
Abstract
Inhibition of adipocyte triglyceride biosynthesis is required for fatty acid mobilization during inflammation. Triglyceride biosynthesis requires glycerol 3-phosphate and phosphoenolpyruvate carboxykinase (PEPCK) plays a key role. We demonstrate that LPS, zymosan, and TNF-α decrease PEPCK in liver and fat. Turpentine decreases PEPCK in liver, but not in fat. The LPS-induced decrease in PEPCK does not occur in TLR4 deficient animals, indicating that this receptor is required. The LPS-induced decrease in hepatic PEPCK does not occur in TNF receptor/IL-1 receptor knockout mice, but occurs in fat, indicating that TNF-α/IL-1 is essential for the decrease in liver but not fat. In 3T3-L1 adipocytes TNF-α, IL-1, IL-6, and IFNγ inhibit PEPCK indicating that there are multiple pathways by which PEPCK is decreased in adipocytes. The binding of PPARγ and RXRα to the PPARγ response element in the PEPCK promoter is markedly decreased in adipose tissue nuclear extracts from LPS treated animals. Lipopolysaccharide and zymosan reduce PPARγ and RXRα expression in fat, suggesting that a decrease in PPARγ and RXRα accounts for the decrease in PEPCK. Thus, there are multiple cytokine pathways by which inflammation inhibits PEPCK expression in adipose tissue which could contribute to the increased mobilization of fatty acids during inflammation.
Introduction
Infection, inflammation, trauma and malignancy induce the acute phase response, which is characterized by increases in the levels of certain plasma proteins (positive acute phase response proteins; for example C-reactive protein and serum amyloid A) and decreases in the levels of other plasma proteins (negative acute phase response proteins; for example albumin and apolipoprotein A1).1,2 Diabetes, obesity, atherosclerosis and a large number of other common clinical disorders are now also recognized to induce a mild acute phase response. 3 – 6 The changes in plasma protein levels during the acute phase response are often accounted for by changes in the synthesis of specific proteins by the liver, an effect that is mediated by cytokines such as TNF-α, IL-1, IL-6, and interferons.1,2
However, it is well recognized that the acute phase response not only results in changes in the pattern of proteins synthesized by the liver, but also in marked alterations in the expression of specific proteins synthesized in extrahepatic tissues. As a consequence of these changes in protein synthesis, the acute phase response also induces alterations in carbohydrate and lipid metabolism.7,8 The acute phase response is characterized by a decrease in serum high-density lipoprotein (HDL) levels and an increase in serum triglyceride (TG) levels. 7
There are a number of mechanisms that contribute to the increase in serum TG levels during inflammation. 7 First, the expression and activity of lipoprotein lipase decreases during inflammation, which results in a delay in the clearance of TG rich lipoproteins.7,9– 13 Second, inflammation induces an increase in hepatic very low-density lipoprotein (VLDL) production, that is stimulated by both an increase in hepatic de novo fatty acid synthesis and by increased delivery of free fatty acids (FAA) from adipose tissue.7,10 Fatty acid oxidation in the liver is suppressed during inflammation, which increases the availability of fatty acids for hepatic TG synthesis.7,14,15 The increase in TG formation in the liver stimulates VLDL production and secretion. 7
During inflammation, an increase in TG lipolysis in adipose tissue causes the elevation of serum free fatty acid levels leading to increased delivery of fatty acids to the liver.7,10 Cytokines increase adipose tissue lipolysis mainly by post-translational phosphorylation of hormone-sensitive lipase and its associated protein, perilipin.7,16 Phosphorylation of hormone-sensitive lipase increases its lipolytic activity, whereas phosphorylation of perilipin induces the translocation of hormone-sensitive lipase to the lipid-rich droplet where lipolysis occurs.17,18 A decrease in perilipin protein content may serve as an additional mechanism for increased lipolysis. 19
Triglyceride synthetic enzymes are highly expressed in adipose tissue and the released free fatty acids may be quickly re-esterified. In addition to promoting lipolysis, inflammation also enhances free fatty acid mobilization by decreasing intracellular fatty acid re-esterification through decreases in the expression of triglyceride biosynthetic enzymes, including acyl-CoA synthetase, glycerol-3-phosphate acyltransferase, 1-acyl-glycerol-3-phosphate acyltransferase, lipin 1, monoacylglyceride acyltransferase and diacylglyceride acyltransferase. 20 – 22 Thus, inhibition of triglyceride biosynthesis in addition to activated lipolysis is required for effective fatty acid mobilization during inflammation.
A crucial step in the formation of TG is the synthesis of glycerol 3-phosphate, which can be formed either by phosphorylation of glycerol by glycerol kinase or by the reduction of dihydroxyacetone by glycerol 3-phosphate reductase.23,24 Glycerol kinase, while very active in the liver, is present at only low activity in adipose tissue. 25 Dihydroxyacetone can be formed from either glucose via glycolysis or from pyruvate via glyceroneogenesis. However, recent studies have shown that in adipose tissue, glyceroneogenesis is the major pathway accounting for approximately 90% of triglyceride glycerol synthesis.23,24,26 The key regulatory enzyme in the glyceroneogenesis pathway is the enzyme phosphoenolpyruvate carboxykinase (PEPCK), which has two separate isoforms: a cytosolic (PEPCK1) and a mitochondrial form (PEPCK2).23,24 In rodents, the cytosolic form is the predominant isoform. For example, in rodent liver 90 to 95% of PEPCK activity is accounted for by PEPCK1 activity. 27 Cytosolic PEPCK1 has been studied in great detail in mammals, while the function and regulation of PEPCK2 has not been extensively studied. 24 In birds, the PEPCK2 isoform predominates and is thought to play a key role in gluconeogenesis. 24
Previous studies have shown that LPS administration and the cytokines induced by LPS decrease PEPCK activity and PEPCK1 mRNA levels in the liver. 28 – 32 Because of the importance of PEPCK in gluconeogenesis, this decrease in hepatic PEPCK activity contributes to the hypoglycemia observed with severe sepsis. Although fasting increases PEPCK in both liver and adipose tissue, the regulation of PEPCK may be discordant in liver and adipose tissue, as is seen with glucocorticoid treatment.33,34 Because adipose tissue lipolysis is markedly stimulated during inflammation, we hypothesized that the expression of PEPCK in adipose tissue, which is essential for glyceroneogenesis, would also be markedly suppressed in order to enhance the release of free fatty acids.
Materials and methods
Materials
Lipopolysaccharide (Escherichia coli 55:B5) was obtained from Difco Laboratories (Detroit, MI, USA) and diluted in pyrogen-free 0.9% saline. Zymosan A and TRI-Reagent were from Sigma (St. Louis, MO, USA). Turpentine was purchased from BDH Laboratories (Poole, England). Recombinant mouse TNF-α, IL-1β, IL-6, and INF-γ were from R&D Systems (Minneapolis, MN, USA).
Animals
Female C57BL/6 mice (8∼12 wk of age) were obtained from Charles River Laboratories (Wilmington, MA, USA). C57Bl/10 ScNJ (TLR4 -/-), C57Bl/10J (controls), B6, and 129S-Tnfrsf1aTm1Imx IL1r1Tm1Imx/J mice were all obtained from Jackson Laboratory (Bar Harbor, ME, USA). The animals were maintained in a normal-light-cycle room and were fed Teklad 8656 rodent chow (Harlan Laboratories, Indianapolis, IN, USA) and water ad libitum. Animals were injected with either saline, LPS (0.005–5 mg/kg, ip), zymosan A (80 mg/kg, ip), Oil of Turpentine (100 µl sc in left hind leg), TNF-α (50 µg/kg, ip) or IL-1β (4 µg/kg, ip) and food was removed from both control and treated animals following injection whilst water was provided ad libitum. At the indicated time points, control mice (injected with saline) and treated mice were rapidly euthanized with an overdose of isoflurane, and liver, heart, kidney, diaphragm muscle, and adipose tissue from the periuterine-urinary bladder removed and snap-frozen in liquid nitrogen. Tissues were then placed in storage tubes in dry-ice bath until the end of experiment and then stored at −80°C until RNA extraction. All experiments were performed according to protocols approved by the Animal Studies Subcommittee of the San Francisco Veterans Affairs Medical Center.
Cell culture experiments
Murine 3T3-L1 cells (ATCC, Manassas, VA, USA) were grown to confluence and differentiated to adipocytes as described.
35
Briefly, pre-adipocytes were cultured in Dulbecco’s Modified Eagle Medium (DMEM) and 10% fetal bovine serum. When cells reached confluence, they were differentiated by treatment with 1 µg/ml insulin, 0.5 m
Experiments were performed on post-differentiation day 10. Cells were treated for 24 h with LPS (0.1–10 µg/ml), TNF-α (10 ng/ml), IL-1 beta (10 ng/ml), IL-6 (10 ng/ml) or INF-γ (100 ng/ml). The dose of cytokines used in these experiments are similar to those previously shown to induce metabolic alterations in 3T3-L1 adipocytes. 35
Free fatty acid assay
Serum was obtained at 16 h after LPS treatment and free fatty acid (FFA) was measured by an enzymatic method (WAKO Diagnostics, Richmond, VA, USA) according to the manufacturer’s instructions.
Isolation of RNA and cDNA preparation
Total RNA was isolated from tissues or cells by the TRI Reagent method from Sigma. First-strand cDNA was reverse-transcribed from 1µg of total RNA using iScript cDNA Synthesis Kit (Bio-Rad, Hercules, CA, USA).
Real-time quantitative PCR
The real-time quantitative PCR reaction contained 20 ng of reverse-transcribed total RNA, 450 n
Phosphoenolpyruvate carboxykinase activity
Phosphoenolpyruvate carboxykinase enzyme activity was measured by a
14
C radiometric method similar to that described by McCallum et al.
30
Briefly, tissues were homogenized in 0.25 m
Nuclear extract preparation and gel mobility shift assay
Nuclear proteins from adipose tissue were prepared using the method described by Musri et al.
36
For the gel shift assay, a double-strand oligonucleotide corresponding to the sequence of rPCK2 was end-labeled with [γ-
32
P] ATP using the Invitrogen KinaseMax kit (Invitrogen, Carlsbad, CA, USA). The oligonucleotide sequences used were: wt rPCK2, CAC AAC TGG GAT AAA GGT CTC GCT GCT; mt rPCK2, CAC AAC TGG GAT CCT GGT CTC GCT GCT. Gel shift assay was performed as previously described.
37
Briefly, nuclear extracts (10 µg) were first incubated for 15 min at room temperature with or without a 100 x molar excess of double-strand oligonucleotide in a reaction buffer consisting of 25 m
Statistical analysis
The results are presented as means ± SD. Statistical significance between two groups was determined by using the Student’s t-test. When multiple samples were compared, one-way (one independent variable) and two-way (two independent variables) ANOVA were used to determine statistical significance. A P value < 0.05 was considered significant.
Results
Our initial experiments examined the effect of the administration of LPS, a TLR4 activator, on PEPCK in multiple tissues. In previous LPS studies, we have shown no alterations in serum creatinine levels or liver function tests.38,39 However, serum free fatty acid levels are increased in LPS-treated animals (Control = 1.169 m The effect of LPS administration on phosphoenolpyruvate carboxykinase (PEPCK). Mice were injected intraperitoneally with LPS (5 mg/kg) and the animals were euthanized at 16 h after LPS administration. (A) PEPCK1 mRNA. (B) PEPCK2 mRNA. For mRNA quantification, total RNA was isolated, cDNA was synthesized with reverse transcriptase, and quantitative real-time PCR performed as described in the Materials and Methods. (C) PEPCK activity was measured as described in the Materials and Methods. In control animals, PEPCK activity in liver was 375 nmoles/15 min/mg protein, in heart 0.67 nmoles/15 min/mg protein and in adipose tissue 325 nmoles/15 min/mg protein. The data are presented as the mean ± SD. Data are expressed as a percentage of controls. N = 5 per group. *P < 0.05, **P < 0.01, ***P < 0.001.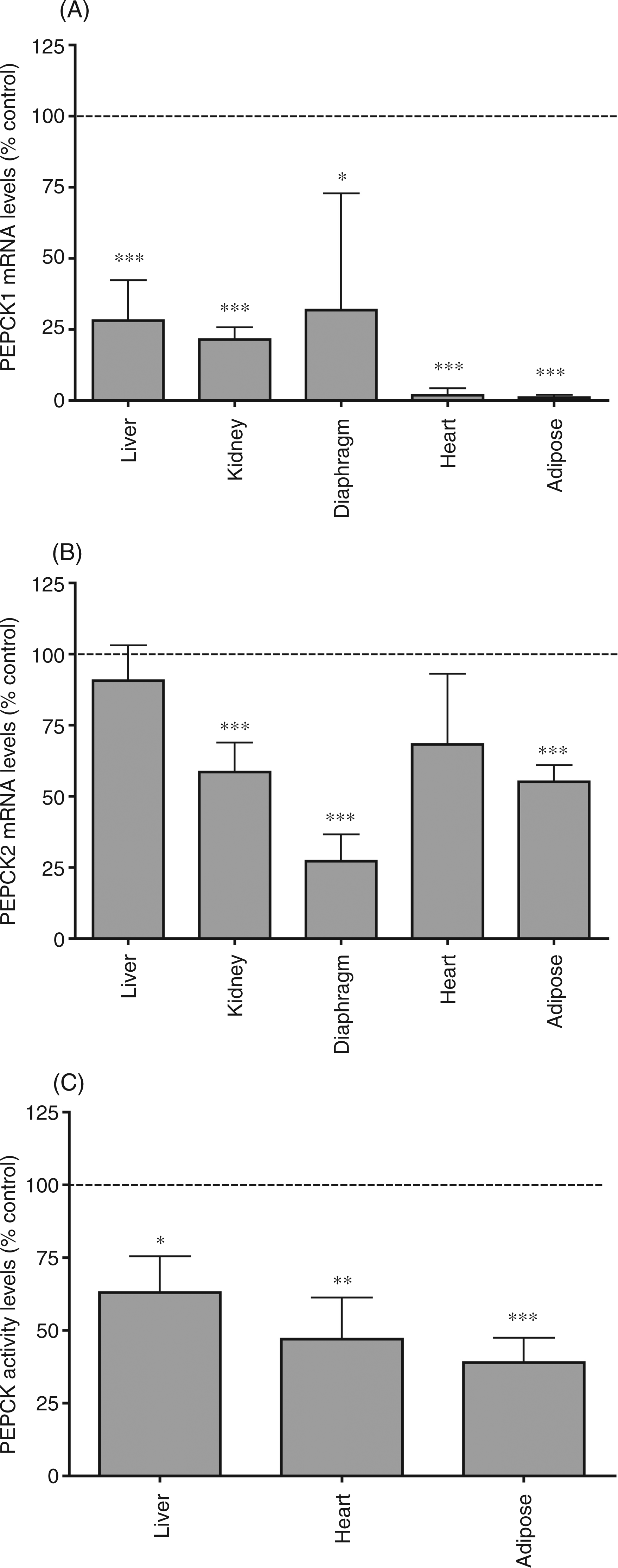
Given the important role of PEPCK in glyceroneogenesis in adipose tissue and that PEPCK1 is the major isoform expressed in adipose tissue, we examined the effect of LPS on PEPCK1 expression in adipose tissue. As shown in Figure 2A, LPS administration results in a very rapid decrease in PEPCK1 mRNA levels in adipose tissue. Two hours after LPS treatment PEPCK1 mRNA levels are decreased by 50%, and by 4 h there is an approximately 90% decrease that is sustained for at least 48 h. The decrease in PEPCK1 mRNA levels is a sensitive response, with a half-maximal effect seen with the administration of approximately 5 µg/kg of LPS, and a maximal effect at 0.5 mg/kg (Figure 2B). Thus, the decrease in PEPCK1 expression in adipose tissue induced by LPS administration is a very sensitive response that occurs rapidly and is sustained for a prolonged period of time.
The effect of LPS administration on phosphoenolpyruvate carboxykinase 1 (PEPCK1) mRNA in adipose tissue. Mice were injected intraperitoneally with the indicated dose of LPS and the animals were euthanized at the indicated times after LPS administration. For mRNA quantification, total RNA was isolated from adipose tissue, cDNA was synthesized with reverse transcriptase, and quantitative real-time PCR performed as described in the Materials and Methods. (A) Mice were treated with saline or 5 mg/kg LPS and euthanized at the indicated time and total RNA prepared from adipose tissue. The controls for each time point serve as the 100% value. (B) Mice were injected with the indicated dose of LPS, euthanized 16 h later and total RNA was prepared from adipose tissue. The data are presented as the mean ± SD. Data are expressed as a percentage of controls. N = 5 per group. *P < 0.05, **P < 0.01, ***P < 0.001.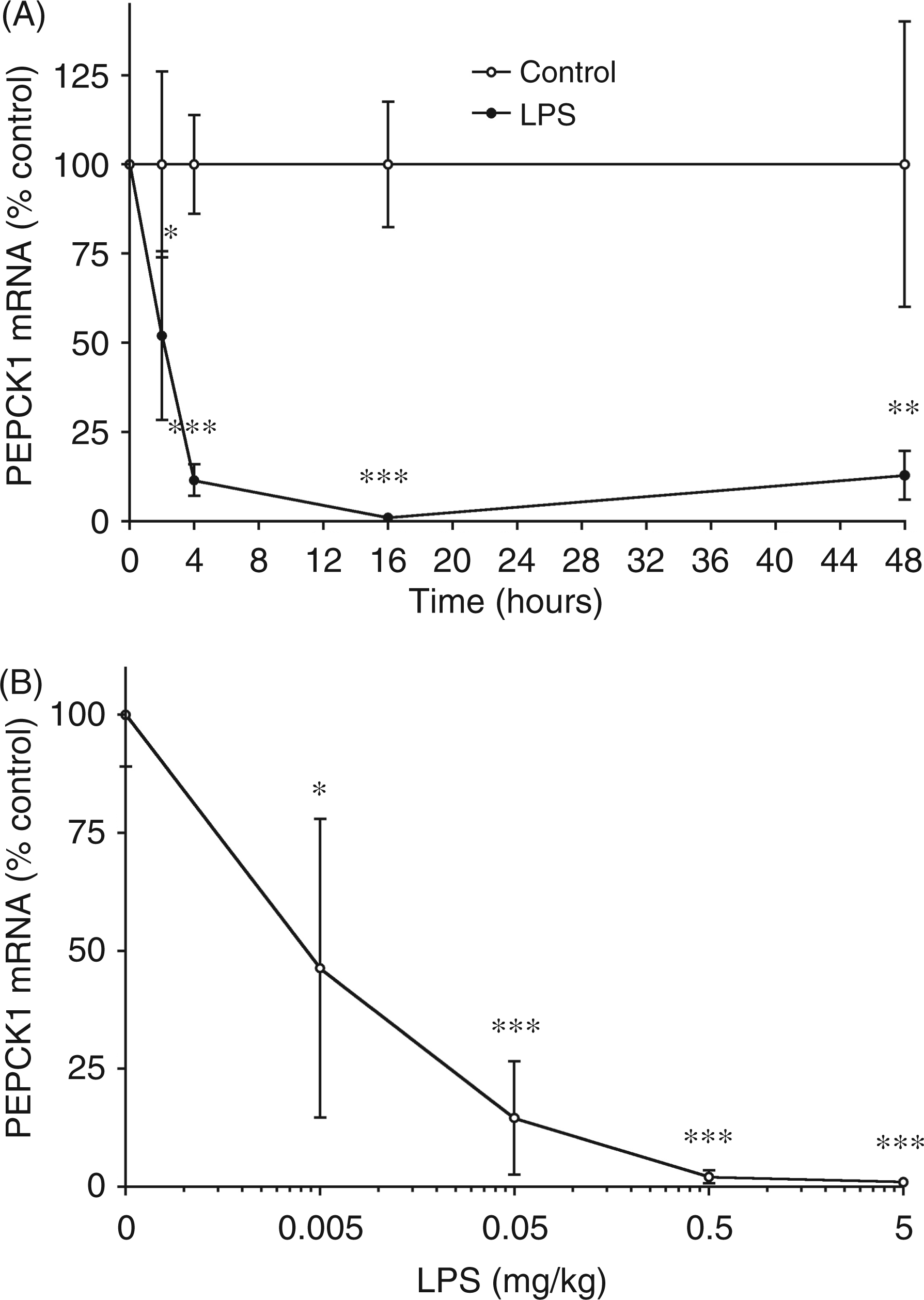
We next studied the effect of LPS administration on PEPCK1 expression in mice with a deletion of the TLR4 receptor. As shown in Figure 3 (A, B), mice with a deletion of the TLR4 receptor do not exhibit a decrease in PEPCK1 mRNA levels in either liver or adipose tissue. These results demonstrate that the activation of the TLR4 receptor plays an important role in mediating the LPS induced decrease in PEPCK1 expression in both liver and adipose tissue.
The effect of LPS administration on phosphoenolpyruvate carboxykinase 1 (PEPCK1) mRNA in liver and adipose tissue from animals deficient in TLR4. Wild-type and TLR4 -/- mice were injected intraperitoneally with LPS (0.5 mg/kg) and the animals were euthanized at 16 h after LPS administration. (A) PEPCK1 mRNA levels in liver. (B) PEPCK1 mRNA levels in adipose tissue. For mRNA quantification, total RNA was isolated from adipose tissue, cDNA was synthesized with reverse transcriptase, and quantitative real-time PCR performed as described in the Materials and Methods. The data are presented as the mean ± SD. Data are expressed as a percentage of controls. N = 5 per group. ***P < 0.001.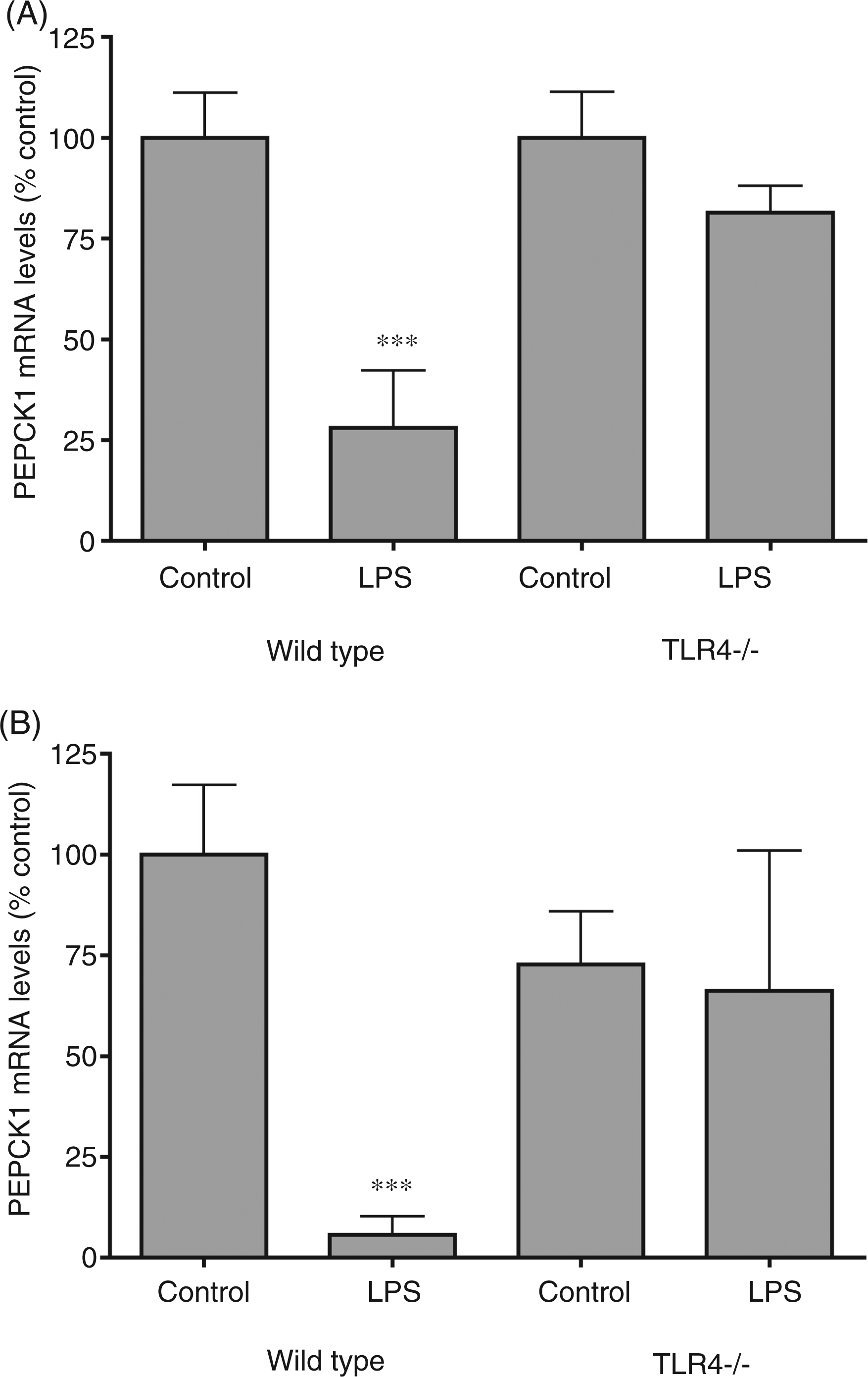
We next studied two other treatments that stimulate inflammation by different pathways. Zymosan stimulates inflammation by activating TLR240,41, whilst turpentine directly increases IL-1 production.42,43 Similar to LPS, zymosan administration induced a marked decrease in PEPCK1 mRNA levels in liver and adipose tissue (Figure 4). Surprisingly, while turpentine induced a marked decrease in PEPCK1 expression in liver, the administration of turpentine had no effect on PEPCK1 expression in adipose tissue (Figure 4).
The effect of zymosan and turpentine administration on phosphoenolpyruvate carboxykinase 1 (PEPCK1) mRNA. Mice were injected with zymosan (80 mg/kg IP) or Oil of Turpentine (100 µl sc in left hind leg) and the animals were euthanized at 16 h after administration. For mRNA quantification, total RNA was isolated from liver and adipose tissue, cDNA was synthesized with reverse transcriptase, and quantitative real-time PCR performed as described in the Materials and Methods. The data are presented as the mean ± SD. Data are expressed as a percentage of controls. N = 5 per group. ***P < 0.001.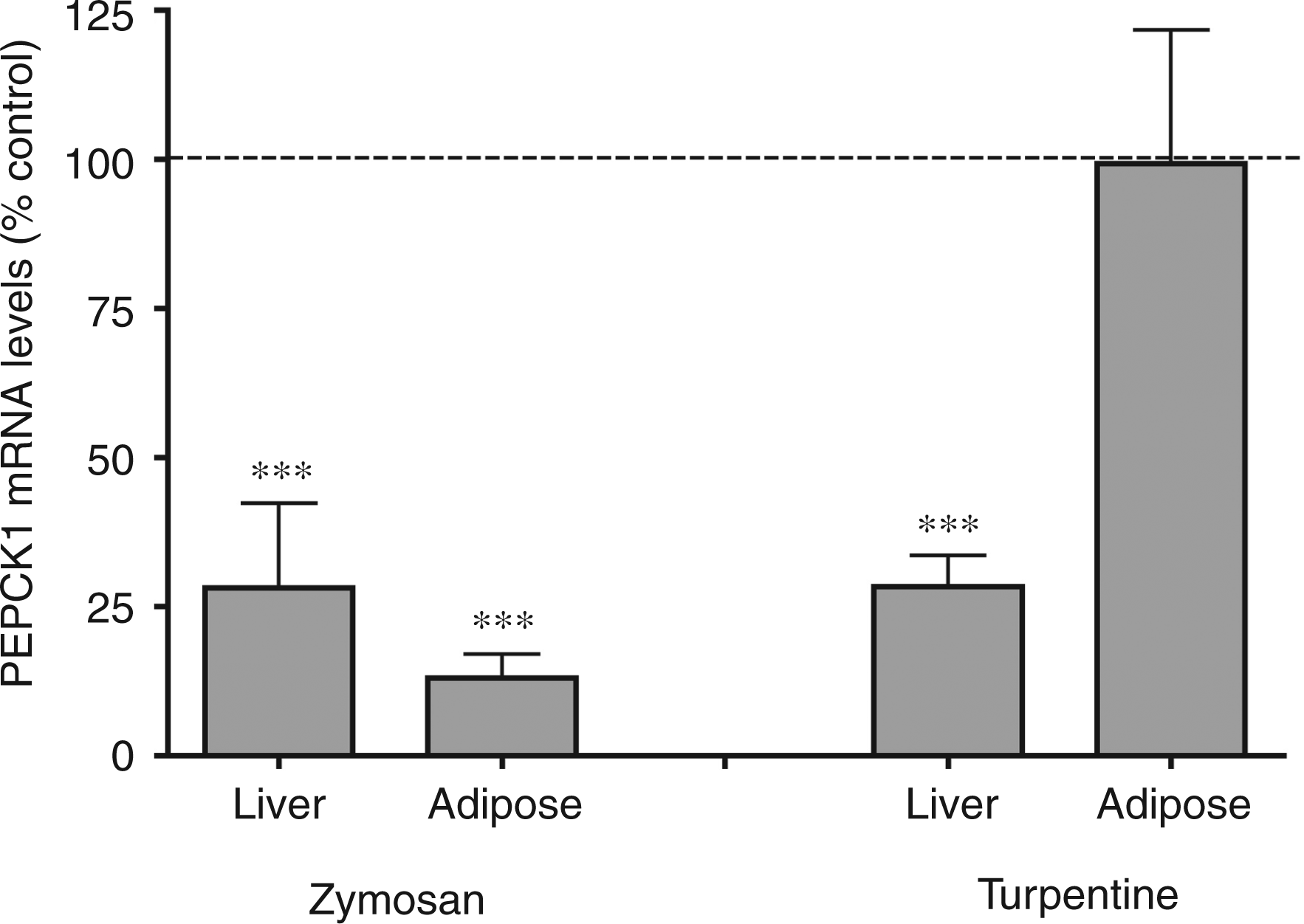
Lipopolysaccharide, zymosan, and turpentine stimulate cytokine production and cytokines are well known to mediate many of the changes in metabolism that occur during inflammation. As shown in Figure 5(A), both TNF-α and IL-1 administration resulted in a decrease in PEPCK1 mRNA levels in the liver. In adipose tissue, TNF-α induced a marked decrease in PEPCK1 mRNA, while IL-1 treatment resulted in only a modest decrease that was not statistically significant (Figure 5B), consistent with the lack of effect of turpentine in adipose tissue. In mice deficient in both the TNF receptor and IL-1 receptor, LPS administration did not decrease PEPCK1 mRNA levels in the liver (Figure 5C) indicating that the LPS-induced inhibition of PEPCK1 expression in the liver is mediated by TNF-α and/or IL-1. In contrast, in adipose tissue, LPS treatment still induced a marked reduction in PEPCK1 mRNA levels (Figure 5C), indicating that neither TNF-α nor IL-1 is required for the LPS induced inhibition of PEPCK1 expression in adipose tissue.
The effect of cytokines on phosphoenolpyruvate carboxykinase 1 (PEPCK1) mRNA in adipose tissue. For mRNA quantification, total RNA was isolated from adipose tissue, cDNA was synthesized with reverse transcriptase, and quantitative real-time PCR performed as described in the Materials and Methods. Mice were injected intraperitoneally with TNF-α (50 µg/kg) or IL-1β (4 µg/kg) and the animals were euthanized at 16 h after cytokine administration. (A) PEPCK1 mRNA levels in liver. (B) PEPCK1 mRNA levels in adipose tissue. (C) Wild type and TNF receptor/IL-1 receptor double knockout (KO) mice were injected intraperitoneally with lipopolysaccharide (LPS) (5 mg/kg) and the animals were euthanized at 4 h after LPS administration. Phosphoenolpyruvate carboxykinase1 mRNA levels in liver and adipose. The data are presented as the mean ± SD. Data are expressed as a percentage of controls. N = 5 per group. *P < 0.05, ***P < 0.001. N.S., not significant.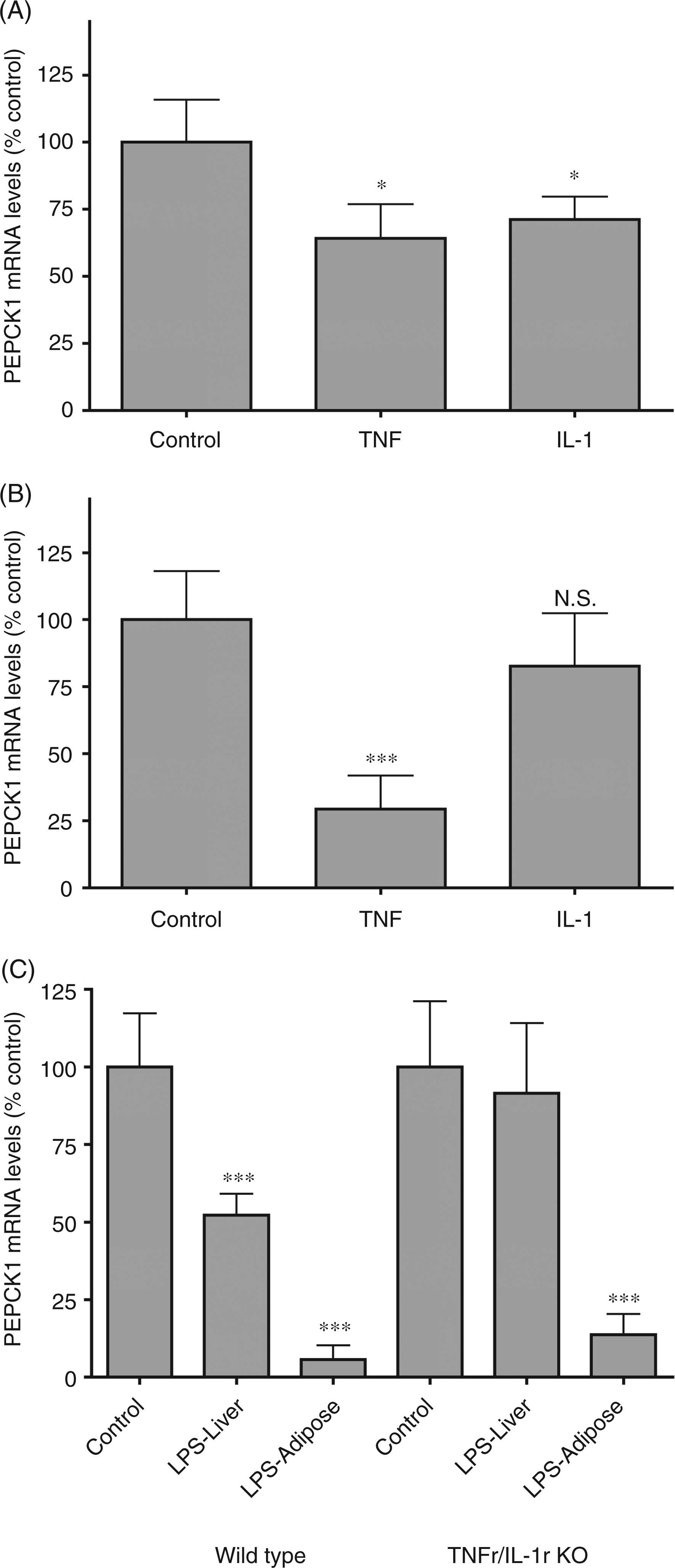
To further define the role of cytokines in decreasing adipose tissue PEPCK1 expression, we next determined the effect of various cytokines on PEPCK1 mRNA levels in 3T3-L1 adipocytes in culture. As shown in Figure 6(A), in addition to TNF-α and IL-1 decreasing PEPCK1 expression, IL-6 and INF-γ also decreased PEPCK1 mRNA levels. In fact, the effect of INF-γ was particularly pronounced. These results suggest that multiple cytokines may mediate the effects of LPS and zymosan on PEPCK1 expression in adipocytes. In contrast, low concentrations of LPS had no effect on PEPCK1 expression, presumably because adipocytes express minimal amounts of the TLR receptors (Figure 6B). However, very high concentrations (1–10 µg/ml) of LPS modestly reduced PEPCK1 mRNA levels.
The effect of LPS and cytokines on phosphoenolpyruvate carboxykinase 1 (PEPCK1) mRNA in adipocytes. Murine 3T3-L1 cells were cultured and induced to differentiate into adipocytes as described in the Materials and Methods. After 10 d of differentiation, the differentiated adipocytes were washed once and treated with the indicated cytokines TNF-α (10 ng/ml), IL-1β (10 ng/ml), IL-6 (10 ng/ml) or INF-γ (100 ng/ml), or LPS (0.1–10 µg/ml) in 0.2% BSA-supplemented medium for 24 h. Total RNA was isolated and PEPCK1 mRNA levels were determined by real-time quantitative PCR analysis as described in the Materials and Methods. (A) Effect of cytokine treatment. (B) Effect of LPS treatment. Data are presented as the mean ± SD. Data are expressed as a percentage of controls. N = 6 per group. qPCR data were normalized to 36B4 mRNA as an invariant control. Data are pooled from two separate experiments for each figure. One-way ANOVA analysis was done to compare all treatments to controls using Tukey-Kramer as the post hoc test. *P < 0.05, **P < 0.01, ***P < 0.001 vs. controls. Ct = 24.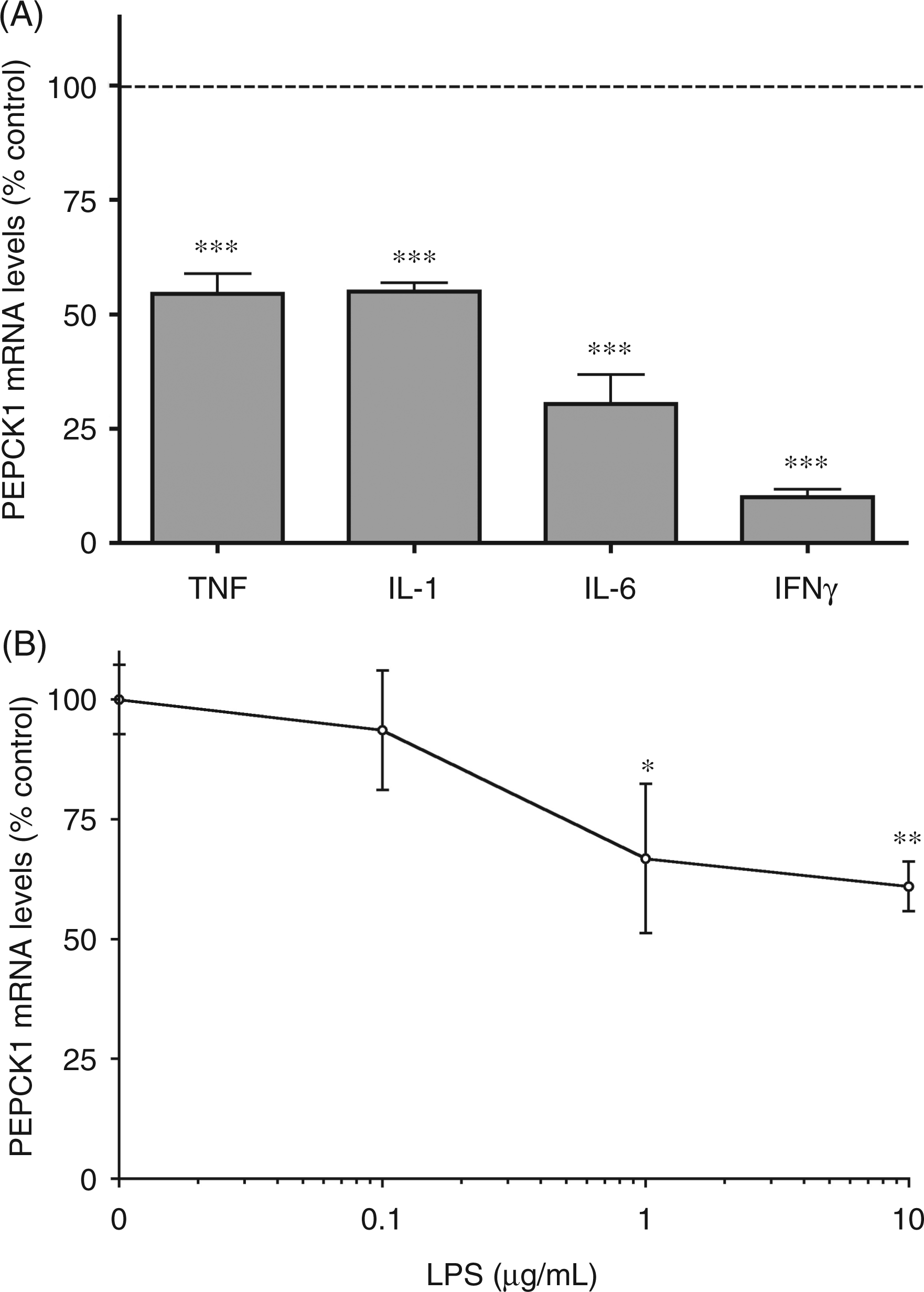
The PEPCK1 promoter contains a peroxisome proliferator activated receptor (PPAR) response element and in adipose tissue PEPCK1 transcription is strongly regulated by PPARγ activation.
33
As seen in previous studies, LPS administration reduces PPARγ (81% decrease) and retinoid X receptor alpha (RXRα) (60% decrease) expression in adipose tissue.21,44 We therefore hypothesized that the decrease in PEPCK1 expression could be due to decreased PPARγ activation. We next used gel shift assays to determine if the binding of PPARγ and RXRα to an oligonucleotide identical to the PPARγ response element in the PEPCK1 promoter is reduced in nuclei of adipose tissue from animals treated with LPS. As shown in Figure 7(A), nuclear extracts from control adipose tissue resulted in a strong shift in the PPARγ response element oligonucleotide. Competition by cold 100-fold molar excess of the wild-type oligonucleotide (lane 3), but not by mutated oligonucleotide (lane 4) demonstrated the specificity of the binding activity in these samples. Moreover, the bound probe was strongly supershifted when the nuclear extracts were incubated with antibodies against either PPARγ (lane 5) or RXRα (lane 6) (Figure 7A). As shown in Figure 7(B), nuclear extracts isolated from adipose tissue of animals treated with LPS had a marked reduction in gel shifting of this oligonucleotide compared to saline treated animals. Together, these results demonstrate that the binding of PPARγ and RXRα to the PPARγ response element in the PEPCK1 promoter is markedly reduced following LPS treatment, strongly suggesting that a decrease in PPARγ activation accounts for the decrease in PEPCK1 expression.
The effect of LPS on PPARγ binding in mouse adipose tissue. Electrophoretic mobility shift assay was performed on mouse adipose nuclear extracts (NE) as described in the Materials and Methods. (A) The assay was carried out using 32P labeled rPCK2 probe incubated with no extract (lane 1), nuclear extract pre-incubated with cold wild type (lane 3) or mutated (lane 4) oligos, and supershifted with antibodies against PPARγ (lane 5) or RXRα (lane 6). Arrows indicate shifted and supershifted bands. (B) Mice were injected intraperitoneally with LPS (5 mg/kg) and the animals were euthanized at 16 h after LPS administration and electrophoretic mobility shift assay was performed. The assay was carried out using 32P labeled rPCK2 probe incubated with no extract (lane 1), control nuclear extract (lanes 2–5) or LPS treated nuclear extract (lanes 6–9).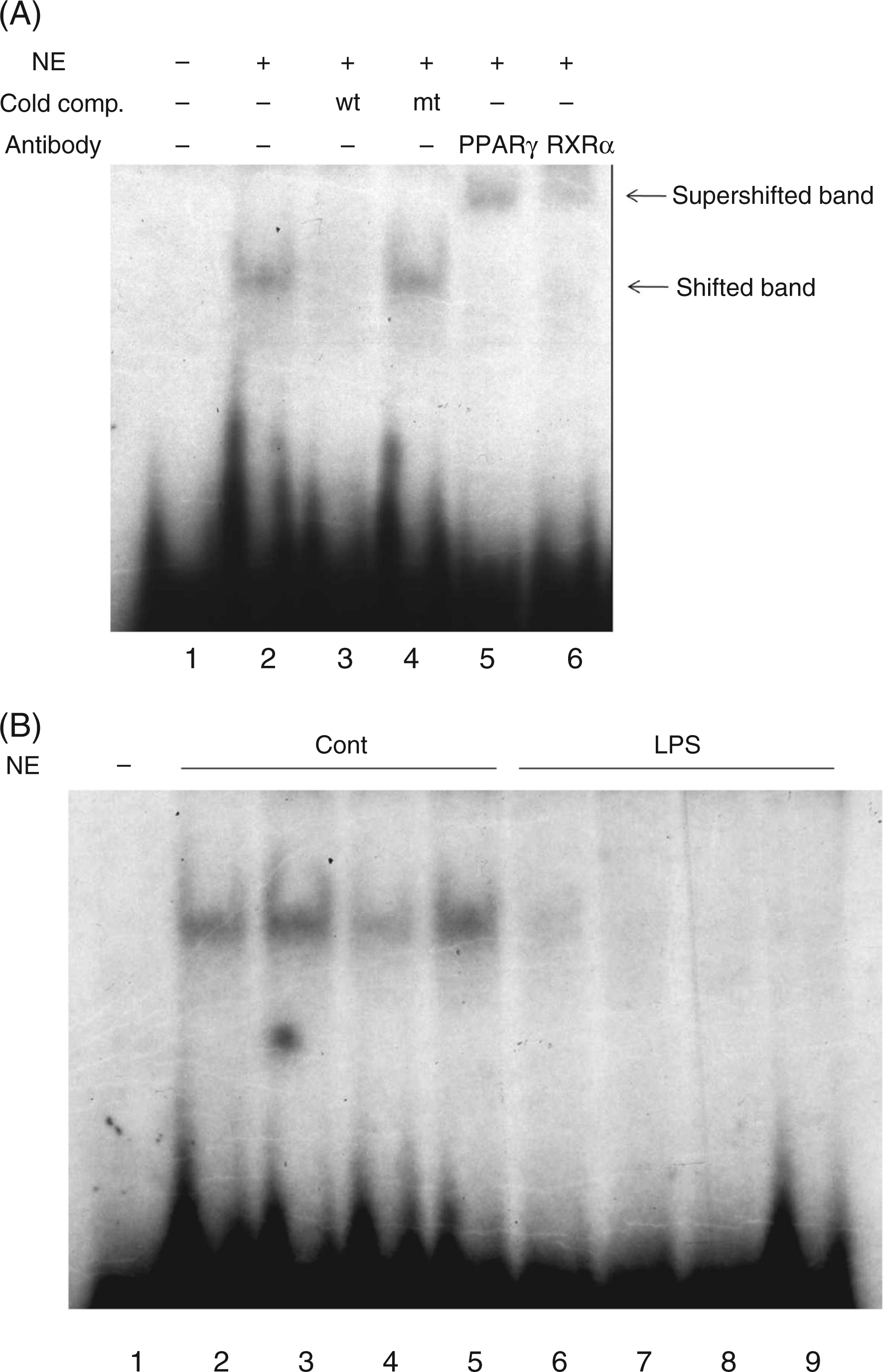
In contrast to the decrease in PPARγ and RXRα induced by LPS or zymosan administration, treatment with turpentine did not alter either PPARγ or RXRα expression in adipose tissue (PPARγ control = 100% ± 15.6, turpentine = 128% ± 13.5; RXRα control = 100% ± 5.8, turpentine = 89% ± 14.6), consistent with the inability of turpentine treatment to down-regulate PEPCK1 mRNA levels. However, it should be noted that turpentine treatment was able to induce a marked increase in Serum amyloid A mRNA levels (89 fold increase) in adipose tissue, indicating that adipose tissue is responsive to turpentine induced inflammation.
Discussion
Previous studies have shown that LPS administration, a model of Gram-negative infection, decreases PEPCK activity and PEPCK1 mRNA levels in the liver. 28 – 32 This decrease has been shown to be caused by the inhibition of PEPCK1 transcription.28,32 The present study confirms and extends these observations and demonstrates that LPS administration decreases PEPCK1 mRNA levels not only in the liver, but in other tissues including the kidney, heart, diaphragm and adipose tissue. Additionally, LPS also decreases PEPCK2 mRNA levels in kidney, diaphragm, heart (not statistically significant) and adipose tissue, but not in liver. As expected, in all tissues except the heart, the expression of PEPCK1 was much more robust than the expression of PEPCK2. The PEPCK activity was decreased in liver, heart, and adipose tissue of LPS treated mice. The effect of LPS on the expression of PEPCK1 is mediated by TLR4 as animals in which TLR4 was deleted did not demonstrate a decrease in PEPCK1 expression in either the liver or adipose tissue following LPS administration. Moreover, zymosan treatment, a model of fungal infections, also induced a decrease in PEPCK1 expression in liver and adipose tissue. In contrast, treatment with turpentine, a model of a sterile abscess, induced a decrease in PEPCK1 expression in the liver, but not in adipose tissue.
In liver, a major function of PEPCK1 is in gluconeogenesis. 24 During severe sepsis, hypoglycemia frequently occurs as a result of a failure in gluconeogenesis, which has been attributed to the decrease in PEPCK1 activity.30,45– 48 In previous studies, we, and others, have shown that the administration of high doses of LPS results in decreases in serum glucose levels.49,50 In adipose tissue, PEPCK1 has a different function, which is to provide glycerol-3-phosphate for the synthesis of triglycerides.23,24 The LPS-induced decrease in PEPCK1 in adipose tissue is a rapid response with a 90% decrease at 4 h following LPS treatment which is sustained for at least 48 h. Additionally, this response is very sensitive with a half-maximal effect seen with the administration of approximately 5 µg/kg of LPS and a maximal effect at 0.5 mg/kg. Lipolysis is stimulated during sepsis leading to elevations in serum free fatty acid levels 7 and it is likely that the decrease in PEPCK activity facilitates the increase in serum free fatty acid levels by decreasing the re-esterification of fatty acids in adipose tissue. Preventing the re-esterification of fatty acids will result in the increased export of fatty acids from the adipocyte, thereby increasing serum free fatty acid levels. It should be recognized that there are multiple mechanisms by which inflammation leads to increases in serum free fatty acid levels. As discussed in detail in the Introduction, cytokines stimulate lipolysis by enhancing the activity of hormone sensitive lipase. 7 Additionally, while the inhibition of PEPCK activity will decrease fatty acid re-esterification, decreases in other enzymes including acyl-CoA synthetase, glycerol-3-phosphate acyltransferase, 1-acyl-glycerol-3-phosphate acyltransferase, lipin 1, monoacylglyceride acyltransferase and diacylglyceride acyltransferase will also contribute to a decrease in fatty acid re-esterification. 20 – 22
The factors that regulate PEPCK1 expression can vary in different tissues.33,34,51 While fasting increases PEPCK1 activity in both liver and adipose tissue, such concordant responses are not always observed.33,34,51 For example, glucocorticoid administration increases hepatic PEPCK1 activity while decreasing PEPCK1 in adipose tissue.33,34,51 It is therefore notable that the effect of LPS and zymosan on PEPCK1 expression in liver and adipose tissue is concordant. In contrast, turpentine administration decreased PEPCK1 mRNA levels in liver but had no effect in adipose tissue. Lipopolysaccharide and zymosan stimulate the production of multiple cytokines, including TNF-α, IL-1, IL-6 and interferons, which mediate many of the metabolic effects observed with LPS and zymosan administration.1,2,41 In contrast, the systemic effects of turpentine treatment are mediated primarily by increases in IL-142,43. The administration of TNF-α decreases the expression of PEPCK1 in liver and adipose tissue, while the administration of IL-1 significantly decreased PEPCK1 expression in the liver, but had more modest effects on adipose tissue that did not reach statistical significance.
Previous studies have shown an important role for TNF-α in the LPS-induced decrease in hepatic PEPCK1 expression.29,48 Specifically, antibodies that neutralize TNF-α activity inhibited the ability of LPS administration to suppress hepatic PEPCK1 expression. 29 In the present study, we demonstrate that the LPS-induced decrease in PEPCK expression in liver does not occur in TNF receptor/IL-1 receptor knockout mice, indicating that TNF-α and/or IL-1 signaling is essential for inhibition of PEPCK1 expression. However, the LPS induced decrease in PEPCK1 expression in adipose tissue still occurs in TNF receptor/IL-1 receptor knockout mice, indicating that neither TNF-α nor IL-1 signaling is required for LPS- induced decrease in adipose tissue. Our studies have further shown that both IL-6 and INF-γ inhibit PEPCK1 expression in 3T3-L1 adipocytes, indicating that other cytokines are also capable of inhibiting PEPCK1 expression. Additionally, LPS can directly affect adipocytes and decrease PEPCK1 expression. However, only very high concentrations of LPS (1–10 µg/ml) affected PEPCK1 expression, presumably because of the low level of TLR4 expression in adipocytes. 52 Thus, in contrast to the liver, it is likely that there are several pathways, other than TNF-α and IL-1, by which LPS administration results in a decrease in PEPCK1 expression in adipose tissue. Lipopolysaccharide administration stimulates the production of a large number of different cytokines and biologically active compounds, which could alter adipose tissue metabolism. In contrast, the systemic effects of turpentine treatment are mediated primarily by increases in IL-1.42,43 It is likely that turpentine treatment does not produce the cytokines/biologically active compounds required to inhibit PEPCK expression in adipose tissue.
The PEPCK1 promoter contains a PPAR response element and in adipose tissue PEPCK1 transcription is regulated by PPARγ activation.33,34,51 Thiazolidinediones, activators of PPAR gamma, stimulate PEPCK1 expression. 51 In this study, using gel shift assays, we demonstrated that the binding of PPARγ and RXRα to the PPARγ response element in the PEPCK1 promoter is markedly decreased in adipose tissue nuclear extracts obtained from LPS- treated animals. In the present study, and in prior studies, we, and others, have shown that both LPS and zymosan administration markedly decrease PPARγ and RXRα expression in adipose tissue.21,44 Additionally, we have shown that TNF-α treatment decreases PPARγ and RXRα in 3T3-L1 adipocytes. 21 In contrast, turpentine treatment does not alter PPARγ or RXRα expression in adipose tissue. These results suggest that a decrease in PPARγ and RXRα accounts for the decrease in PEPCK1 expression in adipose tissue that occurs during inflammation.
In conclusion, the present study demonstrates that inflammation results in a decrease in PEPCK1 and 2 expression in several different tissues. The decrease seen in adipose tissue is mediated by different cytokine pathways than the decrease that occurs in the liver. The decrease in PEPCK1 in adipose tissue is a rapid, sensitive response that would increase the availability of fatty acids for export into the circulation thereby increasing serum free fatty acid levels. An increase in serum free fatty acids would provide the liver with substrate for the synthesis of triglycerides and stimulate VLDL secretion. Additionally, increased serum free fatty acids could provide an energy source for cells that play crucial roles during inflammation.
Footnotes
Acknowledgements
This work was supported by grants from the Research Service of the Department of Veterans Affairs and by the Albert L. and Janet A. Shultz Supporting Foundation.