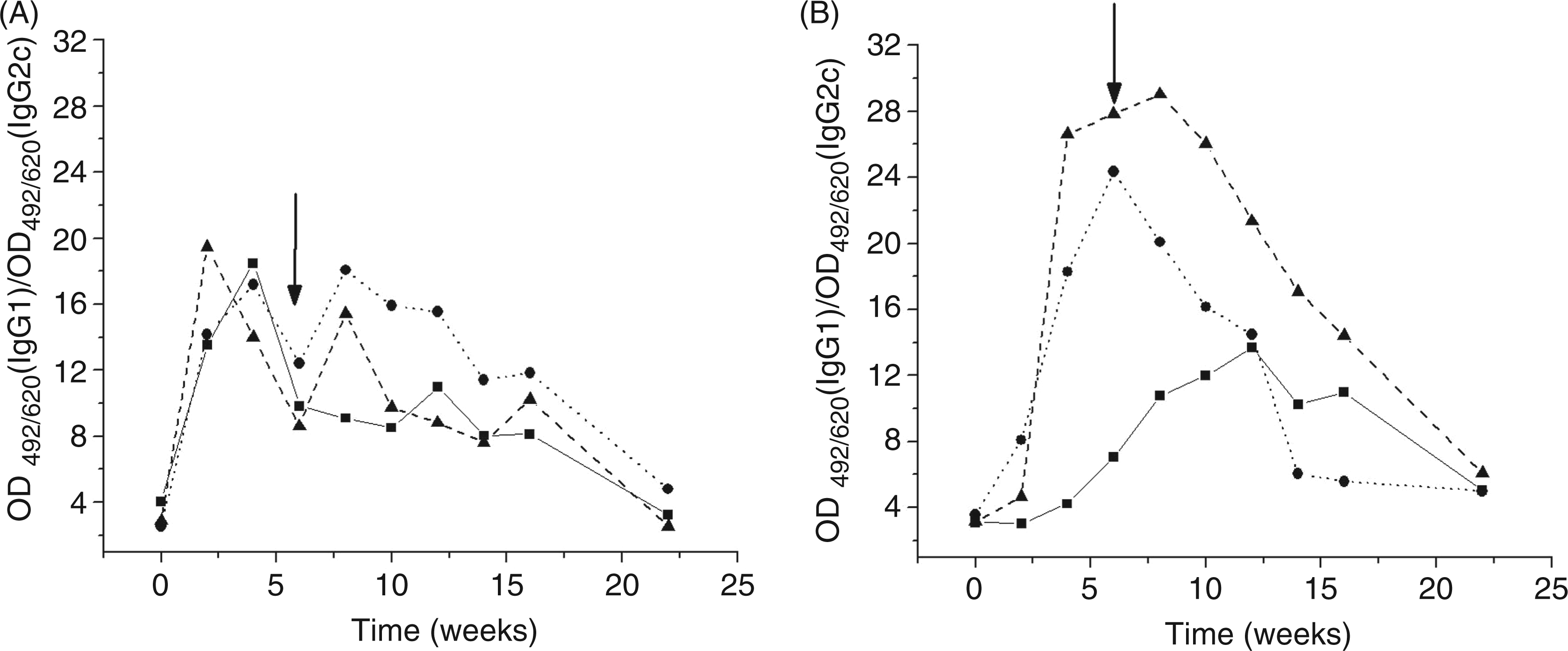
Numerous studies on mice have shown that pathogenic Abs in APS cause pregnancy loss by binding to PLs expressed on the invading trophoblast, thus inhibiting successful embryonic implantation into the endometrium. Additionally, after placentation is established, their thrombogenic action leads to decreased placental perfusion and subsequent infarctions. 19 In the animal model presented, TTd hyper-immunization of C57BL/6 mice leads to decreased fecundity, but not fertility, indicating that the observed reproductive pathology is probably caused by a defect in the process of implantation and not by placental infractions. In accordance with these results are our findings that TTd hyper-immunization does not lead to fetal resorption or thrombocytopenia in C57BL/6 mice. 20
The influence of applied TTd-hyper-immunization protocol on changes within aPL Abs pool was examined by three ELISA-based systems utilizing CL (alone or complexed with human β2GPI) or human β2GPI alone as antigens.
It is well known that detection of anti-CL Abs is the standard for laboratory diagnostics of APS. 15 However, the increases in anti-CL IgG levels was observed in all experimental groups, so the recorded decrease in fecundity in Al treated groups could not be connected to these Abs. In addition, the dynamics of the anti-CL Abs pool were pretreatment-dependent, while the induction of reproductive pathology (decrease in fecundity) was adjuvant-dependent. Finally, these results are in accordance with data21,22 showing that infection-induced anti-CL IgG Abs, which are predominantly β2GPI independent, 23 are not pathogenic and do not cause thrombotic complications.
The measurement of β2GPI-dependent anti-CL IgG was performed as numerous literature data imply that these Abs are almost exclusively pathogenic. 24 The binding of β2GPI to PLs results in the exposure of a cryptic epitope on β2GPI, which is then recognized by β2GPI-dependent anti-CL Abs. However, we did not detect a rise in the concentration of anti-CL/β2GPI Abs, while at the same time the anti-CL and anti-β2GPI Abs were elevated. This observation can be explained by the presumption that CL and β2GPI are bound through an epitope recognized by anti-CL/β2GPI Abs. This possibility has been already suggested by other authors. 16
Recently it has been accepted that an increase in the amount of anti-β2GPI Abs is one of the most important laboratory markers for APS diagnostics. 4 However, these Abs are also present in the circulation under physiological conditions as constituents of the natural Ab pool in humans and mice. Moreover, there are reports that ‘infectious' anti-β2GPI Abs are non-pathogenic. All these facts contribute to the complexity of the research focused on the role of β2GPI-specific Abs in APS. Their presence in healthy individuals seems contradictory to the pathological role proposed for them in APS patients, unless differences in quality or quantity are not responsible for their pathological potential. Our observations regarding these Abs are similar to those for anti-CL Abs: the rise in anti-β2GPI IgG level due to TTd-hyper-immunization was recorded in all experimental groups, but its level was pretreatment-dependent. As the recorded decrease in fecundity was adjuvant dependent, it can be concluded that anti-β2GPI Abs are not connected to decreased fecundity. 25
One should bear in mind that the Ab level is dependent on both the amount and the affinity of Abs present in the serum, i.e. serum with a relatively large amount of low affinity Abs can have a similar Ab level to that of serum with a relatively small amount of high affinity Abs. Hence, measurement of total Abs without any distinction between high- and low-affinity populations can hide important information. It is generally accepted that anti-β2GPI Abs found under physiological condition are of low affinity, while only the high-affinity β2GPI-specific Abs may be pathologically relevant.
26
We consider that inclusion of a washing step with 1
As we used TTd as the antigen in our experiments, we indirectly evaluated systemic cytokine profiles at the time of mating by measuring the ratios of corresponding IgG subclasses within the TTd-specific sera IgG pool. Although Al is known as an adjuvant that induces a Th2 response, IgG1/IgG2c ratio recorded in the sera of mice immunized with Al-adsorbed TTd at the time of mating (after the third TTd dose), implied a dominant Th1 response, which could be connected to decreased fecundity. It has been already suggested that adjuvant can lead to an autoimmune type of illness, which was defined as “adjuvant disease”. 29 ’ 30 Al, the most widely used adjuvant in human medicine, was shown to stimulate the production and secretion of certain cytokines, and is also nowadays connected to human macrophagic myofasciitis syndrome. 9
On the contrary, the Th2 response greatly predominated in animals immunized with glycerol as the adjuvant. It has been demonstrated that women with recurrent fetal loss exhibit primarily Th1 cytokines systemically, whereas healthy women have less Th1 cytokines and more Th2 cytokines, implying a potential role for a dichotomous T-helper response in the mediation of subsequent reproductive events. There are very few data on the mechanism by which factors of the immune system act, but clear evidence shows that the balance between the Th1 and Th2 cytokines at the feto-maternal interface is extremely important for fetal survival. 31 Hence, it is possible that systemic Th2 prevalence is “healthy”, i.e. Th2 milieu after implantation results in unaffected fertility, while the Th1 profile at the time of mating induces a decrease in fecundity.
Altogether, we report that in TTd hyper-immunized C57BL/6 mice there is a decrease in fecundity, a rise in the level of low-affinity anti-β2GPI Abs, and Th2 prevalence depended on Al-induced mechanisms. Although CFA and Glyc used for pretreatment undoubtedly have some modulatory effects (CFA is a mixture of TLR agonists 32 , and Glyc is a chemical chaperone with T cell stimulatory activity 33 ), Al exerted the dominant influence on the above-mentioned properties. It is possible that Al can stimulate the production of low-affinity anti-β2GPI Abs independently on TLR activation of dendritic cells and T cell stimulation. The particulate form of Al enables it to act not through TLR on dendritic cells, but through direct cytotoxic effects, leading to necrosis and, in that way, probably to bystander activation. Additionally, Al enhances antigen uptake and presentation. 34 To answer the question of which mechanisms are involved in these “adjuvant assisted” phenomena, more sophisticated investigations need to be conducted.
Results presented in this article show that in C57BL/6 mice, TTd hyper-immunization with Al adjuvant leads to a decrease in fecundity, which is associated with changes in anti-β2GPI IgG affinity. However, wecan conclude that in C57BL/6 mice, TTd hyper-immunization with Al adjuvant leads to a decrease in fecundity, which can be very important in terms of infertility investigations in the human population, whether they are connected to APS or not.
Footnotes
Funding
This work was supported by the Ministry of Science and Technology, Republic of Serbia [grant number 172049].