
Abstract
Keywords
Introduction
Neuropathic pain, usually caused by damage to the somatosensory nervous system or harmful stimuli, has affected 7%–10% of the world’s population. 1 For nociceptive pain, opioids and non-steroidal anti-inflammatory drugs have only a modest effect, which make the current lacking of effective drugs to treat neuropathic pain. Thus, it’s necessary to identify underlying pathomechanism of neuropathic pain and to develop precise treatments.
Recently, researchers have found that peripheral nerve regeneration is tightly associated with neuropathic pain. The invalid peripheral nerve regeneration process may be precisely the origin of neuropathic pain. In a rat model of spinal nerve ligation (SNL), both Xie and Fang’s studies2,3 showed that the regeneration process in primary sensory neurons was linked to observed pain behaviors involving peripheral nerve transection. Thus, targeting peripheral nerve regeneration may be a good analgesic strategy. Unfortunately, the current management of neuropathic pain related to peripheral nerve regeneration is far from satisfactory.
Toll-like receptor 4 (TLR4) protects the host cells by recognizing external pathogens and adjusting downstream inflammatory cascades in innate immunity.4,5 Multiple lines of evidence show that TLR4 signaling was involved in different types of pain6–8 and nerve regeneration.9,10 Mechanistically, TLR4 induced the recruitment of myeloid differentiation factor-88 adaptor protein (MyD88) and NF-κB-depended transcriptional process in sensory neurons and glial cells, which produced multiple cytokines and promoted the induction and persistence of pain.11–13 TLR4-MyD88 signaling pathway and downstream TNF-α and IL-1β in the dorsal root ganglion were also involved in nerve injury induced neuroinflammation. 14 In the spinal dorsal horn, TLR4 expressed on astrocytes has been shown to contribute to central sensitization and the persistence of pain by releasing multiple cytokines. 15
Grape seed proanthocyanidin extract (GSPE) are the most abundant source of proanthocyanidins. 16 GSPE intake is reported to be associated with significant decreases of neuropathic pain by selectively inhibiting the activation and/or recruitment of inflammatory cells. 17 After nerve injury, GSPE can significantly attenuate all oxidative parameters such as LPO (lipid peroxide), TBARS (Thiobarbituric Acid Reactive Substances)and nitrate by increasing the number of antioxidants (i.e., SOD (Superoxide dismutase;), hydroBAX and glutathione),18–21which was thought to be a one mechanism on neuropathic pain relief. Other studies also illustrate that GSPE may inhibit the activation of the NOD-like receptors containing a PYD 3 (NLRP3) inflammasome and increase of IL-1β both in models of goat pain and morphine tolerance.14,22 Grape seed-derived procyanidins possess multiple pharmacological properties, including their anti-inflammatory, antioxidant, neuroprotective, and vasodilatory effects, making them promising candidates for treating various types of pain. What’s more noteworthy is their safety profile. Grape seed-derived procyanidins are generally considered safe and well-tolerated when consumed in the diet. Compared to conventional pain medications, such as nonsteroidal anti-inflammatory drugs (NSAIDs) and opioids, procyanidins may offer a safer alternative with fewer side effects and a lower risk of dependency or addiction23,24.In this study, we wonder that GSPE might decrease neuropathic pain and nerve regeneration by suppression of TLR4-Myd88 signaling.
Methods
Animals
Adult female Sprague Dawley rats (180–220 g) were purchased from Hangzhou HangSI Biotechnology Co., Ltd. All rats were housed in a controlled environment (room temperature 21 ± 24°C, lights on 07:00–19:00 h, 4-5 rats per cage) with food and water ad libitum. All experimental procedures were approved by the Institutional Animal Care and Use Committee in Hangzhou Medical College. 32 rats were used in the behavioral testing for mechanical and thermal hypersensitivity testing, respectively. To investigate the impact of procyanidins on neuropathic pain maintenance, we first measured baseline behavior. Rats exhibiting abnormal behavior were excluded, and the remaining rats were randomly divided into four groups, with three groups were subjected to SNL surgery. On postoperative day (POD), 7 the rats were given orally (p.o.) of dissolved GSPE in saline three times per week until POD 28 25 in GSPE1 group (100 mg/kg) and GSPE2 group (300 mg/kg). These three groups have an equal volume of 200 ul per gavage. The dosages of 100 and 300 mg/kg and the oral administration route were selected based on previous studies and considerations of safety, efficacy, and translational relevance.25–27.The rats in the Saline group orally received an equal volume of physiological saline at the same time.All animal experimental procedures in this study were approved (approval number: ZJCLA-IACUC-20020118) by the Institutional Animal Care and Use Committee of Zhejiang Experimental Animal Center. The study was carried out in compliance with the guidelines for humane treatment set by the Center for the Institutional Animal Care and Use Committee of Zhejiang Experimental Animal Center and in accordance with ARRIVE guidelines. At the end of the experience, all mice were euthanized.
SNL model
The SNL model is a representative animal model of peripheral neuropathic pain. 28 Briefly, rats were anesthetized with sodium pentobarbital (30–45 mg/kg, ip), and the left L6 transverse process was carefully removed to expose the L4 and L5 spinal nerves. The L5 spinal nerve was then isolated and tightly ligated with a 5-0 silk thread. The incision was closed with a 3-0 silk thread and disinfected with ethanol and iodophor. For the sham operated group, we removed the L6 transverse process and then the L5 spinal nerve was only exposed and isolated without nerve ligation.
Behavioral analysis
Animals were habituated to the testing environment for at least 2 days before baseline testing. The temperature and humidity were kept stable for all experiments. Rats were placed in plastic chambers and allowed 30 min for acclimation before the examination. As previously described, 29 the mechanical or thermal stimulation was carried out three times to each hind paw at 5 min intervals.
DRG microscopy and DiI tracing
The process of anatomic nerve regeneration was measured by image analysis of regenerated L5 nerve in POD 21 SNL rats using camera in a dissecting microscope. In addition, FAST Dil oil (5 mg/mL in DMSO; catalog #D3899, Thermo Fisher Scientific) was used to determine the degree of nerve regeneration of L5 spinal nerve by checking sections of L5 DRGs to estimate whether Dil had been transported back to the ligated L5 DRG, as described in Xie’s study. 2
Western-blot
The L3-L5 DRG were collected for Western-bolt analysis. The protein concentration were determined by BCA Protein Assay (Thermo Fisher, Waltham, MA). A total of 20 μg of proteins were loaded and separated by SDS-PAGE and transferred onto polyvinylidene fluoride membranes (Millipore Corp, Bedford, MA). The membranes were blocked with 5% bovine serum albumin for 2 h at room temperature, probed with the primary antibodies overnight at 4°C, and then incubated with HRP-coupled secondary antibodies. The primary antibodies used included TLR4 (1:3000, Rabbit, Servicebio) and ACTIN (1:3000, Rabbit, Servicebio). The filters were then developed by enhanced chemiluminescence reagents (PerkinElmer, Waltham, MA) with secondary antibodies (Chemicon, Billerica, MA). Data were acquired with the Molecular Imager (Gel DocTM XR, 170-8170) and analyzed with Quantity One-4.6.5(Bio-Rad Laboratories, Berkeley, CA, USA).
Immunohistochemistry
Rats were deeply anesthetized with sodium pentobarbital and perfused with 0.9% saline followed by 4% paraformaldehyde in 0.1 M phosphate buffer (PB) via the cardiovascular system. The L4/5 spinal segments and L3-5 DRGs were removed and post-fixed overnight. After that, segments were dehydrated using graded sucrose (20% and 30%) in PB for at least two nights until they completely sunk to the bottom. Both DRG and spinal sections (30 μm) were cut on a cryostat and prepared for immunofluorescence staining. Sections were randomly selected and placed into different wells of 6-well plates. After washing with PBS, the sections were first blocked with 0.2% BSA for 2 h at 37°C, and then incubated overnight at 4°C with the following primary antibodies: rabbit anti-GAP43 (1:500, Abcam), rabbit anti-pNF-κB (1:500, Proteintech), rabbit anti-Ki67 (1:500, Abcam), rabbit anti-TLR4 (1:500, Abcam), rabbit anti-MyD88 (1:500, Invitrogen), Mouse anti-GFAP (1:1000, Abcam), Mouse anti-IBA1 (1:500, Abcam). After rewarming, the sections were washed with PBS and incubated with the corresponding secondary antibodies conjugated with Alexa Fluor 488 or 594 (1:1000, CST) for 2 h at 37°C. For double immunofluorescence staining, all the sections were incubated with a mixture of primary and secondary antibodies appropriately. The stained sections were examined and images were captured with a Nikon fluorescence microscope (DS-Qi2, Nikon Co., Tokyo, Japan).
Statistical analysis
SPSS Rel 15 (SPSS Inc, Chicago,IL) was used to conduct all the statistical analyses. Alteration of expression of the proteins detected and the behavioral responses were tested with one-way ANOVA followed by Tukey’s post-hoc test and the differences in latency over time among groups were tested with two-way ANOVA followed by Tukey’s post-hoc test. Bonferroni post hoc tests were conducted for all ANOVA models. Results are expressed as mean ± SEM of three independent experiments. Results described as significant are based on a criterion of p < .05.
Results
Procyanidins reduced pain behavior induced by SNL surgery
To study the analgesic effect of procyanidins in neuropathic pain, the SNL rat model was used as previously described.
25
Two doses of GSPE (100 and 300 mg/kg) were administered orally three times per week after SNL surgery, and the pain behavior were observed until 28 days post nerve injury. The results showed that both mechanical and thermal withdrawal were significantly decreased from 7 to 28 days after surgery compared with administration of saline (Figure 1(a) and (b)). In addition, the two doses of GSPE do not appear to show significant differences in analgesic effects. The time course of surgery, drug administration and behavior tests were showed in Figure 1(c). These data showed that procyanidins can alleviate neuropathic pain. GSPE alleviates mechanical allodynia and thermal hyperalgesia in SNL rats. (a) and (b) Both 100 mg/kg and 300 mg/kg oral administration of GSPE significantly reduced mechanical allodynia (a) and thermal hyperalgesia (b) from 14 to 28 days after surgery compared with that of saline group (n = 8/group). (Two-Way ANOVA followed by Tukey’s post-hoc test, F (15, 120) = 4.664, p < .0001; two-way ANOVA followed by Tukey’s post-hoc test, F (15, 120) = 2.755, p = .0011) (c) The timeline shows GSPE given date, the surgery and tissue harvest date. *p < .05, **p < .01.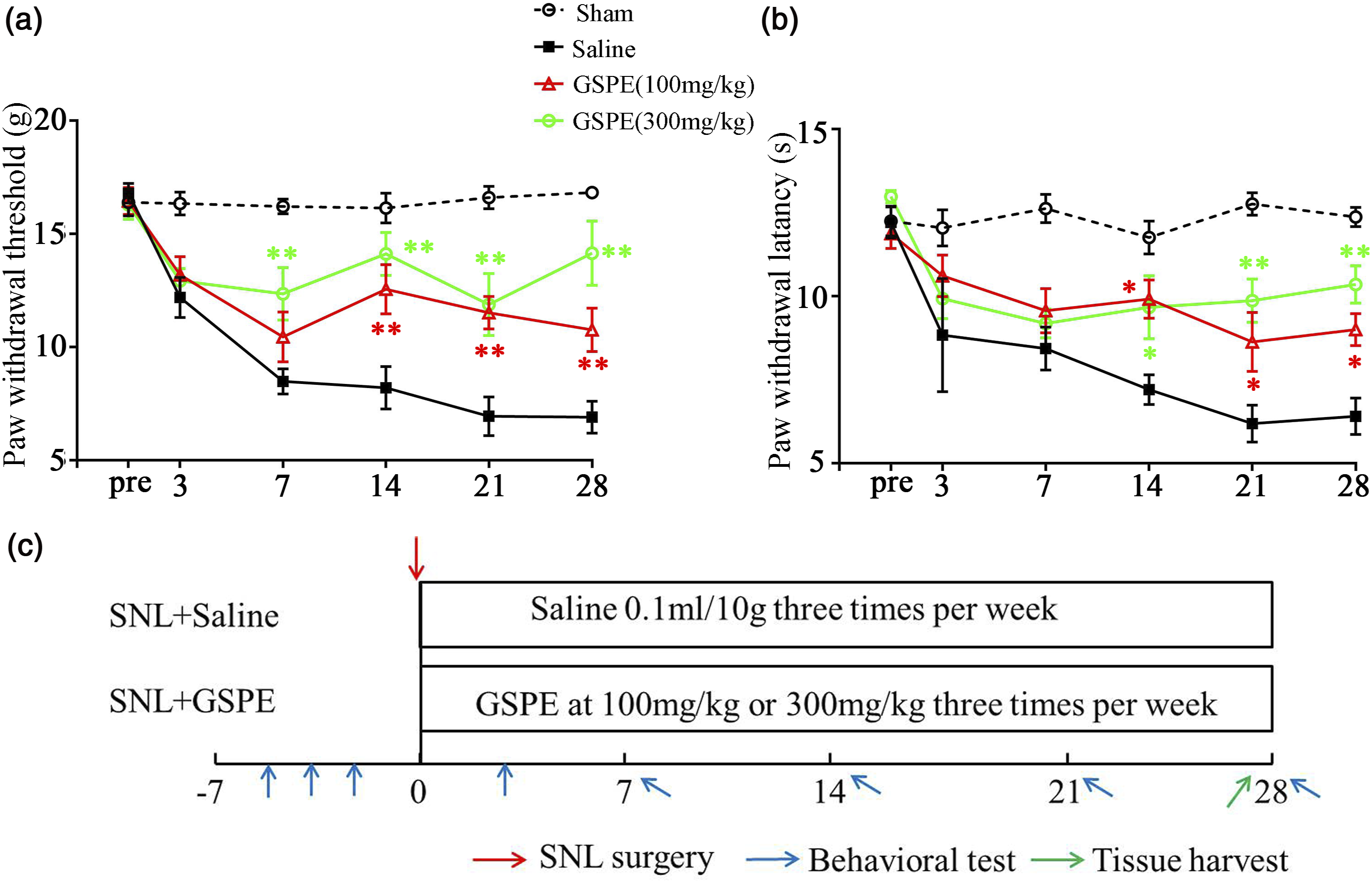
Procyanidins suppressed peripheral nerve regeneration in SNL model
Next, we determined the effect of procyanidins on peripheral nerve regeneration. We also used the SNL model for observation of nerve regeneration as Xie and Fang’s report. Here, obvious L5 nerve regeneration was appeared morphologically 28 days after surgery (Figure 2(a)). GSPE administration inhibited L5 spinal nerve regeneration, while no effect was detected in saline group (Figure 2(b) and (c)). The following DiI tracing results also showed procyanidins blocked DiI transfer to L5 DRG neurons (Figure 2 (d) and (f)), which indicating the suppression effect on nerve regeneration. GSPE suppresses peripheral nerve regeneration. (a)–(c) The images show that spinal L5 nerve regeneration in saline, GSPE (100 mg/kg) and GSPE (300 mg/kg) group, respectively. (d)–(f) The images of DRG sections show Dil tracing in saline, GSPE (100 mg/kg) and GSPE (300 mg/kg) group, respectively. Scale bar, 100 µm.
Procyanidins reduced regeneration-associated molecules
GAP43 is considered a regeneration marker in peripheral axon regeneration.
30
To investigate the effect of procyanidins on regeneration, GAP43 expression was examined by immunofluorescence analysis. As Figure 3(a)–(d) showed, the baseline of GAP43 expression level was low but soared in SNL rats. After GSPE administration, marked down-regulation of GAP43 expression was observed. The summary data in Figure 3(f) also showed a similar tendency. In addition, the Ki67 (a proliferation marker) expression was also reversed by GSPE administration, which showed inhibition effect of procyanidins on peripheral neuron and nerve (Figure 3(f)–(j)). GSPE reduces GAP43 and Ki67 expression in DRGs. (a)–(d) Representative immunofluorescence images of L5 DRG showing GAP43 expression in the sham group (a), SNL group (b), GSPE1 (100 mg/kg) group (c), and GSPE2 (300 mg/kg) group (d). (e) The immunofluorescence intensity analysis of respective groups. Scale bar, 100 µm. (***p = .009, Saline group compared with sham group; ##p = .003, GSPE1 group compared with Saline group; ##p = .0013, GSPE2 group compared with Saline group) (f)–(i) Representative immunofluorescence images of L5 DRG showing Ki67 expression (green) and neuron marker PGP9.5 (red) in different groups. (j) The immunofluorescence intensity analysis. (***p = .001, Saline group compared with sham group; ####p < .001, GSPE1 group compared with Saline group, ###p = .001 GSPE2 group compared with Saline group.)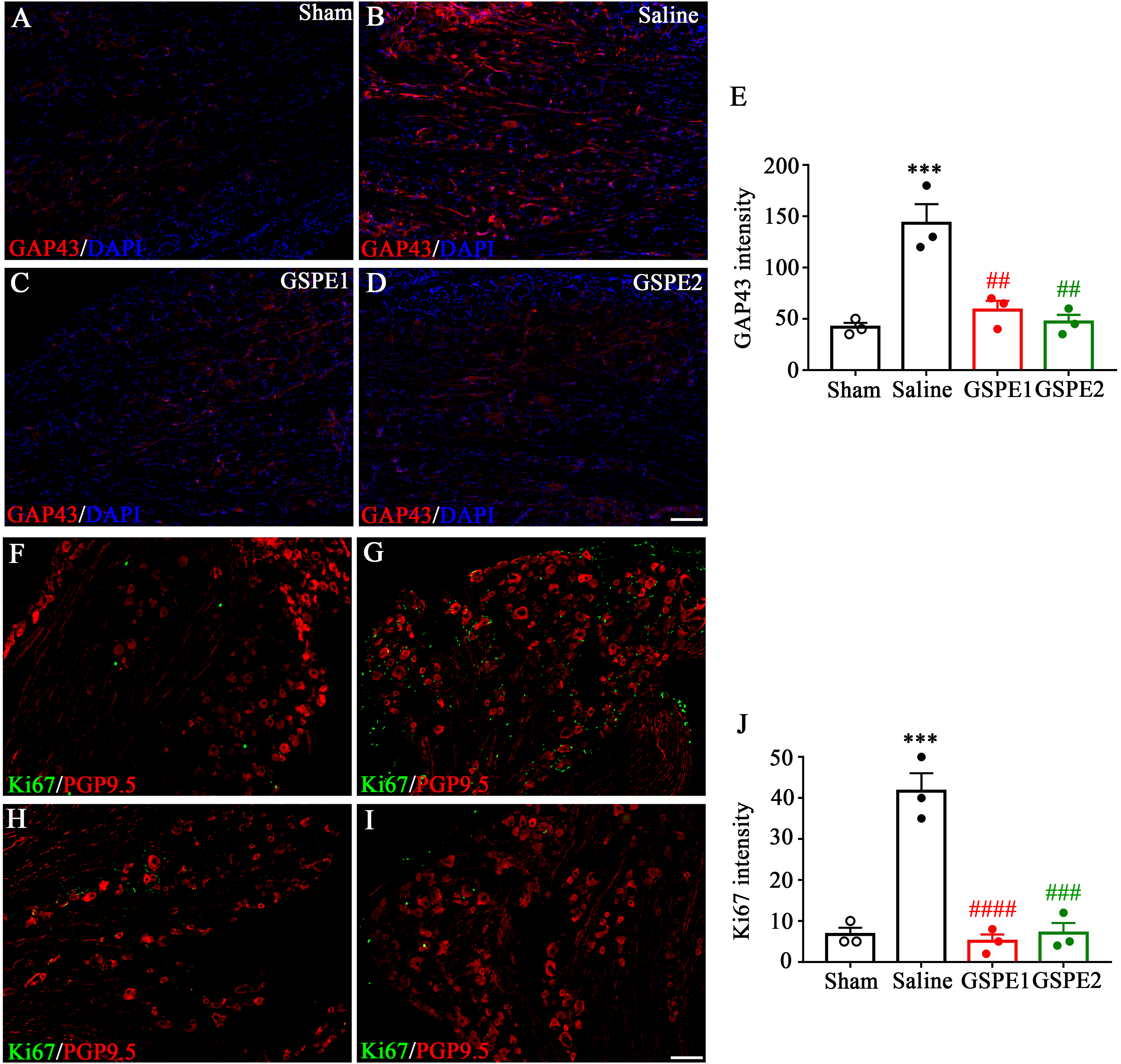
SNL-induced increases in neuronal TLR4 and MyD88 and were reduced by procyanidins adiministration
TLR4 express usually in macrophages and is the core mediator in adaptive and innate immune process. Recent years has reported that TLR4 also expressed in DRG and spinal dorsal horn neurons which play role in neuroimmune. Here, using western boltting analysis, the expression of TLR4 was significantly upregulated in DRG neurons of SNL rats compared with the sham rats. GSPE administration downregulated the high level of TLR4 expression in a dose-dependent manner (Figure 4(a)). The immunofluorescence staining also showed TLR4 located in sensory neurons, co-localizing with PGP 9.5 in the DRG increased in SNL rats and reversed after GSPE administration. Particularly, the TLR4 expression was upregulated in small-diameter neurons of DRG in SNL group compared to sham group, but not in middle- and large-diameter neurons (Figure 4(b)–(f)). GSPE reduces TLR4 expression in DRGs. (a) Representative western-bolt images and statistical analysis of the sham group, SNL group, GSPE1 (100 mg/kg) group, and GSPE2 (300 mg/kg) group. Scale bar, 100 µm. (####p < .0001, GSPE1 group compared with Saline group; ####p < .0001, GSPE2 group compared with Saline group;****p <.0001; Saline group compared with sham group). (b)–(e) Representative immunofluorescence images of L5 DRG showing TLR4 expression (green) and neuron marker PGP9.5 (red) in different groups. (f) The statistical analysis of TLR4-positive neurons in small, medium and large size neurons, respectively.
As the adaptor of TLR4, MyD88 expression was also detected in DRG neurons. The immunofluorescence result showed that, similar to the expression pattern of TLR4, the expression of MyD88 was upregulated in SNL rats and reversed by GSPE only in the small-diameter DRG neurons (Figure 5(a)–(e)). GSPE reduces MyD88 expression in DRGs. (a)–(d) Representative immunofluorescence images of L5 DRG showing MyD88 expression (green) and neuron marker PGP9.5 (red) in different groups. (E) The statistical analysis of MyD88-positive neurons in small, medium and large size neurons, respectively. Scale bar, 100 µm.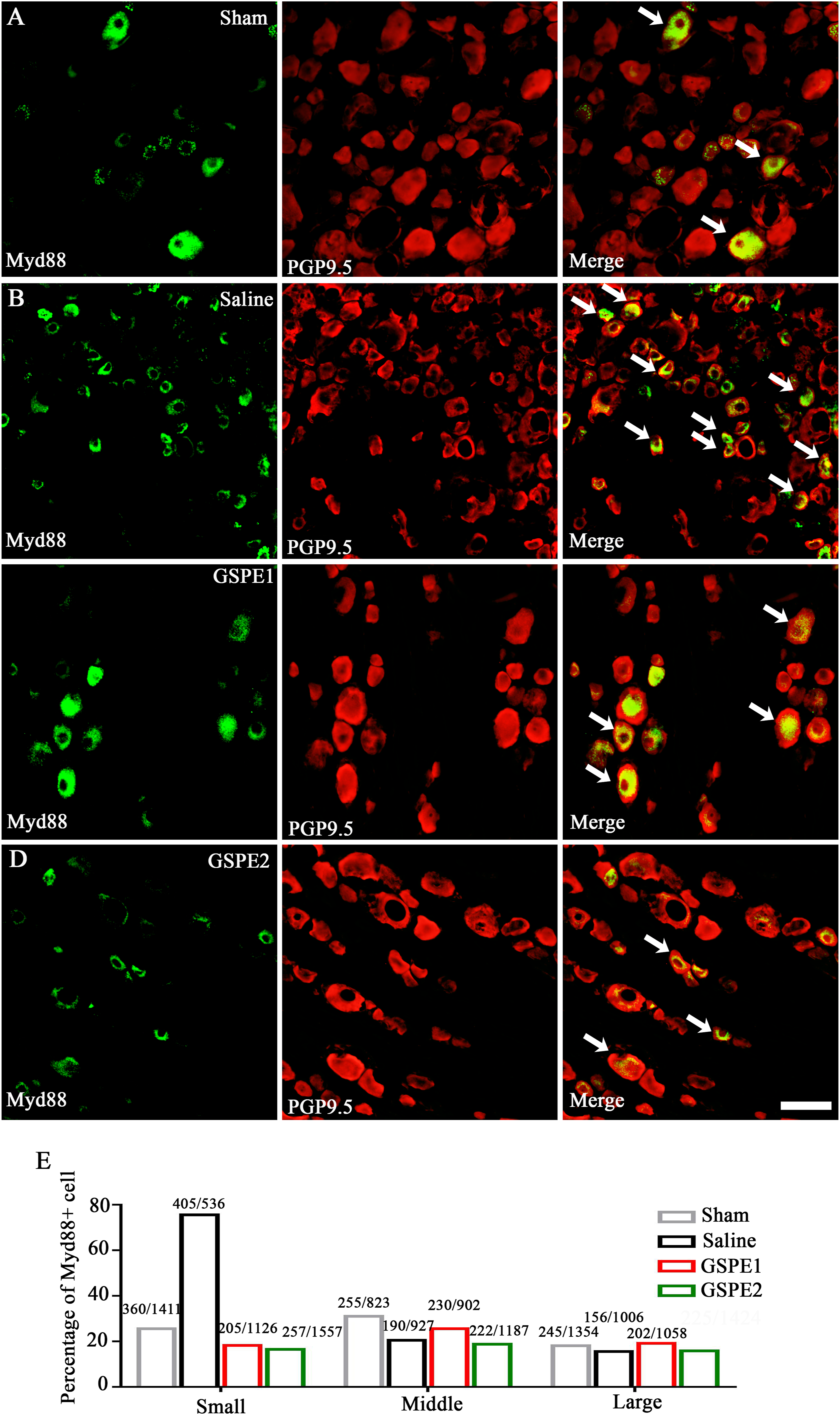
Procyanidins suppressed the activation of NF-κB and macrophage density in DRG and activation of glia in SDH (spinal dorsal horns) in SNL
Macrophage activity and central inflammation have been widely reported that play a role in both neuropathic pain and nerve regeneration. In this study, we detected the macrophage activation using pan-macrophage marker IBA1. As Figure 6(a)–(e) showed, IBA1 intensity increased in SNL 28 days rats and was reversed by GSPE oral administration. As the transcription factor of the TLR4-MyD88 signaling pathway, activated phosphorylated NF-κB was also tested by immunofluorescence analysis. The result showed that phospho-NF-κB p65 increased in DRG of SNL rats. The GSPE administration decreased this tendency with a phospho-NF-κB positive neurons went from 60.5% (451/745 in SNL group) to 25.3% (156/617 in GSPE group) (Figure 6(f)–(i)). Glia cell activation was also detected using the GFAP staining for activated astrocytes and IBA1staining for activated microglia. The immunofluorescence showed that astrocytes and microglia were both activated on ipsilateral SDH compared to contralateral SDH and were weakened after GSPE administration (Figure 7(a)–(h)). The above results indicated that procyanidins suppressed both peripheral macrophage activation and spinal glial cell activation in a TLR4/MyD88/NF-κB pathway dependent manner. GSPE reduces IBA1 and pNF-κB-p65 expression in DRGs. (a)–(d) Representative immunofluorescence images of L5 DRG showing IBA1 expression in the sham group, SNL group, GSPE1 (100 mg/kg) group, and GSPE2 (300 mg/kg) group ***p = .001, Saline group compared with sham group; ###p = .008, GSPE1 group compared with Saline group; ###p = .002, GSPE2 group compared with Saline group. (E) The immunofluorescence intensity analysis of IBA1. (f)–(h) Representative immunofluorescence images of L5 DRG showing pNF-κB expression in the sham group, SNL group and GSPE2 (300 mg/kg) group. (E) The immunofluorescence intensity analysis of pNF-κB. Scale bar, 100 µm. GSPE reduces GFAP and IBA1 expression in spinal dorsal horn. (a)–(c) Representative immunofluorescence images of spinal cord showing GFAP expression in the sham group, SNL group and GSPE2 (300 mg/kg) group. (D) The immunofluorescence intensity analysis of GFAP in both Ipsilateral (Ipsi) GSPE2 (300 mg/kg) group ***p = .0016, Saline group compared with sham group; ###p = .0009, GSPE2 group compared with Saline group. and contralateral (Contra) spinal dorsal horn. (e)–(g) Representative immunofluorescence images of spinal cord showing IBA1 expression in the sham group, SNL group and GSPE2 (300 mg/kg) group. (h) The immunofluorescence intensity analysis of IBA1 in both Ipsilateral (Ipsi) GSPE2 (300 mg/kg) group **p = .0046, Saline group compared with sham group; ##p = .0025, GSPE2 group compared with Saline group. and contralateral (Contra) spinal dorsal horn. *p < .05, **p < .01. Scale bar, 100 µm.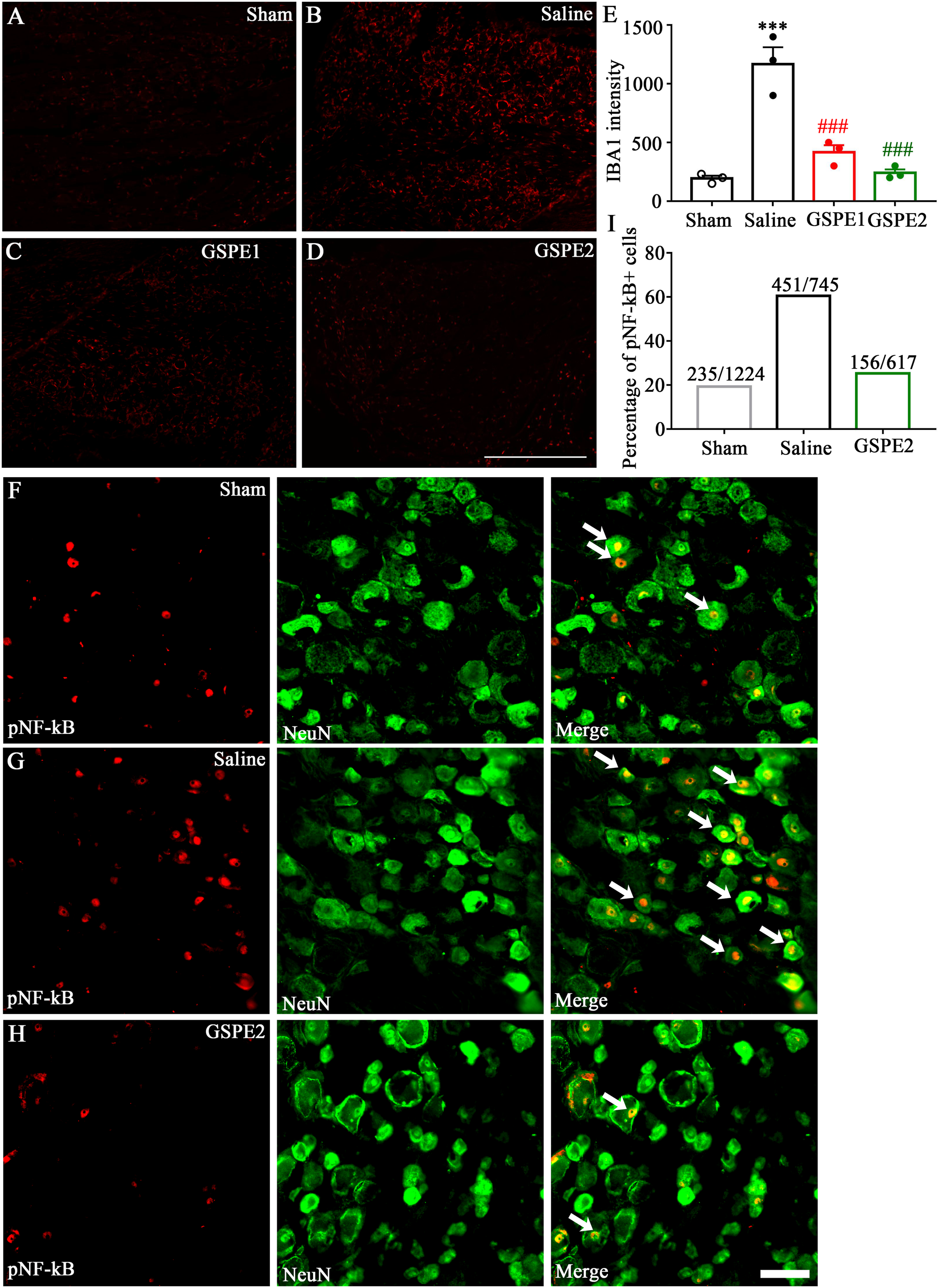

Discussion
In this study, we reported a biological mechanism by which procyanidins inhibits peripheral nerve regeneration and improves neuropathic pain by inhibiting TLR4-MyD88 signaling. Our results demonstrated that clinically used health products, procyanidins, had a significant inhibitory effect to the TLR4-MyD88 signaling pathway, and this suppression contributes to the alleviation of neuropathic pain and reduction of nerve regeneration. Additionally, procyanidins also suppressed activation of spinal astrocytes and glial cells. Our results indicated that Grape seed-derived procyanidins may represent potential candidate drugs for the management of the neuropathic pain.
Procyanidins are natural anti-oxidants and are currently recognized as the most effective free radical scavengers. Recently, Procyanidins in grape seeds and cherries have attracted considerable attention and interest as a promising candidate for the prevention and management of gout, osteoarthritis and related pain.14,21,26 Procyanidins faces a significant challenge due to its low bioavailability, resulting from its large molecular size and low solubility, which leads to poor absorption. 31 This may limit its effectiveness in producing systemic effects, requiring higher doses to achieve the desired outcomes. However, Procyanidins has a high level of biological safety. Acute toxicity studies have shown an LD50 of 4 g·kg−1 in rats, significantly higher than our experimental doses. In subchronic toxicity studies conducted over 90 days, the highest dose group received 2% Procyanidins, equivalent to a daily intake of 1410 mg·kg−1 in male rats and 1501 mg·kg−1 in female rats, without any adverse reactions. 32 Therefore, we can increase the dosage to achieve the desired therapeutic effects. Determining the dosage of grape seed extract typically involves multiple factors, including individual differences, treatment purposes, and formulation of the medication. Factors such as age, weight, health status, and treatment needs can influence the determination of Procyanidins dosage. Procyanidins, as an antioxidant and anti-inflammatory substance, may have varying dosages depending on the treatment purpose; the dosage for improving cardiovascular health may differ from that for anti-inflammatory or antioxidant purposes. Procyanidins can be supplied in various forms, including oral capsules, powder, liquid, or foods containing grape seed extract. Different forms may have different dosages, hence the need to determine based on specific circumstances. Generally, the recommended dosage of Procyanidins for adults ranges from 100 to 1000 mg, but the exact dosage should be determined based on the advice of a doctor or healthcare professional.
As for neuropathic pain, Procyanidins was found to have an analgesic effect in chronic constriction sciatic nerve injury (CCI) model by inhibiting the activation of Matrix metalloproteinase (MMP)-9 and MMP-2. 26 In our study, similar to this finding, intraperitoneal administration of GSPE also attenuated pain behavior in SNL rats from 7 to 28 days after surgery. This result indicates the acute analgesic effect of GSPE, but the long-tern effect is obscure and may be explored in the future research.
Neuropathic pain is usually accompanied by peripheral nerve regeneration. Failed target reinnervation after the nerve injury may lead to persistent neuropathy. 1 As GSPE is highly association with neuropathic pain, we wondered if it also plays a role in nerve regeneration. Here, the GSPE administration not only inhibited nerve regeneration morphologically but also decreased the expression of regeneration marker GAP43 and proliferation marker Ki67. These results revealed that GSPE hindered the process of regeneration, which may partly explain the analgesic effect of procyanidins in another way.
We then investigated the possible mechanisms underlying the analgesia and inhibition of regeneration of procyanidins. TLR family usually play a role in innate immunity and recently it was found that TLR4, mainly expressed in DRG neurons and spinal astrocytes, induces a proinflammatory environment in pain situation through the interplay between MyD88-dependent NF-κB signaling. 33 MyD88 is the adaptor protein for TLR4 and was also found expressed in nociceptors and spinal glia cells which associated with pain process.13,34 In this study, we investigated the activation the TLR4-Myd88-NF-κB pathway in SNL rats and was suppressed by procyanidins. The glia activation in spinal cord was also detected to be inhibited after GSPE administration. These results revealed procyanidins may inhibit peripheral nerve regeneration and alleviate neuropathic pain by inhibiting TLR4-MyD88 signaling. Considering TLR4 may activate MyD88-independent pathways, the effect of other signals could be further studied in the future.
Footnotes
Author contributions
LH, EDA, HJG and HMZ designed the study. LH, EA, ZPZ, YC, XYY and HJG conducted the experiments. LH, EDA, HJG and HMZ analyzed the data. LH, EDA and HMZ wrote the manuscript. All authors have read and approved the final manuscript.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants from the Medical and Health Research Project of Zhejiang Province (2022ZH013).
Ethical statement
Data availability statement
We agree to share our data obtained in the study.