
Abstract
Migraine is the second most prevalent disorder in the world; yet, its underlying mechanisms are still poorly understood. Cumulative studies have revealed pivotal roles of cerebral cortex in the initiation, propagation, and termination of migraine attacks as well as the interictal phase. Investigation of basic mechanisms of the cortex in migraine not only brings insight into the underlying pathophysiology but also provides the basis for designing novel treatments. We aim to summarize the current research literatures and give a brief overview of the cortex and its role in migraine, including the basic structure and function; structural, functional, and biochemical neuroimaging; migraine-related genes; and theories related to cortex in migraine pathophysiology. We propose that long-term plasticity of synaptic transmission in the cortex encodes migraine.
Introduction
Migraine is a disabling neurological disorder characterized by recurrent attacks of moderate to severe headache and a host of neurological, gastrointestinal, autonomic, cognitive, and emotional symptoms.1,2 Ranking as the second most prevalent disorder in the world, migraine has affected 12% of the world’s population and 9.3% of the mainland Chinese population.3–5 Although the initiation of migraine attack is identified to be related to a range of internal and external triggers, such as stress, hormone fluctuations, skipping meals, sleeping disturbances, odor, light, alcohol, and so on, 6 the pathophysiology of migraine remains to be completely understood. 1
The etiopathogenesis of migraine includes various pathological processes and mechanisms involving activation of the trigeminal nerve system, abnormality of brain structure, cortical dysfunction, and vasoconstriction and vasodilation.7–9 However, there is no single hypothesis that can explain all clinical features of migraine. Previous studies have indicated that migraine is a pain disorder associated with abnormal brain structure and function, among which the brain cortex serves a vital role. 8 Cortical hyperexcitability and aberrant resting-state brain activity may underlie the cascade of migraine attacks. 8 Meanwhile, the neural-vascular theory has combined initiation, amplification, and propagation of migraine with vascular reactions as well as responses of cortical neurons. 10 Interfering with cortical activity can affect pain perception, while pain-related cortical regions undergoing changes in synaptic plasticity can generate pain perception even with no detectable sensory input from the periphery. 11
In this review, we aim to summarize the involvement of different cortices (Figure 1) in migraine from several perspectives, including cortex structure and function, structural neuroimaging, functional neuroimaging (including functional magnetic resonance imaging (fMRI), positron emission tomography (PET), and magnetoencephalography (MEG)), biochemical neuroimaging (including magnetic resonance spectroscopy (MRS)), migraine-related genes, neurotransmitters, and peptides. Meanwhile several theories related to cortex in migraine pathophysiology and lately progressed are discussed, including cortical spreading depression, central sensitization, and long-term plasticity of synaptic transmission, which may encode migraine. Pain pathways of migraine in the brain. Peripheral sensory information is first collected by the trigeminal nerve to the trigeminal ganglia (first order neurons in the trigeminal system). Then the trigeminocervical complex, including the trigeminal subnucleus caudalis (TNC) and dorsal horn of the first cervical segments, are the second order central nervous system relay in the trigeminal system, receive input from trigeminal ganglia. Thalamus (Thal.), as the third-order relay in the trigeminal system, receives direct projection from TNC (blue arrows) and modulate the activity of pain-related cortex, such as anterior cingulate cortex (ACC), insula cortex (IC), and primary/secondary somatosensory cortex (S1/S2). Moreover, sensory inputs from periphery are relayed by TNC neurons and further projected to affective/motivational circuits through parabrachial nucleus (PBN, red arrows) and cerebellar cortex (green arrows). A1, primary auditory cortex; Amyg, amygdala; PFC, prefrontal cortex; V1, primary visual cortex.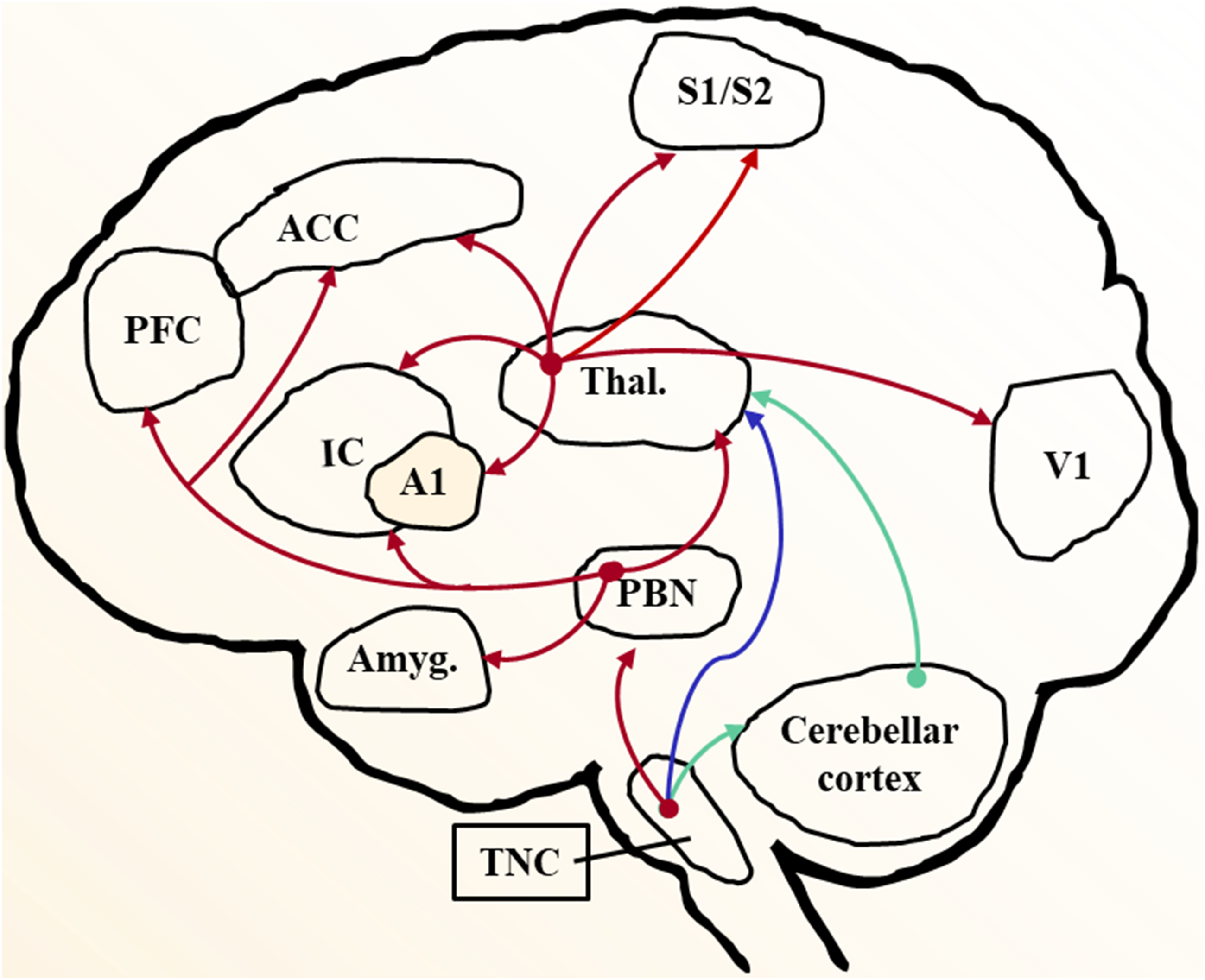
Anterior cingulate cortex
Structure and function
The cingulate is the forebrain structure in mammal, composed of a thick belt of cortex surrounding the corpus callosum. 12 Pyramidal cells in the anterior cingulate cortex (ACC) receive sensory inputs from the thalamus and subcortical areas and project sensory outputs to the motor cortex, amygdala, midbrain areas, periaqueductal gray (PAG), rostral ventromedial medulla, and spinal dorsal horn.11,13–16 The ACC is known to belong to the endogenous opioid pain control circuit and functions in the affective interpretation of pain, cognition, emotion, and motivation.11,17 Projections from ACC to the spinal dorsal horn have been proved to facilitate nociceptive excitatory transmission. 18 The activation of ACC has been indicated to be associated with chronic migraine, acute noxious stimuli, and pain-like aversive behaviors, while inhibition of excitatory transmission in ACC has been suggested to form a blockade of pain-aversive learning, which indicates its pro-nociceptive role.19,20 It is worth mentioning that there are different definitions of the subregions of ACC. For example, according to Vogt, 14 regions in what is typically called the dorsal ACC fall within the anterior MCC.
Structural neuroimaging
Using high-resolution MRI, Maleki et al. 21 demonstrated that migraineurs with more frequent headache attacks showed significant reduced volume of ACC than less frequent ones. Similar findings that decreased gray matter volume in the dorsal ACC was also shown in migraine patients without aura comparing with healthy controls; and these changes were significantly negatively related to the migraine duration. 22
Functional neuroimaging
Functional magnetic resonance imaging
A resting-state fMRI study showed that chronic migraine patients present enhanced connectivity between the right ACC and right amygdala, which as well plays a vital role in the processes of emotion, motivation, learning, and memory. 23 Meanwhile, ACC showed stronger function connectivity to frontal pole, temporal pole, inferior temporal gyrus, the thalamus, and parahippocampal gyrus in migraine patients with more attacks compared with those with less frequent attacks. 21 Another resting-state MR study focused on migraineurs with and without ictal allodynia, with results showing that migraineurs with allodynia had decreased functional connectivity between the PAG to the ACC, prefrontal cortex (PFC), and anterior insula. 24 The decreased functional connectivity results here implied an impairment of the descending pain modulatory pathway, yielding loss of pain inhibition in the migraine process, 24 while the increased functional connectivity between ACC and several higher-order pain-processing cortices and nuclei mentioned before implied an enhanced synchronous neuronal activity of these brain regions, as compensation or cause of migraine process.
Positron emission tomography
In chronic migraine patients, H215O PET study showed that regional cerebral blood flow changes in the ACC, which was positively associated with severity of pain and negatively associated with the paresthesia scores induced by suboccipital stimulation. 25 A FDG-PET study demonstrated hypometabolism in the rostral ACC in episodic migraine patients at rest, which could be relieved after reducing migraine attacks through external trigeminal nerve stimulation. 26 These studies not only implied the role of ACC dysfunction in migraine but also its involvement in the neuromodulation of headache pain.
Biochemical neuroimaging
Using MRS techniques, the metabolites of ACC of migraineurs were studied during the interictal phase, showing slightly reduced level of aspartate, N-acetyl-aspartate (NAA), and glutamine, which did not reach significance. 27 The author, nevertheless, could build a model based on these three metabolites and their ratio to creatine to differentiate migraineurs from healthy controls. 27 Meanwhile, Niddam et al. 28 discovered that chronic, but not episodic migraine, was associated with reduced NAA metabolism in the ACC and the thalamus, as well as with altered interregional NAA correlations between the thalamus and the ACC, reaching the point that a dysfunctional thalamo-cortical pathway was correlated to migraine chronification.
Insular cortex
Structure and function
The insula is a part of the cerebral cortex located deep within the lateral fissure between the frontal and temporal lobe. 29 The anterior insula is part of the salience network with connections to the middle and inferior temporal cortex, ACC, and limbic region, which is related to emotional modulations. 30 The posterior insula has greater connectivity to the premotor, sensorimotor, supplementary motor, and middle posterior cingulate cortex, indicating its role in sensorimotor integration. 30 Nociceptive input is first processed in the posterior insula, where it is coded in terms of intensity and anatomical location. Afterward, it is conveyed to the anterior insula, where the emotional reaction to pain is elaborated. 31 The insula functions in sensory, emotional, salience, and cognition, of which there is a large overlap related to the symptoms involved in migraine. 32
Structural neuroimaging
Significantly reduced volume of the left insula was detected in migraineurs with more frequent headache attacks comparing with less frequent attacks, indicating an adaptive response to the repetitious headache attacks. 21 Similarly, thinning of bilateral insula was observed in chronic migraine patients compared to healthy controls, which was negatively correlated with the frequency of migraine attacks. 33 It has been reported that age-related thinning of the insula was lacking in female migraineurs compared to healthy controls, which may contribute to the overall cortical hyperexcitability of migraine brain. 34 These results also added more proof to the importance of insula in migraine pathophysiology.
Functional neuroimaging
Functional magnetic resonance imaging
Elevated functional connectivity between insula and other pain-processing brain regions and intrinsic networks have been reported. Tso et al. 35 used fMRI to evaluate intrinsic connectivity patterns of migraineurs and discovered stronger connectivity between the dorsal anterior insula and sensory cortex in interictal periods, which might partially explain why visual and auditory stimuli became unpleasant to migraineurs rather than controls even not during an attack. Patients who have spontaneous migraine attacks without aura were observed to show increased functional connectivity between medial PFC and the insula, which was negatively correlated with the intensity of headache. 36 The strength of functional connectivity between the anterior insula and the mediodorsal thalamus as well as the PAG was significantly related to the disease duration in chronic migraine. 37 In addition, migraineurs with aura were indicated to have more fluctuating functional connectivity within the salience network (mainly composed of dACC and anterior insula) compared with migraineurs without aura and healthy individuals. 38 Apart from alterations within the salience network, the insula as well showed enhanced connectivity with the default network and the central executive network in migraineurs. 39 These findings demonstrated that insula may be a critical node in the increased intrinsic connectivity in migraine patients. 39
Positron emission tomography
In H215O PET scans, bilateral insula was indicated to be activated course with other pain-processing cortices either in spontaneous or nitroglycerin-triggered migraine attack.40,41 Whereas, another study with the help of nitroglycerin-triggered migraine showed the right insula was activated in the headache phase but not in the premonitory phase. 42 And right insula activation was also presented in vestibular migraine patients with spontaneous attacks, showing increased metabolisms in temporal-parieto-insular cortices. 43 The author referred this to the activation of vestibulo-thalamo-cortical pathway. 43
Frontal/prefrontal cortex
Structure and function
The frontal lobe contains many dopamine-sensitive neurons and acts as a higher-order pain-processing region in the descending pain modulatory system. Connecting with various neural areas including the medial temporal cortex, hypothalamus, brainstem, and amygdala, the orbital frontal cortex receives sensory input and sends output to various regions for emotional processing and expression. 44 Studies have also indicated that the medial PFC is involved in mediating the attenuation of pain perception via cognitive control mechanisms.45,46 The motor and premotor cortex are parts of pain intensity pathway 47 and play a role in the generation of sensory information along with the supplementary motor area and ACC. 48 Meta-analysis showed that non-invasive stimulation of the excitatory primary motor cortex can significantly reduce intensity and frequency of migraines in migraineurs. 49
Structural neuroimaging
Among patients with migraines, meta-analysis studies of whole-brain voxel-based morphometry (VBM) have shown a widespread reduction in cortex volume, especially in the frontal cortex and cingulate gyrus. 50 One VBM study showed significantly decreased gray matter volume in the left medial PFC in migraineurs without aura. 22
Functional neuroimaging
Functional magnetic resonance imaging
A seed-based MR study showed stronger resting-state functional connectivity between PAG and nucleus cuneiformis to frontal and temporal regions in migraine patients with severe ictal allodynia compared with those without allodynia. 51 Another resting-state fMRI study indicated that bilateral mid-dorsolateral PFC and right ventrolateral PFC presented decreased functional connectivity with PAG in episodic migraine patients and that the mid-dorsolateral PFC may be one of the target brain areas in the impaired PAG dysfunctional network. 16 In episodic migraineurs with higher headache frequency, increased connectivity between the PAG and supplementary motor area was observed, while decreased connectivity between PAG and pain modulation regions such as PFC was shown, indicating high pain expectancy and impaired pain modulatory functions in migraine. 52 Meanwhile, decreased functional connectivity between the left hippocampus, contralateral supplementary motor area, and bilateral inferior parietal gyri has been shown. 53 Experiments showed that increased PFC activation was correlated with trigeminal heat stimulation in migraineurs with cutaneous allodynia, as well as in healthy subjects. 54
Positron emission tomography
Both sides of the frontal cortex have been found to have decreased regional cerebral blood flow related to the headache severity in migraine patients. 55 In addition, one study indicated hypometabolism in the orbitofrontal cortex in episodic migraine patients, which can be relieved by lowering the frequency of migraine attacks with treatment. 26 This hypometabolism was also shown in premotor cortex in patients with episodic migraine. 56
Magnetoencephalography
Several MEG studies showed that acute migraine attacks had higher neuromagnetic activity in the frontal cortex, which was associated with the execution of flexible behavioral responses to various stimuli and can possibly influence the experience of the headache pain process.8,57,58
Temporal cortex
Structure and function
The temporal lobe is located beneath the lateral sulcus and is an associative multisensory area that processes visual, taste, olfactory, and auditory information. 59 It receives projections from prefrontal cortex, insula, thalamus, and raphe nucleus. 60 The temporal pole, which occupies the most rostral part of the temporal lobe, mainly interconnects with amygdala, hippocampus, insula and, in particular, the superior temporal gyrus. 61 The temporal pole is considered to play a role in assigning affective tone to short-term memories related to pain, and integrating multisensory stimuli including painful, visual, auditory, and olfactory. 62
Structural neuroimaging
Gray matter volume in the temporal lobe was shown to be decreased in both adult and pediatric migraine patients. 63 As for interictal periods of migraine, decreased gray matter density in the left temporal pole was detected in patients compared with healthy controls. 64 The interregional cortical thickness correlations of the temporal cortex have also been demonstrated to be able to differentiate the migraineurs’ brain structure from healthy controls in a cross-sectional investigation. 65
Functional neuroimaging
Functional magnetic resonance imaging
In migraine patients, there was enhanced functional connectivity between the temporal pole and several brain areas, such as the ACC, insula, S1, and temporoparietal junction compared to controls, indicating aberrant connectivity of temporal pole might contribute to clinical abnormalities in migraine patients.66–68 Meanwhile, the decreased functional connectivity of the bilateral middle temporal gyrus with PAG, implying disrupted visual information processing in migraineurs. 16 Moreover, it was indicated that the decreased connectivity within temporal cortex of the default mode network in migraineurs without aura in interictal phase. 69 Studies revealed that temporal pole density and functional activity dynamically fluctuated according to the migraine cycle,65,66 yielding it to be a vital site involved in the pathophysiology of migraine, as well as a potential target for treatment.
Positron emission tomography
The temporal cortex was found to have increased activation in migraineurs in response to a noxious heat stimulation in ictal versus interictal states. 66 The activity of temporal pole was enhanced during the migraine attacks compared to the interictal phase, as well as in patients with more frequent attacks than those with fewer attacks.21,70 When enhancing excitability of temporal pole by anodal transcranial direct current stimulation, the migraine patients presented with normal habituation responses to visual stimulation compared with untreatment state. 71 Another PET study showed that the temporal pole of interictal migraineurs had significant increased glucose metabolism in left temporal pole compared to healthy individuals during olfactory stimuli, which reflected the unique role of temporal pole in odor hypersensitivity and odor-triggered migraine. 72 The fusiform gyrus, premotor, and somatosensory cortex were reported to be activated in patients with episodic migraine patients than healthy individuals when painful heat stimulation was applied in interictal period, and the enhanced cognitive pain processing indicates high pain expectancy and hypervigilance in migraine. 73 Both sides of the temporal cortex were found to have decreased regional cerebral blood flow at rest which was related to the severity of headache in migraine patients. 55
Comorbidity with epilepsy
It has been reported that patients with temporal lobe epilepsy showed higher comorbidity of migraine (up to 51.9%), and the removal of the anterior temporal lobe or the hippocampus in these patients could completely relieve the patients from migraine.74–76 And migraine and epilepsy may have some shared pathophysiological mechanisms. Resting-state functional MRI has detected reduced connectivity between PAG and the pedunculopontine nucleus in temporal lobe epilepsy comorbided with migraine. 77 And patients with occipitotemporal lobe epilepsy combined migraine with aura showed monogenic gene defect on chromosome 9q. 75
Parietal cortex
Structure and function
The S1 is located in the postcentral gyrus, responsible for recognizing the localization and intensity of pain. 78 The secondary somatosensory cortex (S2) is located in the parietal operculum and on the ceiling of the lateral sulcus. 79 The S2 is particularly important for pain perception and the processing of pain intensity and quality. 80 Both S1 and S2 receive nociceptive inputs from the spinothalamic pathway via nuclei of the thalamus. 81 These two cortices are also reciprocally connected with posterior parietal and insula through cortico-limbic somatosensory pathway which integrates somatosensory nociceptive inputs with other contextual inputs. 82 It was reported that migraine patients had impaired lateral inhibition in the somatosensory cortex between attacks, which was directly associated with the intensity and duration of migraine attacks. 83
Structural neuroimaging
Studies showed controversial results of thickness change of the somatosensory cortex. Chong et al. 84 reported accelerated age-related thinning in the bilateral somatosensory cortex in episodic migraine. Meanwhile, thinning of bilateral parietal lobes was observed in chronic migraine patients compared to healthy controls, which was negatively correlated with the frequency of migraine attacks. 33 However, a study investigating episodic migraine was found to have a thicker somatosensory cortex in patients with more frequent attacks than whom with lower frequency or no attacks. 21 Another research showed the thickening of somatosensory cortex reached 21% in migraine with aura patients than healthy controls, with most of the patients developed migraine since childhood. 85 Lower gray matter volume was also reported in the S2 of patients with episodic migraine compared with controls. 86 The thickness changes of somatosensory cortex was hypothesized to be the accumulative effects of migraine-specific factors as disease duration, headache frequency, or different circles. 87 The controversial results could also be due to reasons beyond the disease such as limited sample size or different scanning parameters. Further details regarding the controversial volume results are discussed in Occipital cortex section.
Functional neuroimaging
Functional magnetic resonance imaging
There have been various studies providing evidence of altered intrinsic brain connectivity during migraine attacks; using resting-state fMRI, Hougaard et al. 88 showed significant increases in connectivity between the dorsal pons and ipsilateral head and face areas of the S1 during attacks in patients with migraine with aura. Increased S1 activation was observed in migraine patients compared with healthy controls when painful heat stimulation was applied. 73 In addition, activation of S2 was positively associated to cutaneous allodynia in migraineurs without aura detected by fMRI. 54 While some MRI studies based on arterial spin labeling showed diverging results in S1 of headache-free migraineurs, with both hyperperfusion89,90 and hypoperfusion 56 compared with controls, and it was speculated that the resting blood flow changes and alteration in responsivity within S1 may be the result of repetitive attacks rather than the etiology of disease. 90
Magnetoencephalography
Multiple studies have demonstrated that the somatosensory cortex has been continuously undergoing migraine-induced changes.21,85,89,90 One MEG study indicated that migraine patients had increased S1 excitability in response to electrical stimulation compared to controls and that the activation strength was positively correlated to the headache frequency. 91 Hsiao et al. 92 recorded somatosensory gating by MEG and reported that migraineurs presented with excitability changes in S1, which reflected altered sensory modulation associated with migraine chronification.
Occipital cortex
Structure and function
The occipital lobes are located in the forebrain and contain most of the anatomical regions of the visual cortex. 93 As the visual processing center, the occipital cortex is considered to be highly relevant to cortical spreading depression (CSD), migraine aura, and a series of photophobia-like symptoms. 94
Migraine aura and CSD theory
The migraine aura presents in about one-third of migraineurs and is characterized by transient neurological symptoms that generally last no longer than 60 minutes. 95 Its underlying mechanisms are not fully understood, though it is generally acknowledged that the dominant cause of aura is initially associated with CSD that occurred 30–60 minutes prior to the headache attack. 96 In animal models, CSD is characterized by a short-lasting, intense wave of neuronal and glial depolarization, which spreads slowly over the cortex at a rate of 2–4 mm/min, followed by a long-lasting inhibition of spontaneous and evoked neuronal activity. 97 In human beings, CSD is related to short duration of hyperemia and is followed by depression with long-lasting oligemia. 98 The visual, sensory, language, and motor symptoms are also parallel with the wave of altered brain activity which spreads slowly across the cortex. 99
Structural neuroimaging
Decreased gray matter volume of occipital lobe has been reported in migraine patients without aura. 22 Similar findings were shown in right occipital lobe of chronic migraine patients complicated with medication overuse. 100 Another study based on general population has showed significantly decreased gray matter volume in occipital lobe, mainly in visual motion processing areas, regardless of migraine activity. 101 A recent study using novel coordinate-based network mapping method discovered that heterogeneous areas of decreased gray matter volume in migraine were commonly connected to a single region in the left extrastriate visual cortex. 102 Regarding to the VBM studies in migraine, many different brain regions or controversial results of the same region were both reported; and the coordinate-based network mapping method could be used and tested in the future study. It may provide a chance to answer whether volume change in migraine brains is the cause, result, epiphenomenon, or just false positive results in migraine disease.
Functional neuroimaging
Functional magnetic resonance imaging
After documenting a patient for more than 1000 migraine aura attacks, Hansen et al. 103 suggested it is possible for aura to initiate in multiple sites in the visual cortex and that the aura could cease spontaneously with limited propagations. Arngrim et al. 104 investigated five migraine patients using fMRI during provoked aura, showing that in the visual cortex negative symptoms (scotoma) were associated with decreased mean blood-oxygen-level-dependent response while positive symptoms (flickering) were associated with increased blood-oxygen-level-dependent response. These findings indicated that CSD could induce different effects on the brain activity, metabolism, and hemodynamics and presenting inverse aura symptoms. 104
Positron emission tomography
One study reported decreased metabolism and increased activation-to-resting glucose uptake ratio of neurons in the visual cortex of migraine patients during the interictal phase. 105 In another study using luminous stimulations, the visual cortex was activated as a response to pain in interictal and postictal phases, but deactivated in ictal phase of migraineurs, indicating that in cases of increased trigeminal nociceptive input, the pain-related visual cortex activity might subsequently be higher and lead to lesser responses to additional pain stimulation. 106 A recent PET/MRI study using「11C」PBR28 as marker for glia activation has proved increased standard uptake value ratio in the S1, S2, insula cortex, and thalamus as well as visual cortex, linking CSD to glia activation and neuroinflammation. 107
Magnetoencephalography
Hyperexcitability of the occipital cortex was observed in spontaneous and visually induced migraine patients, with smaller widespread regions beyond the primary visual cortex in spontaneous subjects. 108 MEG was also used to evaluate treatment response, reporting hyperexcitability of the occipital cortex to visual stimulation in migraineurs, and reduced DC-MEG shifts after 30 days of valproate therapy. 109
Biochemical neuroimaging
Imbalances of metabolic and neural transmitters have been reported in the occipital lobe of migraine patients. Recently, MRS studies showed higher lactate levels in the occipital visual cortex in migraine with aura patients during interictal period 110 . It is found that when facing checkerboard stimulation during interictal phase, migraine aura patients who have both visual and non-visual aura (paraesthesia, paresis or dysphasia) showed lactate increase only during stimulation and only in visual cortex; however, patients with visual aura showed higher level of lactate before stimulation, and no more increase during stimulation, indicating mitochondria dysfunction in the patients. 111 Additional research focusing on the visual cortex showed consistently decreased NAA levels and increased lactate levels during visual stimulation in migraineurs with aura compared to patients without aura and normal controls. 112 These findings indicated that mitochondrial dysfunction might only present in migraine with aura. Furthermore, a 1H-MRS study showed that glutamate levels were increased 113 while gamma-aminobutyric acid (GABA) levels were decreased 114 in the visual cortex in migraine patients with aura. Similar 1H-MRS study also reported higher ratios of glutamate/creatine 115 and glutamate/glutamine 116 in the visual cortex of migraineurs. These findings suggested disturbances in the cortical excitatory-inhibitory balance, which could predispose the cortex to CSD and aura. 117
Cerebellar cortex
Structure and function
The cerebellum has been demonstrated to have vast connections with migraine-related brain regions such as cerebrum, thalamus, midbrain, brainstem, and spinal cord.118,119 Furthermore, the cerebellum has been implicated in motor control and coordination, 120 cognition, 121 emotion, 122 and pain processing.123,124 The trigeminal spinal nucleus also directly projects to precerebellar regions such as the pontine nuclei and inferior olive. 125 The deep cerebellar nuclei receives inhibitory inputs from Purkinje cells in the cerebellar cortex, and excitatory inputs from the mossy fiber and the climbing fiber pathways, projects to the brainstem nuclei and thalamus, then finally terminates in the cerebral cortex. 123
Structural neuroimaging
MRI studies have indicated decrease volume of the cerebellar cortex in chronic migraine patients compared with normal controls. 126 Recent VBM study indicated significant decreased gray matter volume in the cerebellum in migraine patients with or without aura than healthy subjects. 127 A longitudinal study showed that cerebellar structural volume and connectivity changes were correlated to the 2-year headache frequency prognosis in migraineurs. 128 Furthermore, postmortem neuropathological examination of a familial hemiplegic migraine patient revealed cerebellar vermis atrophy and reduction of Purkinje cells and granular cells alongside proliferation of Bergmann glia. 129
Functional neuroimaging
Functional magnetic resonance imaging
Studies showed decreased functional connectivity between crus I of the cerebellum and the thalamus, as well as the fusiformis gyrus in migraine patients. 130 The author proposed the aberrant reduced inhibitory involvement of the migraine cerebellum on gating and evaluation nociceptive sensory information. 130 Furthermore, the activation of the cerebellum in response to nociceptive input was higher during the migraine attack than in interictal periods. 70 These functional studies add proof to the cerebellar dysfunction and cerebellum involvement in migraine. Here, we would like to add some results other than fMRI studies. Cerebellar hypoperfusion has been reported in migraine patients,131,132 and its relationship to the cause of cerebellar infarct-like lesions in migraine showed controversial results.132,133 The CAMERA (Cerebral Abnormalities in Migraine, an Epidemiologic Risk Analysis) study indicated that prevalence of infarct-like lesions in the cerebellar region of migraine patients were significantly higher than that in controls (5.4% vs 0.7%, p = .02, adjusted OR, 7.1). 134 In addition, crossed cerebellar diaschisis has also been observed in migraine patients and is considered to pose a probable risk of infarctions and further studies are required.133,135
Positron emission tomography
Studies have indicated activation of the cerebellum during the various phases and in different subtypes of migraine. H215O PET study with the help of nitroglycerin-triggered migraine showed that cerebellum activation exists in the premonitory phases of migraine. 42 One 18F-FDG-PET study demonstrated increased metabolism in the bilateral cerebellum in the ictal phase compared with interictal phase in vestibular migraine patients. 43
Biochemical neuroimaging
MRS studies showed significantly reduced NAA and glutamate, with elevated myo-inositol in cerebellum of patients with familial hemiplegic migraine-type 1 (FHM-1), and the NAA level was negatively correlated with cerebellar dysfunction (gait ataxia scores). 136 Another single-voxel MRS research demonstrated significantly decreased choline levels in the cerebellum by comparing patients of migraine with aura and healthy controls. 137
Migraine and related cerebellar dysfunction
Several studies have suggested that migraine might be related with cerebellar dysfunction.138–143 The cerebellar cortex exhibited rich expressions of CGRP and CGRP receptor components. 144 The high concentration of CGRP in Purkinje cells with receptor elements in fibers indicates a functional role of CGRP in the cerebellum. 145 It was concluded that nearly 20% of hemiplegic migraine patients showed permanent mild cerebellar symptoms. 146 FHM-1 is a type of headache disorder caused by mutations in CACNA1A (Calcium Voltage-Gated Channel Subunit Alpha1 A) gene, which codes for the α1 subunit of a neuronal P/Q-Ca2+ channel. 146 The P/Q-Ca2+ channels are widely expressed in the central nervous system, primarily in the Purkinje cells, 147 and this mutation has also been implicated in causing episodic ataxia type-2. 148 Apart from FHM-1, there are three other migraine types in which the cerebellum was involved, including migraine with aura, migraine without aura, and basilar-type migraine. Several cerebellar symptoms including ataxia, dysarthria, oculomotor deficits, and dizziness have been observed in migraine.135,149 Balance abnormalities have been detected by stabilometry studies during ictal and interictal periods in untreated migraine patients.139,150 Subclinical cerebellar dysfunctions such as hypermetria have also been shown in both migraine with and without aura, but were more prominent in migraine with aura. 138
A hypothesis: Long-term plasticity in the cortex contributes to migraine
Here, we would like to propose a hypothesis that long-term plasticity (the main mechanism of cortical sensitization, including presynaptic and postsynaptic plasticity) in the cortex encodes migraine (Figure 2). There are many hypotheses trying to explain the etiopathogenesis of migraine, such as activation of the trigeminal nerve, cortical dysfunction, vasoconstriction and vasodilation, and CSD.151,152 Although there is no single hypothesis that can explain all features of migraine, the sensitization of central nervous system is often noted (such as the increased activation of ascending sensory pathways) in migraine.153,154 Due to the synaptic plasticity in cortices is the critical molecular mechanism of chronic pain and its related emotional responses,
20
we believe that the cortical plasticity provides a synaptic model for studying the pathogenesis of migraine. Synaptic model for long-term potentiation (LTP) in cortex. The hyperexcitability of presynaptic neurons leads to increased release of glutamate, which combines with postsynaptic NMDA receptors (NMDARs), and causes increased postsynaptic Ca2+ in dendritic cells. Ca2+ serves as an important intracellular signal for triggering a series of biochemical events that contribute to the expression of AMPA receptors (AMPAR) and LTP. Under migraine conditions, calcitonin gene-related peptide (CGRP) might serve as another important signaling molecule for activating adenylyl cyclase 1 (AC1) and contributing to the expression of LTP. Furthermore, interneurons can inhibit or disinhibit the activity of postsynaptic neurons by releasing γ-aminobutyric acid (GABA). GABAAR, γ-aminobutyric acid type A receptor; cAMP, cyclic AMP; CaM, calmodulin; PKA, protein kinase A; MAPK, mitogen-activated protein kinase; CaMKIV, Ca2+-calmodulin-dependent protein kinase IV; CREB, cAMP response element-binding protein; CRE, cAMP response element.
Somatosensory diffusible allodynia and anxiety, which have been observed in both migraine patients and animal models, are two typical symptoms of migraine.153,155–157 This indicates that central modulation may be the key part for migraine. However, most previous studies focused on trigeminal ganglion (TG) and thalamus.153,154,158,159 Less is known about the possible roles of cortices (such as ACC and insula), which are essential for pain perception and emotional responses in the migraine.13,14,20
Studies from rodents with peripheral injury have shown that the synaptic plasticity in cortices (such as ACC and insula) contribute to the allodynia and pain-associated emotions (such as anxiety and depression).20,160 Similarly, increased neural activities in response to noxious stimulation in interictal migraineurs have also been observed in the cortices involved in cognitive aspects of pain processing and top-down modulation of pain.153,154 CSD is now thought to be a precursor to migraine attacks. 161 Changes in cortical sensory response after CSD suggest the possible involvement of known cortical plasticity in migraine. Specifically, low CSD threshold enhances cortical susceptibility, while cortical excitation can trigger CSD by increasing the release of presynaptic glutamate (related to injury-associated anxiety) and CGRP and enhancing postsynaptic NMDA receptor (NMDAR)–mediated responses (related to injury-induced allodynia).162–165 One recent piece of evidence from mice with familial hemiplegic migraine has shown that the impairment in glutamate uptake promotes NMDA spike generation in ACC neurons and enhances the output firing of these neurons. These findings further support the view that the known plasticity in cortices is involved in migraine. In parallel, this study also shows that local rescue of the impairment in glutamate uptake in ACC can prevent abnormal NMDA spikes and reduces sensitivity to cranial pain triggers in migraine mice. 166 Recently, we found that cortical plasticity contributes to periorbital allodynia and anxiety in rats suffering chronic migraine, in which both glutamate release and NMDAR responses in ACC are increased (unpublished data). Furthermore, our recent slice experiments have shown that CGRP induces a long-term change of α-amino-3-hydroxy-5-methyl-4-isoxazole-propionic acid (AMPA) receptor–mediated responses in the ACC and insula.167,168 This may explain why infusion or overexpression of CGRP receptors in rodents can cause migraine and photophobia.169,170
In conclusion, these findings from cortical plasticity and migraine provide a new direction to explain many symptoms of migraine, such as allodynia, anxiety, and photophobia. However, it is obvious that the current research on cortical plasticity in the regulation of migraine is still insufficient. Therefore, future studies are required to explore the specific mechanism of cortical plasticity in the regulation of migraine.
Conclusion
Migraine is a multiphasic disorder that involves not only intermittent paroxysmal pain but also influences different regions of the brain over time. Various brain cortices act as a whole and perform different roles during this process. More direct evidence and high-quality research should be attained during various phases of migraine in the future. Meanwhile, methods which could analyze the neuroimaging across different parameters and combine the results should be performed. The combination of different experimental approaches such as neuroimaging and electrophysiology studies should be carried out to enhance the base of knowledge and explore yet-unanswered questions.
Footnotes
Acknowledgments
The authors would like to thank Melissa Lepp for English editing.
Author contributions
All authors participated in the interpretation of collected literature, and in the drafting, critical revision, and approval of the final version of the manuscript.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the National Science Foundation of China (grants 81671077, 82071226), Beijing Natural Science Foundation Essential Research Project (Z170002), Hainan Provincial Natural Science Foundation of China (818MS153).