
Abstract
Cancer-induced bone pain is one of the most severe types of pathological pain, which often occurs in patients with advanced prostate, breast, and lung cancer. It is of great significance to improve the therapies of cancer-induced bone pain due to the opioids’ side effects including addiction, sedation, pruritus, and vomiting. Sinomenine, a traditional Chinese medicine, showed obvious analgesic effects on a rat model of chronic inflammatory pain, but has never been proven to treat cancer-induced bone pain. In the present study, we investigated the analgesic effect of sinomenine after tumor cell implantation and specific cellular mechanisms in cancer-induced bone pain. Our results indicated that single administration of sinomenine significantly and dose-dependently alleviated mechanical allodynia in rats with cancer-induced bone pain and the effect lasted for 4 h. After tumor cell implantation, the protein levels of phosphorylated-Janus family tyrosine kinase 2 (p-JAK2), phosphorylated-signal transducers and activators of transcription 3 (p-STAT3), phosphorylated-Ca2+/calmodulin-dependent protein kinase II (p-CAMKII), and phosphorylated-cyclic adenosine monophosphate response element-binding protein (p-CREB) were persistently up-regulated in the spinal cord horn. Chronic intraperitoneal treatment with sinomenine markedly suppressed the activation of microglia and effectively inhibited the expression of JAK2/STAT3 and CAMKII/CREB signaling pathways. We are the first to reveal that up-regulation of microglial JAK2/STAT3 pathway are involved in the development and maintenance of cancer-induced bone pain. Moreover, our investigation provides the first evidence that sinomenine alleviates cancer-induced bone pain by inhibiting microglial JAK2/STAT3 and neuronal CAMKII/CREB cascades.
Introduction
As one of the most severe types of chronic pain, cancer-induced bone pain (CIBP) is an intricate condition which involves allodynia, hyperalgesia, and spontaneous pain.1,2 The reported mechanisms underlying CIBP are associated with spinal cord plasticity, glial cell activation, and inflammatory cytokine signaling.3–5 Despite the availability of diverse analgesic agents, including opioids, non-steroidal anti-inflammatory drugs (NSAIDs), and adjuvant drugs, further elucidation of fresh therapy of CIBP is necessary because of their side effects.6–8 Although there have been marked advances in recent years, mechanisms of CIBP are far from clear. Therefore, further exploration of the underlying mechanism for novel therapies is still demanded.
Sinomenine is a morphinan derivative extracted from the traditional Chinese medicine Sinomenium acutum. 9 It has been shown to possess a variety of pharmacological effects, such as anti-inflammatory, anti-angiogenesis, antiarrhythmic, and immunosuppressive properties.10–13 A large number of preclinical and clinical studies have confirmed that sinomenine is effectively against rheumatoid arthritis, arrhythmia, and glomerular diseases.14–16 Recently, there was evidence suggesting that sinomenine alleviated mechanical allodynia and heat hyperalgesia in the models of neuropathic and inflammatory pain.17–19 In addition, sinomenine was demonstrated to attenuate brain injury by inhibiting microglial activation in intracerebral hemorrhages. 20 However, the potential analgesic effect of sinomenine in CIBP has never been reported.
Considerable research has suggested that numerous extracellular signaling molecules and intracellular transduction pathways are involved in chronic pain.21–24 The Janus family tyrosine kinase (JAK)/signal transducers and activators of transcription (STAT) pathway is one of the most important cell signaling pathways in chronic pain, which includes four types of JAK and seven types of STAT.25,26 Our previous study has demonstrated that STAT1 contributes to CIBP by regulating MHC II expression in spinal microglia. 27 Accumulating evidence suggests that the activation of the JAK2/STAT3 pathway contributes to the development of neuropathic and inflammatory pain.28–31 After nerve injury, the expression of p-STAT3 is significantly up-regulated and leads to a neuroinflammatory response. 32 In addition, the JAK2/STAT3 pathway was activated after chronic treatment of morphine. 33 In the model of ischemic stroke, sinomenine was reported to alleviate neuroinflammatory injury via the STAT3 pathway. 34 However, the role of the JAK2/STAT3 pathway has never been demonstrated in model of CIBP.
Ca2+/calmodulin-dependent protein kinase II (CaMKII) is a multifunctional serine/threonine kinase which is involved in synaptic plasticity, learning, and memory.35–37 The activation of CAMKII was proven to participate in cyclic adenosine monophosphate response element-binding protein (CREB) phosphorylation.38,39 The activated CaMKII/CREB pathway has been extensively confirmed in different pain models.28,40,41 In an inflammatory pain model, sinomenine was reported to attenuate mechanical allodynia and thermal hyperalgesia by down-regulating the CAMKII/CREB pathway. 19 Novel inhibitors targeting the CAMKII/CREB pathway may be an alternative therapy for CIBP.
In the present study, we hypothesized that sinomenine exerts its antinociceptive effects on CIBP through suppressing neuronal CAMKII/CREB and microglial JAK2/STAT3 pathways.
Material and methods
Ethics statement
All experimental protocols were approved by the Animal Care and Use Committee of Huazhong University of Science and Technology and were in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals and the guidelines of the International Association for the Study of Pain. 42 All procedures were made to minimize the number of animals used and their suffering.
Animals
Because female rats are more susceptible to Walker 256 mammary gland carcinoma cells, all of the experiments were performed on virgin female Sprague Dawley rats (180–200 g, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, People’s Republic of China) in this study. Rats were kept in controlled conditions (24 ± .5C, 12-h alternating light-dark cycle, with ad libitum access to water and food).
Drug administration
Sinomenine was purchased from Selleck Chemicals (Houston, TX) and dissolved in saline. Different doses were injected intraperitoneally (i.p) based on previous reports.9,43 The drug administration protocol was as follows: for acute treatment, sinomenine (i.p, 10 mg/kg, 20 mg/kg, and 40 mg/kg) or vehicle (i.p) was administrated on day 14 after CIBP model establishment; for chronic treatment, sinomenine (i.p, 10 mg/kg, 20 mg/kg, and 40 mg/kg) or vehicle (i.p) was administrated from day 14 to day 18 once daily after tumor cell implantation (TCI).
Preparation of carcinoma cells
Walker 256 rat mammary gland carcinoma cells (4 ×107 cells/mL, 1 mL) were inoculated into the abdominal cavity of female rats as previously described. 44 After seven days, carcinoma cells were extracted from the ascitic fluid and were diluted to the final concentration (4 ×107 cells/mL) using a hemocytometer. The cell suspension was maintained on ice until inoculation.
Cancer-induced bone pain model establishment
The model of CIBP was carried out as described previously.45,46 After anesthesia with pentobarbital sodium (50 mg/kg, i.p), the right leg of rat was shaved and disinfected. Then, a minimal incision was made to expose the proximal tibia and the prepared Walker 256 cells (4×107 cells/ml, 10 µl) were slowly injected into the bone cavity. For the sham group, 10-µl D-Hank’s solution was injected instead. The injected site was immediately closed using bone wax after the syringe was removed. Finally, the incision was sutured using 3-0 silk thread.
Behavioral assessment
Pain-related behaviors were measured on day 0, 3, 7, 14, and 21 after the injection of tumor cells. Mechanical allodynia was assessed using ipsilateral hind paw withdraw threshold (PWT) to von Frey filament as described previously.27,47 Rats were placed in individual plastic boxes with a metal mesh floor and allowed to acclimate for half an hour. A range of von Frey filaments (1, 1.4, 2, 4, 6, 8, 10, and 15 g), with ascending order, were applied to determine the mechanical PWT. Quick paw withdraw and licking were considered positive responses. When a positive response occurred, rats were allowed to have a rest for 5 min before the next descending von Frey filament. The lowest amount of force which induced a positive response was recorded as the PWT.
Immunohistochemistry
After deep anesthesia with pentobarbital sodium (60 mg/kg, i.p), the rats were perfused intracardially with saline followed by 4% ice-cold paraformaldehyde in 0.1 M phosphate-buffered saline. The enlargement of spinal cord (L4-L6) was removed and postfixed in 4% paraformaldehyde for 4 h and subsequently dehydrated in 30% sucrose for 24 h at 4°C. The segments were sectioned 20 µm thick in a cryostat (CM1900, Leica, Wetzlar, Germany). To confirm the cell type that expressed p-JAK2, the rabbit anti-phospho-JAK2 antibody (1:1000; #3776; Cell Signaling Technology) was mixed with mouse anti-neuronal nuclei (NeuN) antibody (neuronal marker; 1:200; MAB377; EMD Millipore, Billerica, MA), mouse anti-glial fibrillary acidic protein (GFAP) antibody (astrocytic marker; 1:300; 3670; Cell Signaling Technology, Danvers, MA), or goat anti-ionized calcium-binding adapter molecule 1 (Iba1) antibody (microglial marker; 1:300; ab5076; Abcam). Iba1-immunolabeled surface area was quantified from the spinal dorsal horn using Image Pro Plus software version 6.0. Quantification of Iba1immunoreactivity was accomplished by calculating the percentages of immunostaining ((positive immunofluorescent surface area)/(total measured picture area) × 100). Four rats of each group were used for statistical analysis.
Western blot
Under deep anesthesia with pentobarbital sodium (60 mg/kg, i.p), the L4-L6 spinal segments were collected and homogenized in a radioimmunoprecipitation assay lysis buffer containing 50 mM Tris (pH 7.4), 150 mM NaCl, 1% TritonX-100, 1% sodium deoxycholate, 1% sodium dodecyl sulfate, 1 mM NaF, and 2 mM ethylenediaminetetraacetic acid. The supernatants were collected and protein concentrations were determined by the Bradford method. The protein samples were heated at 95° for 10 min in a loading buffer. Equal amounts of proteins (40–60 µg) were separated by 10% sodium dodecyl sulfate-PAGE and transferred onto polyvinylidene fluoride membranes (IPVH00010; EMD Millipore). After blocking with 5% BSA in 0.1% TBST for 2 h at room temperature, the membranes were incubated overnight at 4°C with rabbit anti-phospho-CAMKII antibody (1:3000; ab5683; Abcam), rabbit anti-phospho-CREB (pSer133) antibody (1:500; MA5–11192; Thermo), rabbit anti-phospho-JAK2 antibody (1:1000; #3776; CST), rabbit anti-phospho-STAT3 antibody (1:1000; #9145; CST), mouse anti-Iba1 antibody (1;500; ab15690; Abcam), and rabbit anti-GAPDH antibody (1:5000; AS1039; Aspen), respectively. The membranes were washed in TBST and incubated with HRP goat anti-rabbit (1:5000; A21020; Abbkine) or goat anti-mouse secondary antibody (1:5000; A21010; Abbkine) for 2 h at room temperature. The protein bands were finally visualized using SuperLumia ECL Plus HRP Substrate Kit (K22030; Abbkine) and then measured by a computerized image analysis system (BIO-RAD, ChemiDoc XRS+, USA).
Statistical analysis
All data are presented as the mean ± standard error of the mean (SEM) and performed using GraphPad Prism version 5.01 for Windows (Graph Pad Software, San Diego, CA). Western blot and immunochemistry data were evaluated via one-way analysis of variance followed by a Bonferroni post hoc test. The differences in the mechanical PWT data were analyzed using two-way analysis of variance with repeated measures, followed by a Bonferroni post hoc test. p < 0.05 was considered statistically significant.
Results
Antinociceptive effects of sinomenine in cancer-induced bone pain
Before cell transplantation, there was no significant difference in pain-related behaviors among the sham and other groups. Consistent with our previous studies, the ipsilateral PWT of CIBP group decreased dramatically from day 7 to day 21 after TCI (Figure 1(a)). The results indicate that TCI leads to the mechanical allodynia.

To investigate the analgesic effect of single administration, we injected different doses of sinomenine (i.p, 10 mg/kg, 20 mg/kg, and 40 mg/kg) and vehicle on day 14 after TCI and tested the PWT at 0, 0.25, 0.5, 1, 2, 4, 6, and 8 h. Compared to the sham group, i.p injection of sinomenine at a dose of 10 mg/kg had no distinct influence on PWT. However, i.p injection of sinomenine at a dose of 20 mg/kg alleviated the mechanical allodynia, beginning at 0.5 h and peaking at 2 h (Figure 1(b)). As shown in Figure 1(b), the maximum dose of sinomenine (40 mg/kg, i.p) significantly reversed the PWT, beginning at 0.25 h, peaking at 1 h, and lasting for 6 h. For chronic treatment, different doses of sinomenine or vehicle were administered i.p once daily from day 14 until day 18 and the PWTs were conducted 2 h after injection. As shown in Figure 1(c), continuous administration of sinomenine obviously and dose-dependently alleviated TCI-induced pain hypersensitivity, except for the 10 mg/kg sinomenine group. In addition, the PWTs of the 40 mg/kg sinomenine group were higher than the 20 mg/kg group.
Collectively, these results demonstrated that single and repeated sinomenine treatment attenuated the development and maintenance of CIBP, and the 40 mg/kg of sinomenine has a better analgesic effect.
Role of the JAK2/STAT3 pathway in CIBP
Emerging evidence has reported the involvement of the JAK2/STAT3 pathway in neuropathic pain. 48 To explore the expression and distribution of the JAK2/STAT3 pathway in the rat model of CIBP, we conducted Western blot analysis and immunohistochemistry experiments. The expression of spinal p-JAK2 and p-STAT3 was significantly increased in CIBP rats beginning and peaking on day 14 until the final examination on day 21 after TCI (Figure 2(a) and (b) and 3(a) and (b)). To investigate the cellular localization, we conducted double immunofluorescence staining of p-JAK2 with NeuN (neuronal marker), GFAP (astrocytic marker), or Iba1. As illustrated in Figure 2(c) to (k), p-JAK2 was only co-localized with microglia in the spinal cord, but not with the astrocyte or neuron.

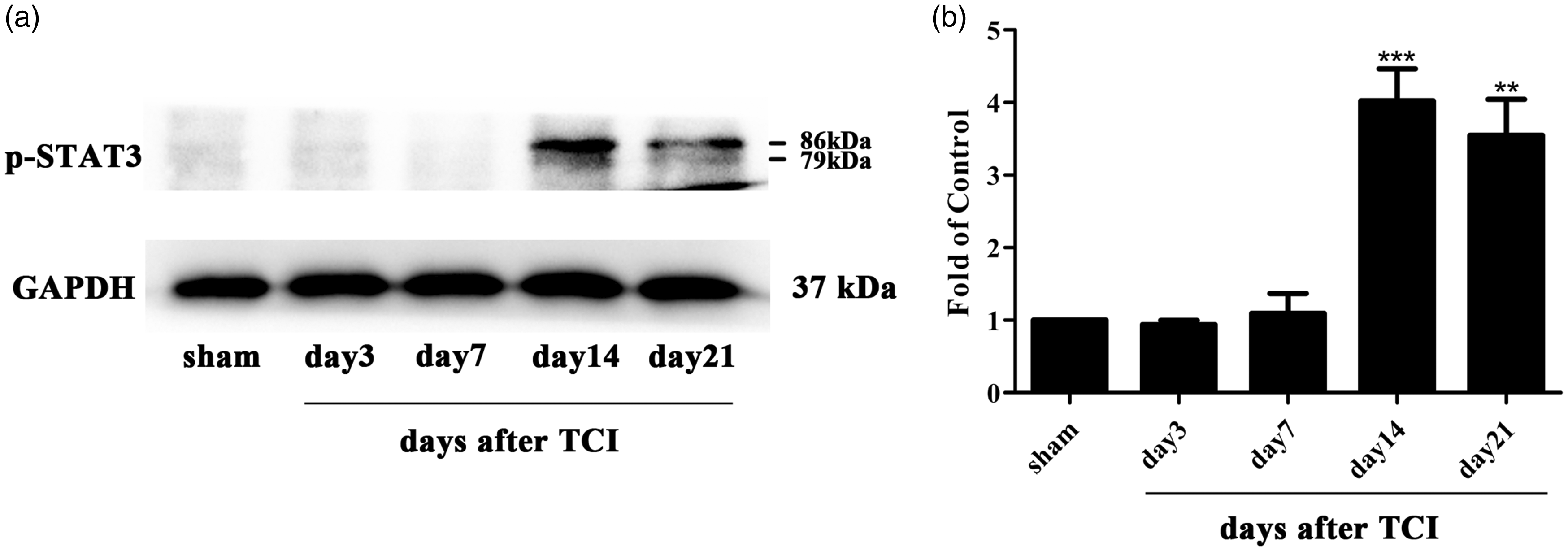
It turns out that the JAK2/STAT3 pathway was phosphorylated and up-regulated in the CIBP group, and p-JAK2 was only distributed in microglia but not with astrocytes or neurons.
Effects of repeated sinomenine on the CIBP-induced activation of JAK2/STAT3 pathway
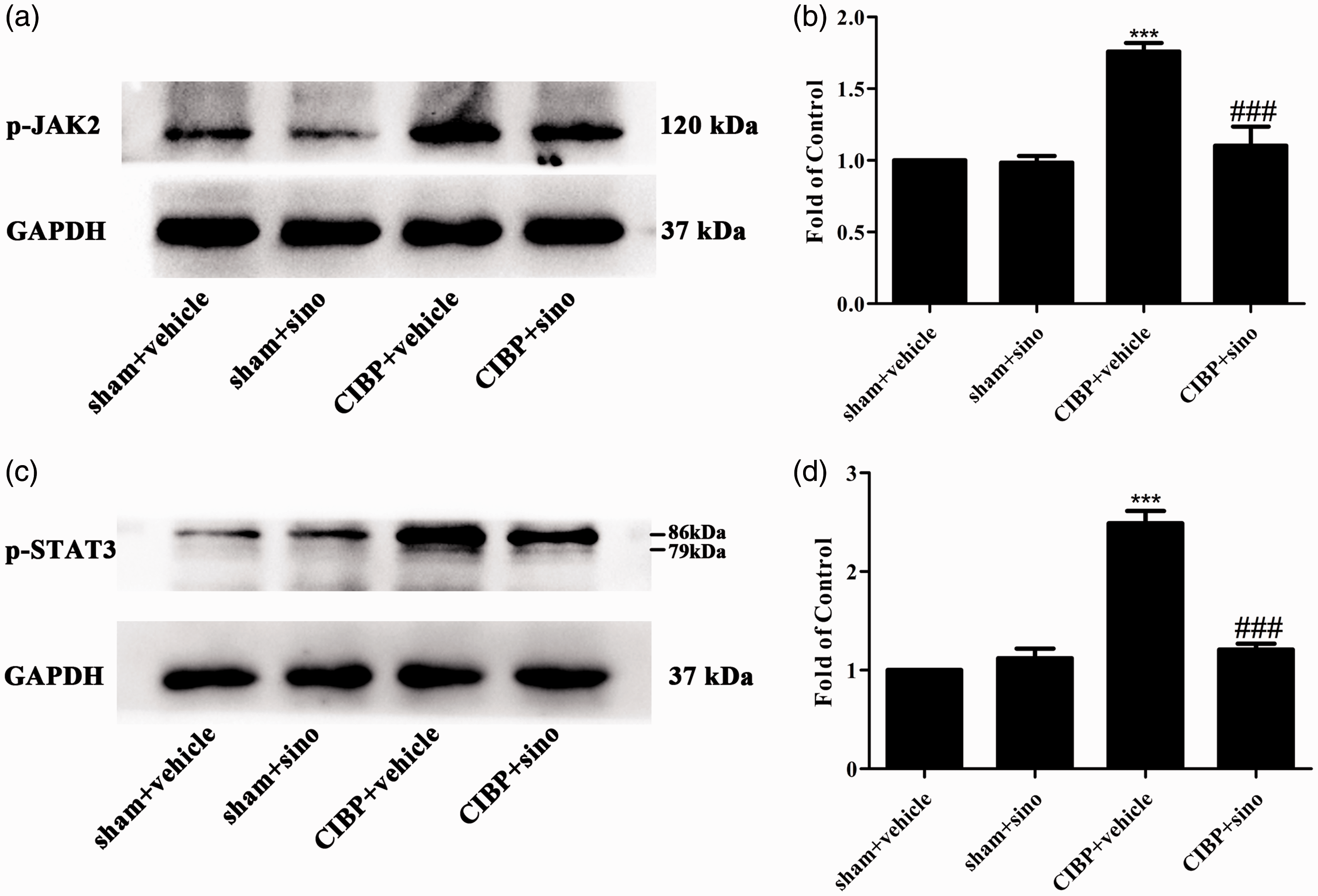
Sinomenine has been shown to alleviate neuroinflammatory injury via the CRYAB/STAT3 pathway after ischemic stroke. 49 To explore the specific mechanisms of sinomenine in attenuating TCI-induced pain, we conducted sinomenine (40 mg/kg, i.p) from day 14 to day 18 after TCI and the spinal cords were collected to detect the expression of cellular signaling pathways. As shown in Figure 4(a) to (d), the expression levels of p-JAK2 and p-STAT3 were reduced by sinomenine comparing with the CIBP + vehicle group.
Role of the CAMKII/CREB pathway in CIBP
Previous studies have suggested that the activation of the CAMKII/CREB pathway in the spinal cord contributes to neuropathic pain and cognitive impairment.50,51 The CAMKII/CREB signaling pathway participates in the intracellular signal transduction pathway which participates in cell proliferation, cell survival, and metabolism.52,53 The cerebral ischemia-induced injury was attenuated by paeoniflorin via modulation of the Ca2+/CAMKII/CREB signaling pathway. 54 In the CIBP group, the protein level of p-CAMKII increased on day 7 and peaked on day 14 (Figure 5(a) and (b)). Consistently, the protein change of p-CREB also raised on day 7, peaking on day 14 (Figure 6(a) and (b)). To investigate the cellular localization, we conducted double immunofluorescence staining of p-CREB with NeuN, GFAP, or Iba1. As shown in Figure 6(c) to (k), p-CREB was predominantly co-localized with neuron and a minority with microglia in the spinal cord.
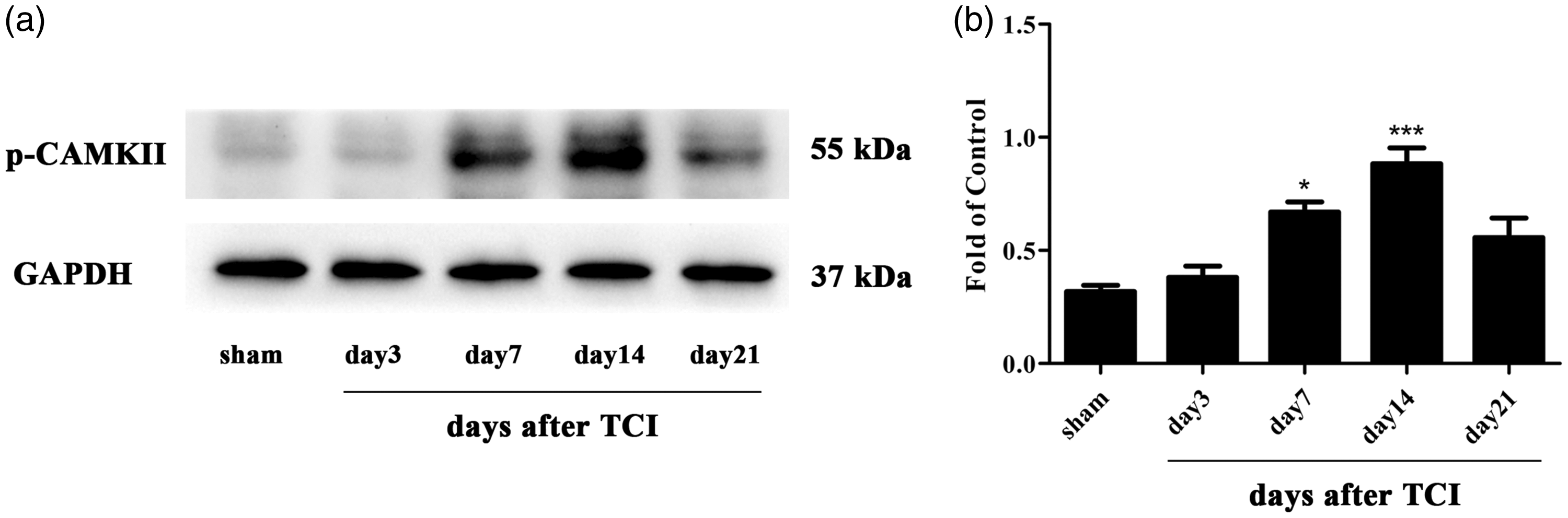

These results indicated that the CAMKII/CREB pathway was activated in the CIBP model and p-CREB was predominantly distributed in neuron.
Effects of repeated sinomenine on the CIBP-induced activation of CAMKII/CREB pathway
Sinomenine was reported to suppress inflammatory pain by down-regulating the CAMKII/CREB pathway. 19 In order to explore the effect of sinomenine on CIBP model, we conducted sinomenine (40 mg/kg, i.p) from day 14 to day 18. As illustrated in Figure 7(a) to (d), the protein expression of CAMKII/CREB was significantly decreased in the CIBP + sinomenine group.

Involvement of spinal microglia in the analgesic effect of sinomenine
In our previous investigations, the inhibition of microglia was proven to relieve cancer-induced allodynia and hyperalgesia.55–57 Expression level of Iba1 has been used as an indicator of microglial activity. As shown in Figure 8(a) and (b), the expression of Iba1 was significantly increased in CIBP rats beginning on day 14 until the final examination on day 21 after TCI. It was reported that sinomenine alleviates brain injury through inhibiting the activation of microglia. 20 In our study, we found that chronic treatment of sinomenine (40 mg/kg, once a day, i.p) suppressed the protein expression of microglia (Figure 8(c) and (d)). Furthermore, the immunofluorescence staining of Iba1 had demonstrated the results. As illustrated in Figure 8(e) to (i), microglia was distinctly activated in CIBP + vehicle group and the activation of Iba1 was significantly suppressed in CIBP + sinomenine group.

We have demonstrated that the JAK2 pathway was co-localized with microglia in this study. These results suggest that one of the mechanisms of sinomenine relieving TCI-related pain was down-regulating the JAK2/STAT3 pathway through inhibiting the activation of microglia. In addition, sinomenine may suppress the expression of the CAMKII/CREB pathway to weaken cancer-induced mechanical allodynia.
Discussion
In the current study, the principal findings were as follows: (1) acute and chronic intraperitoneal administration of sinomenine significantly alleviated TCI-induced mechanical allodynia; (2) transplanting of Walker 256 mammary gland carcinoma cells resulted in the activation of the JAK2/STAT3 pathway and p-JAK2 was only distributed in microglia; (3) the CAMKII/CREB signaling pathway in the spinal cord was activated in the rat model of CIBP; (4) chronic treatment of sinomenine distinctly suppressed the activation of microglia and significantly reduced the expression of both the JAK2/STAT3 pathway and the CAMKII/CREB pathway. To the best of our knowledge, we provide the first evidence that the JAK2/STAT3 signaling pathway is involved in CIBP. Moreover, we are the first to find that sinomenine can attenuate TCI-induced pain by inhibiting the microglial JAK2/STAT3 pathway and the CAMKII/CREB pathway.
CIBP-induced allodynia and hyperalgesia significantly compromise the quality of life in advanced cancer patients.58,59 In the CIBP rats, the activation of glia cells, including microglia and astrocyte, in the spinal segments is distinctly confirmed.60,61 Our previous studies showed that inhibition of microglia by minocycline significantly relieved mechanical allodynia in CIBP rats. 44 Sinomenine, a traditional Chinese medicine, has emerged as a potent inhibitor of microglial activation. Sinomenine was reported to prevent oligomeric Aβ-induced microglia activation and to show protection against Alzheimer’s disease. 62 In diabetic retinopathy, sinomenine reduced the release of TNF-α, IL-1β, and IL-6 in microglia via ROS production and NF-kB translocation. 63 In addition, a series of studies have proven the analgesic effect of sinomenine on chronic inflammatory pain and neuropathic.43,64 However, there is no evidence confirming the analgesic effect of sinomenine on CIBP. In this study, we found that sinomenine alleviated the PWT after TCI through inhibiting microglial activation.
STAT3, a member of the JAK/STAT signaling pathway, is phosphorylated and activated after CNS damage and participates in the microglial inflammatory response.65,66 Previous evidence suggested that JAK2 played a major role in STAT3 phosphorylation, and intrathecal injection of the JAK2 inhibitor AG490 markedly inhibits the accumulation of p-STAT3 in the dorsal spinal cord of spinal nerve ligation rats. 67 Furthermore, microglial JAK/STAT3 pathway activity directly impacted the characteristics of astrocytes and neurons, which participated in spinal cord plasticity and remodeling. 68 The JAK2/STAT3 pathway was shown to contribute to cell apoptosis in rats with transient global cerebral ischemia. 69 In rats with neuropathic pain and inflammatory pain, the JAK2/STAT3 signaling pathway was found to be up-regulated.34,70 Sinomenine was discovered to inhibit A549 human lung cancer cell invasion by mediating the STAT3 pathway. 71 In addition, sinomenine attenuated ischemic stroke-induced neuroinflammation by the CRYAB/STAT3 pathway. 49 Hence, we determined to prove whether inhibiting the microglial JAK2/STAT3 pathway was one of the mechanisms underlying the analgesic effect of sinomenine on CIBP. As shown in Figures 2(a) and (b) and 3(a) and (b), the expression of the JAK2/STAT3 pathway was significantly up-regulated in the rat model of CIBP, peaking on day 14 and lasting until day 21. The changes were the same as expression of microglia. Consistent with previous studies, our results showed that p-JAK2 was co-localized with microglia. Compared with the CIBP + vehicle rats, the activation of the JAK2/STST3 pathway was inhibited by the chronic treatment of sinomenine.
CAMKII was mainly distributed in the superficial laminae of the spinal cord dorsal horn, which played a vital role in transmission and processing of nociceptive signals.37,72,73 Autocamtide 2-related inhibitor peptide, a highly specific and potent inhibitor of CAMKII, was identified to reduce the protein level of p-CREB and showed analgesic effects. 74 Accumulating evidence has established that up-regulation of the CAMKII/CREB signaling pathway participates in models of pathological pain.40,41 As shown in Figures 5(a) and (b) and 6(a) and (b), our study demonstrated that the activation of the CAMKII/CREB pathway began on day 7 and peaked on day 14. Our study confirmed the role of the CAMKII/CREB signaling pathway in CIBP. In addition, our results showed that the expression of CAMKII/CREB was restored by chronic treatment of sinomenine.
Our investigation provides the first evidence that sinomenine alleviates CIBP by inhibiting microglial JAK2/STAT3 and neuronal CAMKII/CREB cascades. Sinomenine has significant analgesic effects against CIBP and may be considered as an alternative therapy for the clinical management of CIBP.
Footnotes
Authors’ contribution
Da-Wei Ye, Yu-Ke Tian, and Fang Luo designed the research study; Shu-Ping Chen, Ya-Qun Zhou, Jia Sun, Fei Cao, and Cody Braun performed the study; Shu-Ping Chen prepared the manuscript; all authors approved the final version of manuscript.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants from National Natural Science Foundation of P.R. China (Grant No. 81400917, 81371250, 81571053, and 81771196).