
Abstract
This study examined whether task-related and environmental constraints influence the vertical ground reaction force (vGRF) waveform during the basketball jump shot, with a focus on phase-specific adaptations across shooting positions. Eighteen senior basketball players performed jump shots from three shooting positions (45° right, middle, 45° left) under three conditions: baseline, simulated defensive opposition, and simulated gym audience noise. vGRF signals were recorded using a force platform and time-normalized to 0–100% of the movement cycle. One-dimensional Statistical Parametric Mapping (SPM) was applied using a hierarchical region-of-interest (ROI) approach, focusing on the unweighting, braking, and propulsive phases. Group-level effects were tested using a two-way SPM{F} ANOVA (Constraint × Position). Exploratory intra-individual SPM{t} analyses were conducted to examine athlete-specific adaptations. No significant main effects of constraint or position, and no constraint × position interactions were observed at the group level in any ROI (α = 0.05). Exploratory intra-individual analyses revealed heterogeneous, athlete-specific adaptations to external constraints and shooting position, predominantly during the braking and propulsive phases, with substantial variability in the timing and presence of significant clusters across athletes.
The results indicate that experienced basketball players preserve a stable force–time organization during the jump shot despite environmental and task-related constraints. While group-level mechanical patterns remain invariant, individual athletes may exhibit phase-specific adaptations that do not generalize across the population. These findings underscore the importance of hierarchical and individualized analytical approaches for understanding skilled performance and support the use of ROI-based SPM for the biomechanical analysis of complex movement tasks.
Introduction
Shooting performance is central to success in basketball, yet in competition it unfolds under task and environmental constraints such as audience noise and defensive pressure. Contested shooting consistently alters how players organize the movement and generally reduces efficiency, motivating analyses that look beyond single discrete outcomes (e.g., accuracy, peak force) to the time structure of the action.1,2
Previous research has shown that several task constraints can influence how basketball players organize their shooting movements. For example, changes in shooting distance have been associated with adjustments in joint coordination, release parameters, and jump mechanics across players of different expertise levels and ages.3,4 Similarly, the presence of defensive opposition can alter spatial and temporal characteristics of the shooting action, requiring players to adapt their technique to maintain shooting effectiveness. 5 These findings highlight that basketball shooting performance is sensitive to task constraints and that intra-individual adaptations in movement organization are a common feature of skilled shooting behavior.
More recent applied investigations specifically examining opponent characteristics further demonstrate that varying the opponent's height and distance leads to observable technique adjustments across the shot, 5 supporting the idea that constraints may relocate or amplify demands within specific phases of the jump (e.g., braking vs. propulsion).
Environmental factors also matter. Laboratory and field reports indicate that spectator presence/noise can modulate performance through arousal and attentional mechanisms, with effects documented in indoor sports settings and in basketball contexts.6,7 While findings vary with expertise and context, this body of work frames noise as a plausible constraint that could shift when differences along the waveform emerge—even if global magnitudes (e.g., jump height) are unchanged.
Force-platform analysis offers a rigorous, time-resolved description of lower-limb mechanics during jumping tasks. In vertical jumps, the vertical ground reaction force (vGRF) curve is commonly segmented into unweighting, braking (eccentric), and propulsive (concentric) phases, followed by flight—each associated with distinct neuromuscular functions.8,9 Moreover, objective signal-based events (e.g., intersections with the body weight (BW) line, vGRF maximum, threshold-based take-off) enable reproducible phase partitioning suited to sport-specific adaptations—in our case, the basketball jump shot, where ball release typically occurs shortly after take-off.8–13
From an inference perspective, analyzing entire time-normalized curves using Statistical Parametric Mapping (SPM) allows us to localize where in time conditions differ, while controlling the family-wise error rate across the continuum.14,15 SPM has proven sensitive to condition effects in countermovement-jump research—detecting phase-specific clusters that classical discrete metrics can miss—and is increasingly used in sports biomechanics for intra-individual designs, reliability questions, and intervention/constraint comparisons.16–18
From a coaching perspective, phase-level, time-localized diagnostics offer clear levers for practice design. If audience noise yields clusters earlier in the waveform (e.g., during unweighting), sessions can integrate sound-pressure levels similar to competition alongside attention-regulation routines (e.g., pre-shot breathing and quiet-eye cues) that have been shown to stabilize execution under pressure.19–21 If SPM shows that defensive opposition shifts differences toward the braking–propulsive complex, coaches can emphasize training that focuses on the corresponding phase of the movement, in line with a constraints-led approach that manipulates task and environmental constraints while preserving representativeness.22–24
Therefore, this study aimed to test, within athletes, whether court position (left 45°, center, right 45°) and constraint conditions (baseline, gym audience noise and simulated defensive opposition) produce time-localized differences in the vGRF waveform during the basketball jump shot, using SPM. To aid interpretation, significant clusters are mapped to functional phases (unweighting, braking, propulsive).8,9,11–13 Based on prior work, we hypothesized that defensive opposition would yield clusters mainly around the braking–propulsive complex (e.g., faster transition, altered peak region), whereas gym-audience noise would produce subtler or athlete-dependent clusters whose timing may differ across phases.1,5,6
Methods
Participants
Eighteen senior basketball players (8 male and 10 female; age: 22.00 ± 3.72 years; height: 172.15 ± 9.79 cm; body mass: 69.6 ± 13.12 kg) volunteered to participate. All were actively competing at a senior national level and had a minimum of seven years of playing experience. At the time of testing, players were regularly engaged in structured team training and official competition during the competitive season. Inclusion criteria required participants to be active senior basketball players with regular training participation and competitive experience. Exclusion criteria included the presence of musculoskeletal injury, pain, or any medical condition that could affect shooting performance in the three months prior to testing. Participants were free from injuries in the three months prior to testing. Written informed consent was obtained from all players, and the study was approved by the institutional ethics committee in accordance with the Declaration of Helsinki (approval number CE/FCDEF-UC/00812021). All data were collected and stored anonymously to ensure participant confidentiality.
Experimental protocol
Immediately before testing, body mass was measured using a calibrated scale (to the nearest 0.1 kg) and later used to normalize vGRF to body weight (BW = mass × g, where g = 9.81 m/s2).25,26 Expressing vGRF relative to BW reduces inter-individual variability.23,24 After that, the player performed a standardized 10-min warm-up that combined functional activation drills with basketball shooting practice. Following the warm-up, players positioned themselves on a pair of force platforms, with one foot on each platform, and were instructed to initiate every attempt from this stance. At the beginning of each trial, the ball was passed by chest pass directly to the participant, who started the attempt grabbing the ball while standing on the force platforms. Participants were instructed to initiate the shot from this position, right after receiving the ball (“catch and shot”), without dribbling to ensure consistent initial conditions across trials and reliable force platform measurements. No restrictions were imposed on the subsequent movement or shot execution to preserve the athletes’ natural shooting technique.
From a fixed distance of 6.75 m, participants performed ten shots from each of three shooting positions (right 45° (45d), central (m), left 45° (45e)) under each constraint condition. The three constraint conditions were baseline (B), simulated gym audience noise (R), and simulated defensive opposition (O). Therefore, each participant completed a total of 90 shots (10 shots × 3 positions × 3 conditions). The sequence of constraint conditions was randomized for each participant using a computer-generated random order to minimize potential order effects such as learning across the experimental trials. Between consecutive shots in the same condition, a 30-s recovery was provided, whereas a 2-min pause separated transitions between shooting positions.
The constraints were designed to reproduce environmental and task-related challenges commonly encountered during games. Environmental noise consisted of pre-recorded gymnasium crowd sounds delivered via external speakers at ∼105 dBA, consistent with levels reported in indoor sports facilities.6,7 Audio began before the first shot of the noise condition and was maintained across repetitions. Opponent pressure was reproduced using adjustable blocking equipment set at 1.20*athlete's height and positioned 1 m in front of the shooter. 27
Data collection was performed indoors under standardized conditions, with athletes using regulation basketballs (size and mass adjusted to sex, pressure set at 0.62 bar), their usual basketball shoes, and training apparel.
Data acquisition
vGRF were collected using two force platforms (Bertec model 4060-05, Bertec Corporation, Columbus, OH, USA) at a sampling frequency of 1000 Hz. The platforms were placed side by side to allow players to stand with both feet within the boundaries of the two plates, but they could self-selected how their feet were placed within these boundaries. The analysis used the sum of the vertical forces from both plates (vGRF = vGRF_left + vGRF_right). Signals were collected with the QTM software and exported in raw format for offline processing in MATLAB (R2023a, MathWorks, USA).
Data processing
Raw force-time signals were visually screened to exclude trials with artifacts or technical errors. Each curve was cropped between the third-to-last crossing of the line representing 1×BW line (BW line) and take-off, Figure 1. Take-off was defined as the instant vGRF fell < 5% BW for ≥3 consecutive frames, a common criterion in jump analyses. 10 To enable time-domain comparisons, curves were resampled (cubic interpolation) to 101 points representing 0–100% of the movement cycle after cropping. This 1D, time-normalized approach is standard in SPM workflows for biomechanical trajectories.14,15
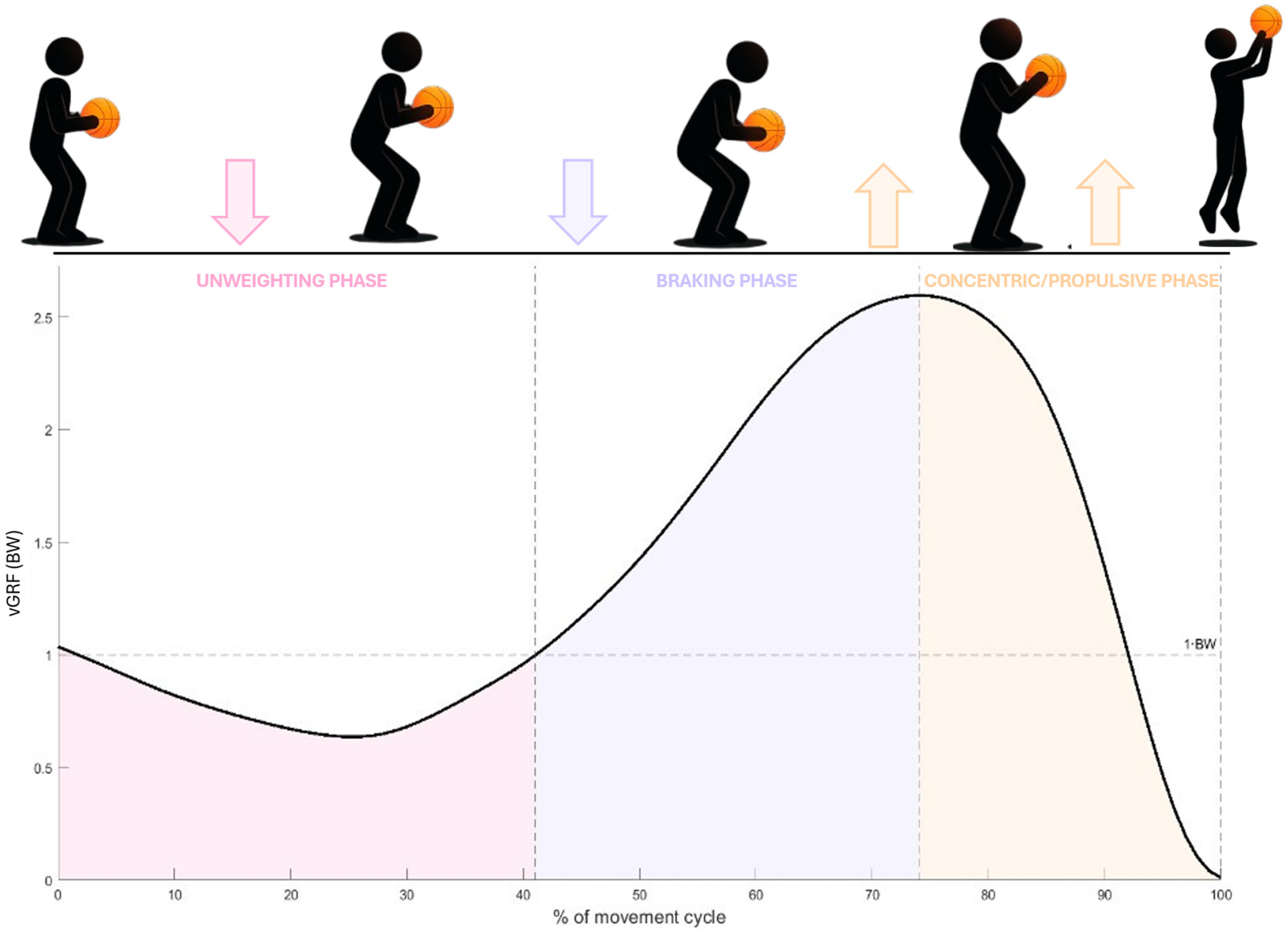
Phases of the basketball jump shot: Unweighting (onset to the second intersection with 1×BW), Braking (second intersection to peak force), and Propulsive/Concentric (peak force to take-off). The curve show Fz normalized to body weight, with the 1×BW reference line displayed for context.
Phase definition
Following operational definitions tailored to the basketball jump shot, three functional phases of the countermovement were identified from the vGRF curve (Figure 1):
- Unweighting: from the antepenultimate crossing of vGRF below the BW line to the second intersection with BW line, corresponding to the end of net unloading and the initiation of downward motion. - Braking: from the second BW line intersection to peak vGRF, representing the upward net impulse applied to decelerate the negative centre of mass (COM) velocity. - Propulsion (concentric): from peak vGRF to take-off, defined as the point when Fz dropped below 5% BW, reflecting the concentric impulse responsible for generating upward COM velocity.
These boundaries are consistent with established descriptions of countermovement jump phases in the biomechanics literature. These phase boundaries are consistent with established descriptions of the countermovement jump force–time curve in the biomechanics literature, where unweighting, braking, and propulsion represent key functional phases of the movement.8,9 Together, these operational definitions provide both ecological validity for basketball-specific jump shots and methodological consistency across studies of vertical jump mechanics.
Statistics analysis
All inferential analyses of the vGRF waveform were conducted using one-dimensional SPM (SPM{F} and SPM{t}), implemented in MATLAB with the open-source spm1d toolbox.14,15 Force–time signals were time-normalized to 0–100% of the movement cycle, and statistical inference across the one-dimensional continuum was based on Random Field Theory (RFT), which controls the family-wise error rate while accounting for temporal correlation in biomechanical waveforms. All SPM{t} contrasts were evaluated using two-tailed inference (α = 0.05), with RFT-based cluster-level inference. Accordingly, both positive and negative supra-threshold clusters were considered significant.
The primary inferential analysis was conducted at the group level. For visualization purposes only, descriptive vGRF curves were first plotted for each experimental condition and shooting position, displaying all individual trials and the across-athlete mean waveform to illustrate the consistency of the unweighting–braking–propulsive pattern. These descriptive plots were not used for statistical inference.
To directly address the study hypotheses while improving functional interpretability, group-level inference was based on phase-specific region-of-interest (ROI) analyses. Three ROIs corresponding to functional phases of the jump shot were defined a priori on the normalized force–time waveform: unweighting, braking, and propulsion. These ROIs were selected based on established biomechanical descriptions of the countermovement jump and basketball shooting mechanics.
Within each ROI, a two-way SPM{F} analysis of variance (ANOVA) was performed with the factors Constraint (B, R and O) and Position (45d, m and 45e). To ensure a balanced design across the 3 × 3 experimental matrix, the number of trials per athlete and condition was equalized by subsampling to the minimum number of available trials within each cell. Statistical significance was assessed at α = 0.05 using RFT-based cluster-level inference.
Post-hoc pairwise SPM{t} comparisons were planned only in the presence of a significant main effect or interaction in the omnibus ROI-based ANOVA.
To complement the group-level findings and explicitly examine inter-individual variability, additional intra-individual analyses were conducted. For each athlete, trials were analyzed separately using SPM{t}, allowing condition effects to be evaluated within each individual rather than across pooled data. This approach enabled the identification of athlete-specific supra-threshold clusters and the comparison of their presence, timing, and duration across participants, thereby characterizing the heterogeneity of individual mechanical adaptations. Within each athlete, constraint effects (B vs R vs O) were examined within each shooting position, and positional effects (45d vs m vs 45e) were examined within each constraint condition. These analyses followed the same ROI definitions and RFT-based inference procedures described above.
To control for multiple planned contrasts within each family of comparisons, the Holm–Bonferroni step-down correction was applied. 28 For each athlete, significant clusters were characterized by their onset, offset, duration, and corresponding movement phase (unweighting, braking, or propulsion). These intra-individual analyses were considered exploratory and were used to describe the heterogeneity of individual mechanical adaptations rather than to support population-level inference.
Results
To provide an overview of the mechanical patterns across experimental conditions, vGRF profiles were first examined (Figure 2). Across all athletes and conditions, the force–time curves consistently exhibited the expected unweighting, braking, and propulsive phases of the basketball jump shot. The overall waveform shape and temporal organization were comparable across shooting positions and constraint conditions, with no visually apparent systematic shifts in force magnitude or timing. These descriptive observations confirm consistent task execution and support the suitability of subsequent phase-specific statistical analyses.

Ensemble vGRF curves across the nine experimental conditions. Each colored line represents the mean curve of one athlete; the grand mean across athletes is shown in black. The x-axis is normalized to the movement cycle (0–100%). Vertical dashed lines indicate the approximate transitions between unweighting, braking, and propulsive phases. Legend: B45d – Right 45°, Baseline condition; Bm – Middle 90°, Baseline condition; B45e – Left 45°, Baseline condition; R45d – Right 45°, Simulated gym audience noise condition; Rm – Middle 90°, Simulated gym audience noise condition; R45e – Left 45°, Simulated gym audience condition; O45d – Right 45°, Opposition condition; Om – Middle 90°, Opposition condition; O45e – Left 45°, Opposition condition.
Across all ROIs, the two-way SPM{F} ANOVA revealed no significant main effects of Constraint (B, R and O), no significant main effects of Position (45d, m and 45e), and no significant Constraint × Position interactions (α = 0.05). In all cases, the SPM{F} trajectories remained below the critical thresholds throughout the analyzed regions, and no supra-threshold clusters were identified (Figure 3).
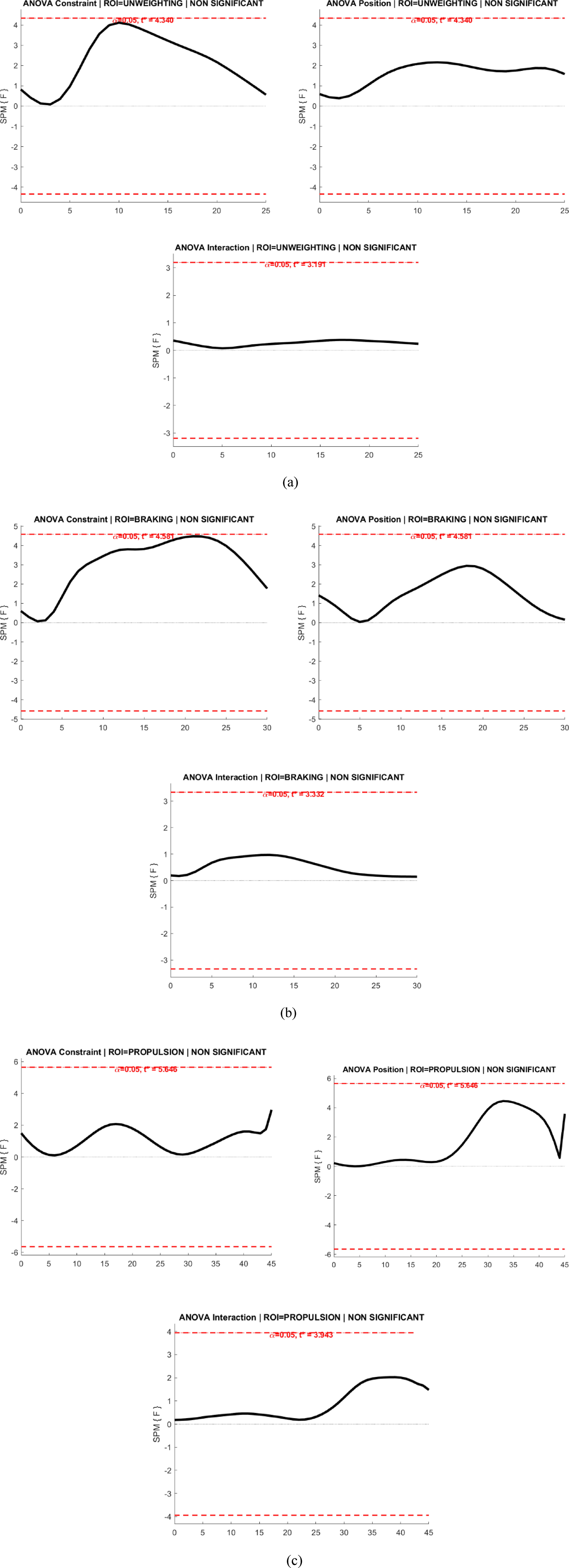
Results of the two-way SPM{F} ANOVA with the factors Constraint (B, R, O) and Position (45d, 45e, m). Panels display the statistical trajectories for (a) the main effect of constraint, (b) the main effect of position, and (c) the constraint × position interaction. The red dashed lines represent the critical threshold.
Given the absence of significant omnibus effects in any ROI, no post-hoc pairwise SPM{t} comparisons were performed.
Although no systematic group-level effects were detected, several athletes exhibited significant supra-threshold clusters within specific ROIs. These effects occurred predominantly during the braking and propulsive phases, whereas fewer clusters were observed during the unweighting phase. The presence, temporal location, and duration of significant clusters varied substantially across athletes and contrasts, with no consistent temporal pattern emerging at the population level.
To further characterize the functional relevance of these findings, each significant supra-threshold cluster identified in the intra-individual analyses was classified according to its temporal overlap with the predefined movement phases. The distribution shown in Figure 4 represents the total number of significant clusters across all athletes and contrasts, with each cluster counted once and assigned to a single phase. Percentages were calculated relative to the total number of detected clusters, thereby describing the relative frequency of clusters occurring in each movement phase rather than the number of athletes.
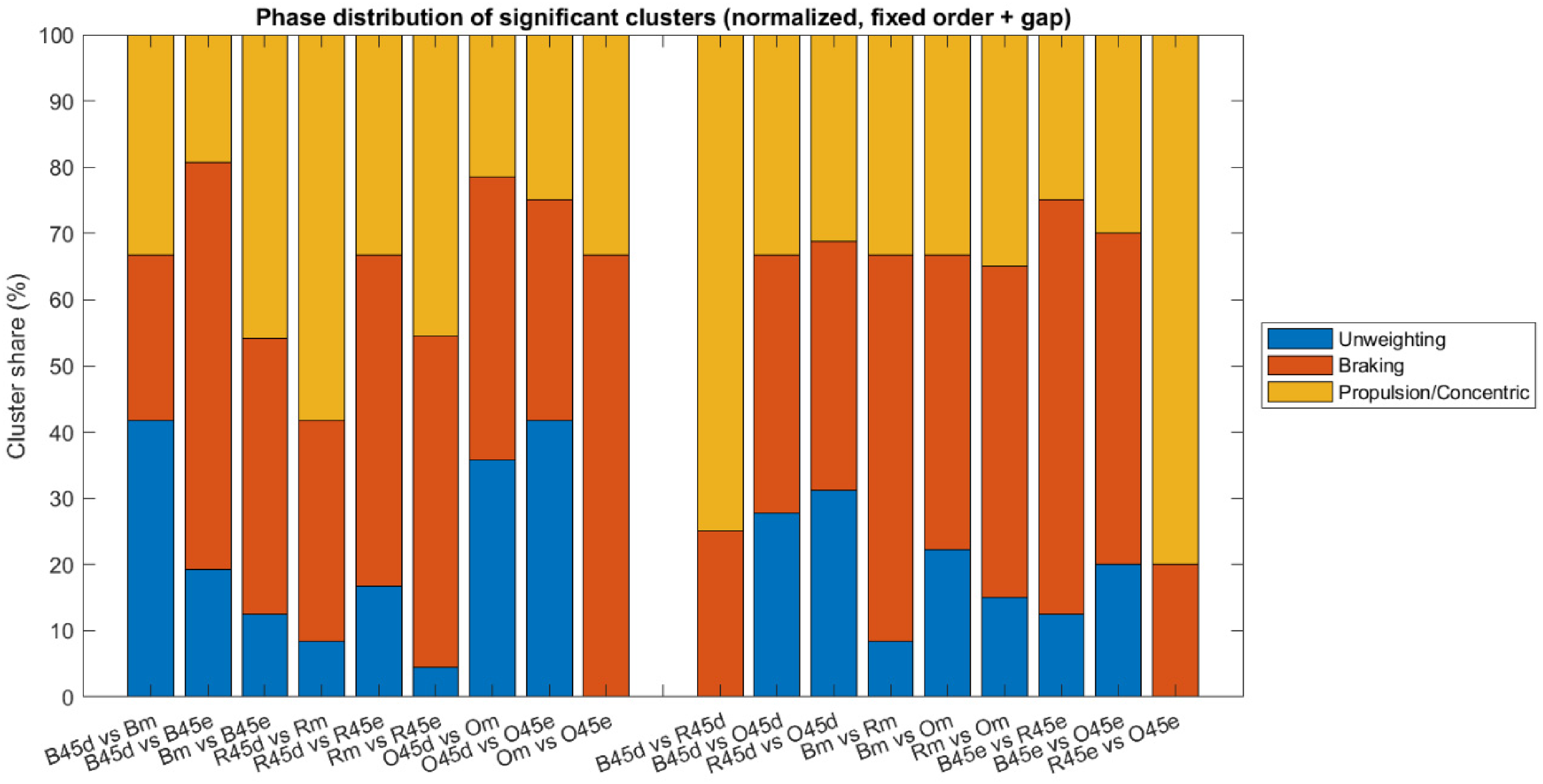
Distribution of significant intra-individual clusters across movement phases. Bars represent the cumulative number of clusters classified within the unweighting, braking, and propulsive phases based on their temporal overlap with phase-specific ROIs.
Taken together, these exploratory analyses indicate heterogeneous, athlete-specific mechanical adaptations that were localized in time but not sufficiently consistent across participants to produce statistically significant effects at the group level.
Overall, intra-individual clusters were most frequently located in the braking and concentric phases, with unweighting less affected. Significant intra-individual effects were most frequently detected in braking (81 clusters) and concentric/propulsive (60 clusters) phases, with durations typically ranging between 7–22% of the normalized cycle.
Discussion
The present findings suggest that experienced basketball players maintain a stable force–time organization during the jump shot despite the presence of environmental and task-related constraints. The absence of consistent group-level effects across phases indicates that the global mechanical structure of the movement is robust to moderate contextual perturbations. Nevertheless, the intra-individual analyses revealed heterogeneous, athlete-specific adaptations, particularly during the braking and propulsive phases, suggesting that performers may rely on individualized coordination strategies rather than a uniform group response.
The absence of significant group-level differences contrasts with previous studies reporting altered lower-limb kinetics or reduced shooting performance under defensive pressure or noisy environments.19,29,30 However, many of these studies relied on discrete outcome variables such as jump height, peak force, or shooting accuracy, rather than continuous waveform-based analyses. Moreover, effects of opposition and noise have been shown to be more pronounced in less experienced performers than those recruited to our study or under high-pressure competitive scenarios. 31
Another factor that may contribute to variability in force–time characteristics is sex-related differences in physical and neuromuscular capacities. Previous research has shown that male and female athletes may exhibit different strength levels, countermovement depths, and force production strategies during jumping tasks.8,32 Although vertical ground reaction forces in the present study were normalized to body weight to reduce inter-individual variability, potential sex-related differences in movement strategies cannot be entirely excluded. Future research with larger samples may therefore explore whether sex-specific adaptations exist in the force–time organization of the basketball jump shot under different task constraints.
In the present study, participants were senior-level athletes with extensive competitive experience, which may explain the stability of the vGRF waveform across conditions. From a motor control perspective, skilled performers are known to preserve task-relevant mechanical variables despite contextual perturbations, reflecting functional invariance at the level of global coordination patterns. 33 Thus, the absence of group-level effects does not indicate a lack of adaptation but rather suggests that force–time organization is a highly stable feature of expert jump-shot execution.
Although no systematic group-level effects were detected, exploratory intra-individual analyses revealed several time-localized supra-threshold clusters within individual athletes, predominantly during the braking and propulsive phases of the vGRF waveform. These clusters reflected localized differences in the magnitude and timing of force production across constraint conditions within individual athletes. Biomechanically, these phases are critical, as they regulate eccentric force absorption, the eccentric–concentric transition, and net impulse generation, which directly influence take-off velocity and release conditions.8,34
From a motor control perspective, the coexistence of stable group-level force–time organization and heterogeneous intra-individual adaptations may reflect the concept of motor abundance, whereby skilled performers exploit multiple coordination strategies to achieve similar task outcomes. Rather than relying on a single optimal movement solution, experienced basketball players may adjust the timing and magnitude of force production within specific phases of the jump shot while preserving the overall mechanical structure of the movement.
The heterogeneous distribution of these effects across athletes aligns with previous work highlighting substantial inter-individual variability in lower-limb force strategies during jumping and shooting tasks.34,35 Rather than converging toward a single optimal solution, skilled athletes appear to exploit multiple coordination strategies to achieve equivalent task outcomes, a phenomenon commonly described within the framework of motor abundance. 36
Importantly, the lack of temporal consistency within athletes could explain why these adaptations did not emerge at the group level. Individual differences in strength profiles, perceptual strategies, and atonement to environmental constraints may lead to localized adjustments in braking or propulsion that are averaged out when pooled across participants.
Simulated defensive opposition and gym audience noise were expected to differentially influence force production due to their distinct informational and psychological demands. Defensive opposition directly constrains available time and space, potentially requiring rapid modulation of force during braking and propulsion, whereas audience noise is thought to influence performance through attentional and arousal-related mechanisms.30,37
The present findings suggest that, at the waveform level, neither constraint elicited consistent group-level changes in vGRF. Nevertheless, the exploratory intra-individual results indicate that certain athletes responded to these constraints with phase-specific adjustments, particularly during braking and propulsion. These findings are consistent with ecological dynamics perspectives, which posit that constraints shape behavior by narrowing the landscape of available solutions rather than enforcing uniform responses across individuals.38,39
A key methodological contribution of this study lies in the use of phase-specific ROI-based SPM within a hierarchical inferential framework. Previous SPM studies in sports biomechanics have typically analyzed the full normalized waveform without explicit segmentation into biomechanically meaningful phases. 15 While such approaches are statistically valid, they may yield temporally localized clusters that are difficult to interpret in functional or mechanical terms. In contrast, organizing inference according to predefined movement phases provides a structured biomechanical context for statistical results. Although the entire propulsion cycle was still analyzed, partitioning the waveform into functional segments improves interpretability and facilitates clearer links between detected clusters and the underlying mechanics of the movement.
The lack of significant group-level effects across all ROIs further emphasizes the importance of cautious interpretation of pairwise waveform comparisons in the absence of omnibus significance. At the same time, exploratory intra-individual SPM analyses provide valuable insight into athlete-specific adaptations that would otherwise remain hidden in group-averaged analyses.
These findings may also have practical implications for performance analysis and coaching. Rather than expecting uniform mechanical responses to environmental or task constraints, coaches and practitioners should consider that skilled players may adapt their force production strategies in individualized ways. Monitoring phase-specific force–time characteristics may therefore provide useful information about how athletes regulate movement coordination when performing under different contextual demands.
This study has some limitations that should be acknowledged. First, although the sample included experienced senior basketball players, the sample size was relatively modest for examining subgroup-specific effects, and both male and female athletes were analyzed together without a sex-specific comparison. Second, the experimental constraints were simulated under controlled conditions and may not fully reproduce the perceptual, emotional, and temporal demands of real competition. In addition, each participant completed a relatively large number of shooting attempts (90 trials), which may have induced some degree of fatigue toward the later stages of the protocol. Although recovery periods were provided between trials and positions, and the order of conditions was randomized, fatigue effects cannot be completely excluded. Therefore, caution is warranted when interpreting the results, as subtle fatigue-related changes in force–time characteristics may have occurred during the session. Third, only the vertical ground reaction force waveform was analyzed, which provides important information about lower-limb force-time organization but does not capture the full complexity of shooting coordination or performance outcomes. Finally, the intra-individual analyses were exploratory in nature and were intended to describe athlete-specific adaptations rather than support population-level inference. Future research should therefore include larger samples, sex-specific analyses, and more representative competitive settings, while integrating force-time data with kinematic, perceptual, and performance measures.
From an applied standpoint, the findings suggest that braking and propulsive phases represent key windows of sensitivity to task and environmental constraints, albeit in an athlete-dependent manner. Coaches and practitioners may therefore benefit from individualized monitoring of force–time characteristics rather than relying solely on group averages. Future research should integrate waveform-level analyses with performance outcomes and visual behavior measures and explore mixed-effects modeling approaches to better capture the interaction between group-level stability and individual variability.
Conclusions
No significant group-level effects of constraint, shooting position, or their interaction were observed across the unweighting, braking, or propulsive phases of the basketball jump shot, indicating that experienced players generally preserve a stable force–time organization under moderate contextual perturbations. However, intra-individual analyses revealed heterogeneous, athlete-specific responses, primarily during the braking and propulsive phases, which did not generalize across the group. Rather than reflecting systematic adaptations at the population level, these findings suggest that players rely on individualized mechanical strategies to accommodate task demands. Together, this coexistence of population-level stability and individual-level variability highlights the value of hierarchical, phase-specific SPM approaches and supports the potential relevance of individualized, rather than solely group-based, training and intervention strategies when addressing complex sport movements.
Footnotes
Acknowledgements
The authors thank all participants for their involvement in the study. The authors also acknowledge RoboCorp, i2A, the Polytechnic Institute of Coimbra, the Mais Centro Program, and the Center Region Coordination Committee of the EU through the European Regional Development Fund. C.M.A, MAC., and B.G.G. acknowledge the support of the Centre for Mechanical Engineering, Materials and Processes, CEMMPRE, of the University of Coimbra, which is sponsored by Fundação para a Ciência e Tecnologia (FCT) (UIDB/00285/2020, LA/P/0112/2020). C.M.A, R.M., and B.B.G. acknowledge the support of the Research Unit for Sport and Physical Activity, CIDAF, of the University of Coimbra, which is sponsored by Fundação para a Ciência e Tecnologia (FCT) (UIDP/DTP/04213/2020).
Ethics approval
This study was approved by the institutional ethics committee of the University of Coimbra (approval number CE/FCDEF-UC/00812021) and conducted in accordance with the Declaration of Helsinki.
Consent to participate
Written informed consent was obtained from all participants prior to data collection.
Consent for publication
Not applicable.
Author contributions
R.M., MAC and B.B.G. conceptualized the study. C.M.A collected and analyzed the data and drafted the manuscript. R.M., MAC. and B.B.G. supervised the study. R.M., MAC., H.R. and B.B.G. contributed to data interpretation, methodology and data analysis and manuscript revision. All authors reviewed and approved the final manuscript.
Funding
This research was funded by the FCT—Fundação para a Ciência e Tecnologia, grant number 2023.05546.BD.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability
The datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request.