
Abstract
Phosphatidylinositol 4-phosphate 5-kinase type 1 gamma (Pip5k1c) generates phosphatidylinositol 4,5-bisphosphate, also known as PI(4,5)P2 or PIP2. Many pronociceptive signaling pathways and receptor tyrosine kinases signal via PIP2 hydrolysis. Previously, we found that pain signaling and pain sensitization were reduced in Pip5k1c+/− global heterozygous knockout mice. Here, we sought to evaluate the extent to which dorsal root ganglia selective deletion of Pip5k1c affected nociception in mice. Initially, we crossed sensory neuron-selective Advillin-Cre mice with a conditional Pip5k1c knockout (cKO) allele (Pip5k1cfl/fl). However, these mice displayed an early onset proprioceptive deficit. To bypass this early onset phenotype, we used two different tamoxifen-inducible Cre lines (Brn3a-Cre-ERT2 and Advillin-Cre-ERT2) to conditionally delete Pip5k1c in adults. Tamoxifen induced high efficiency deletion of PIP5K1C in dorsal root ganglia and slightly reduced PIP5K1C in spinal cord and brain in Brn3a-Cre-ERT2 × Pip5k1cfl/fl (Brn3a cKO) mice while PIP5K1C was selectively deleted in dorsal root ganglia with no changes in spinal cord or brain in Advillin-Cre-ERT2 × Pip5k1cfl/fl (Advil cKO) mice. Acute thermosensation and mechanosensation were not altered in either line relative to wild-type mice. However, thermal hypersensitivity and mechanical allodynia recovered more rapidly in Brn3a cKO mice, but not Advil cKO mice, following hind paw inflammation. These data collectively suggest that PIP5K1C regulates nociceptive sensitization in more regions of the nervous system than dorsal root ganglia alone.
Introduction
Nociceptive or physiological pain alerts an individual of potential threats such as injury or disease. However, acute pain can transform into pathophysiological stimulus-uncoupled chronic pain. Current pain management involves the use of opioids and nonsteroidal anti-inflammatory drugs, but chronic administration of these drugs have serious side effects, highlighting a major unmet medical need for new pain treatments.
Nociceptor sensitization contributes to chronic pain. 1 Pronociceptive factors in the inflammatory soup activate receptors such as G-protein-coupled receptors and receptor tyrosine kinases (RTKs). Downstream signaling cascades potentiate the activity and expression of a variety of ion channels and receptors, driving the increase in neuronal excitability. Inhibiting individual pronociceptive receptors and kinases have worked in animal models but have shown modest to no effects in humans.2–5 An alternative to these strategies is to target signaling molecules immediately downstream of multiple pronociceptive receptors.
Phosphatidylinositol 4,5-bisphosphate, also known as PI(4,5)P2 or PIP2, has important roles in cell signaling and is immediately downstream of many pronociceptive signaling pathways, despite only accounting for 0.5% to 1% of the phospholipid molecules in cells.6,7 Type 1 phosphotidylinositol 4-phosphate 5-kinases (PIP5KIs) synthesize PIP2 by phosphorylating the large pools of phosphatidylinositol 4-phosphate in cells. There are three Pip5k1 genes, two of which are ubiquitously expressed (Pip5k1a and Pip5k1b), while the third (phosphatidylinositol 4-phosphate 5-kinase type 1 gamma, Pip5k1c) is expressed predominantly in neuronal tissues.6,8,9 We previously found that global heterozygous deletion of Pip5k1c reduced PIP2 in dorsal root ganglia (DRG) and reduced pain signaling and inflammatory sensitization. 10 While these studies suggested that it might be possible to reduce pronociceptive signaling by selectively inhibiting a lipid kinase that generates PIP2 in DRG neurons, whether the decrease observed in pain behavior was due to a reduction in the enzyme expression specifically in sensory neurons or globally throughout the nervous system was unresolved. Here, we sought to evaluate the extent to which sensory neuron-selective deletion of Pip5k1c reduced acute and chronic pain sensitivity.
Materials and methods
Animals
All procedures involving vertebrate animals were approved by the Institutional Animal Care and Use Committee at the University of North Carolina at Chapel Hill. Mice were raised on a 12:12 h light:dark cycle, had ad libitum access to food and water, and were tested during the light phase. Estrous cycle was not monitored in females. Pip5k1cfl/fl mice 11 were crossed with Advillin-Cre mice, 12 Brn3a-Cre-ERT2, 13 and Advillin-Cre-ERT2 mice 14 to generate conditional knockouts. All mice were backcrossed to C57BL/6J mice for at least eight generations.
Injections and behavioral assays
Tamoxifen was prepared fresh daily by dissolving in 10% ethanol and 90% corn oil and sonicated for half an hour at room temperature. Intraperitoneal injections of 120 mg/kg tamoxifen were administered for seven consecutive days in mice aged six to eight weeks, and baseline behavioral testings were performed 10 days after the last injection. Mice were acclimated to the testing room, equipment, and experimenter one to three days before behavioral testing. Behavioral assays were performed as described previously. 15
Western blot analysis
Tissue lysates were prepared as described 10 from wild-type (WT) and conditional knockout (cKO) mice after completion of all behavioral testing. Briefly, DRG, spinal cord, and cerebral cortex were dissected and sonicated in radioimmunoprecipitation assay buffer. Protein concentration was determined via bicinchoninic acid assay, and 50 µg protein are loaded onto gels, which were subsequently transferred onto polyvinylidene difluoride membranes. Primary antibodies (1:4000 for PIP5K1C, generously provided by Hara et al. 16 and 1:3000 for β-actin; Abcam, AB6276; served as loading control) were prepared in Licor blocking buffer. Secondary antibodies (1:10,000; Li-Cor IRDye 680 and 800) were prepared in Licor blocking buffer. Blots were imaged on a LiCor gel imaging machine and analyzed using ImageJ. PIP5K1C band intensities were normalized to their respective β-actin controls.
Results
We first crossed Advillin-Cre (Advil-Cre) mice with Pip5k1c floxed (Pip5k1cfl/fl) mice to selectively delete Pip5k1c in the DRG.11,15,17 Advil-Cre is expressed in trigeminal ganglia by embryonic day 16.5 and in DRGs by postnatal day 1.12,17 Beginning around two months of age, we observed hind limb clasping and abnormal gait phenotypes in Advil-Cre × Pip5k1cfl/fl mice (data not shown), suggestive of an early-onset proprioceptive deficit.18–20 Hind limb clasping prevented us from probing the hind paw using standard nociceptive sensory assays.
To circumvent this early-onset proprioceptive phenotype, we crossed Brn3a-Cre-ERT2 mice with Pip5k1cfl/fl mice (henceforth called Brn3a cKO mice) to conditionally delete Pip5k1c in sensory neurons of adult mice.13,21 Tamoxifen administration greatly reduced PIP5K1C protein levels in the DRG of male and female Brn3a cKO mice (Figure 1). Non-Cre recombinase-expressing Pip5k1cfl/fl mice, also tamoxifen-treated, served as WT controls. Although Brn3a-Cre-ERT2 mice were previously used to delete genes in DRG,13,21 we observed a nonsignificant reduction of PIP5K1C in the spinal cord (male, p = 0.1; female, p = 0.14) and a small but significant reduction of PIP5K1C in the cerebral cortex (male, p < 0.05; female, p < 0.005).
Deletion of PIP5K1C in DRG and partial deletion in the cerebral cortex using Brn3a-Cre-ERT2 line. (a)–(c), PIP5K1C protein levels in the indicated tissues of male mice and (d) Western blot quantification relative to β-actin. Tissue harvested 18 to 20 days post tamoxifen injections. n = 10 WT and 14 cKO male mice. (e)–(g) PIP5K1C protein levels in the indicated tissues of female mice and (h) Western blot quantification relative to β-actin. Tissue harvested 18 to 20 days post tamoxifen injections. n = 14 WT and 15 cKO female mice. All data are mean ± SEM. Asterisks indicate significant difference between WT and cKO mice by t test. *p < 0.05, **p < 0.001, and ***p < 0.0005.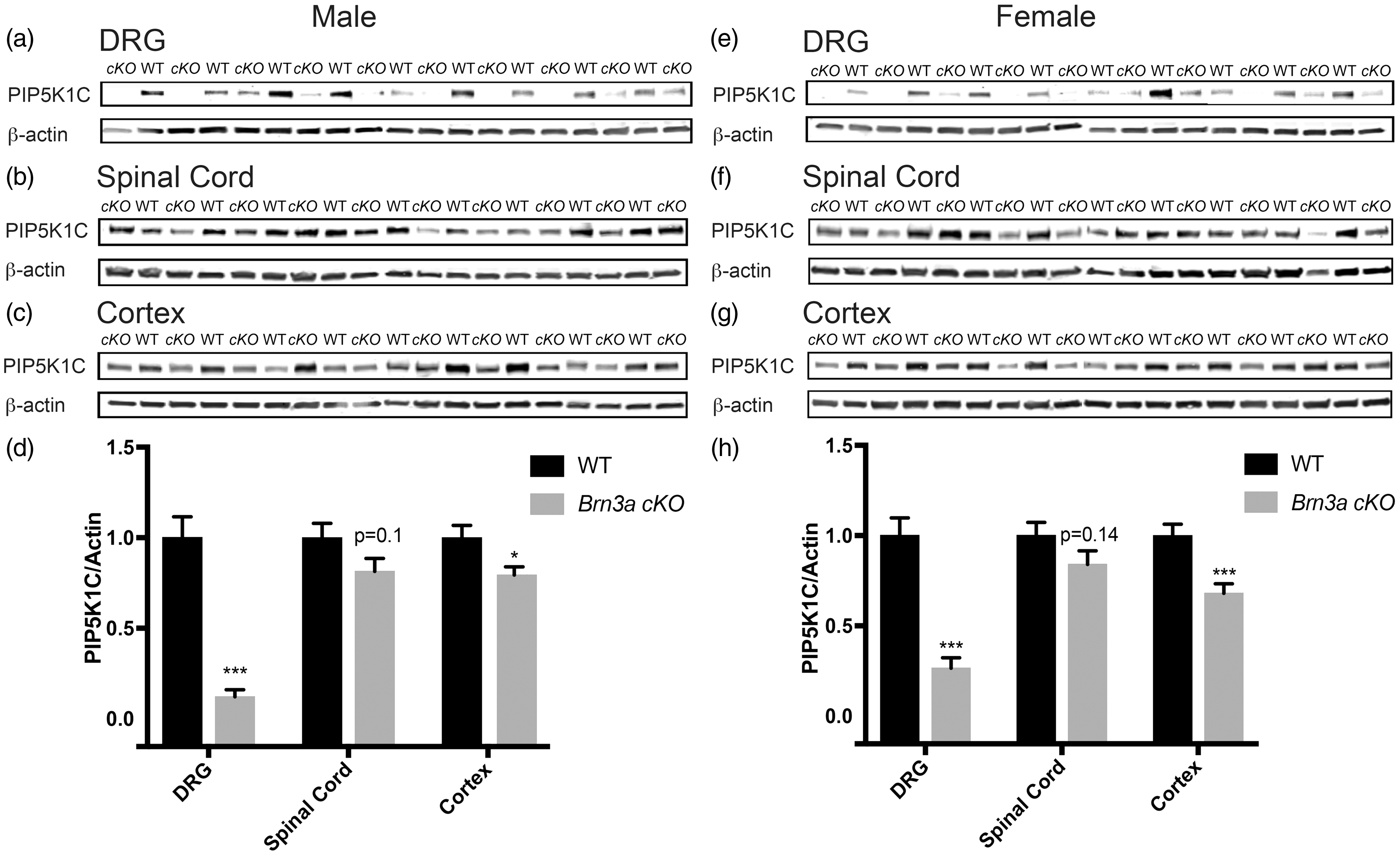
Noxious heat, mechanical, and cold behavioral assays with Brn3a cKO mice.

WT: wild type. Male and female Brn3a × Pip5k1c cKO mice display no significant changes in noxious thermal or mechanical sensitivity relative to WT mice, as determined by t test. n = 7 to 10 mice per group. All data are mean ± SEM.

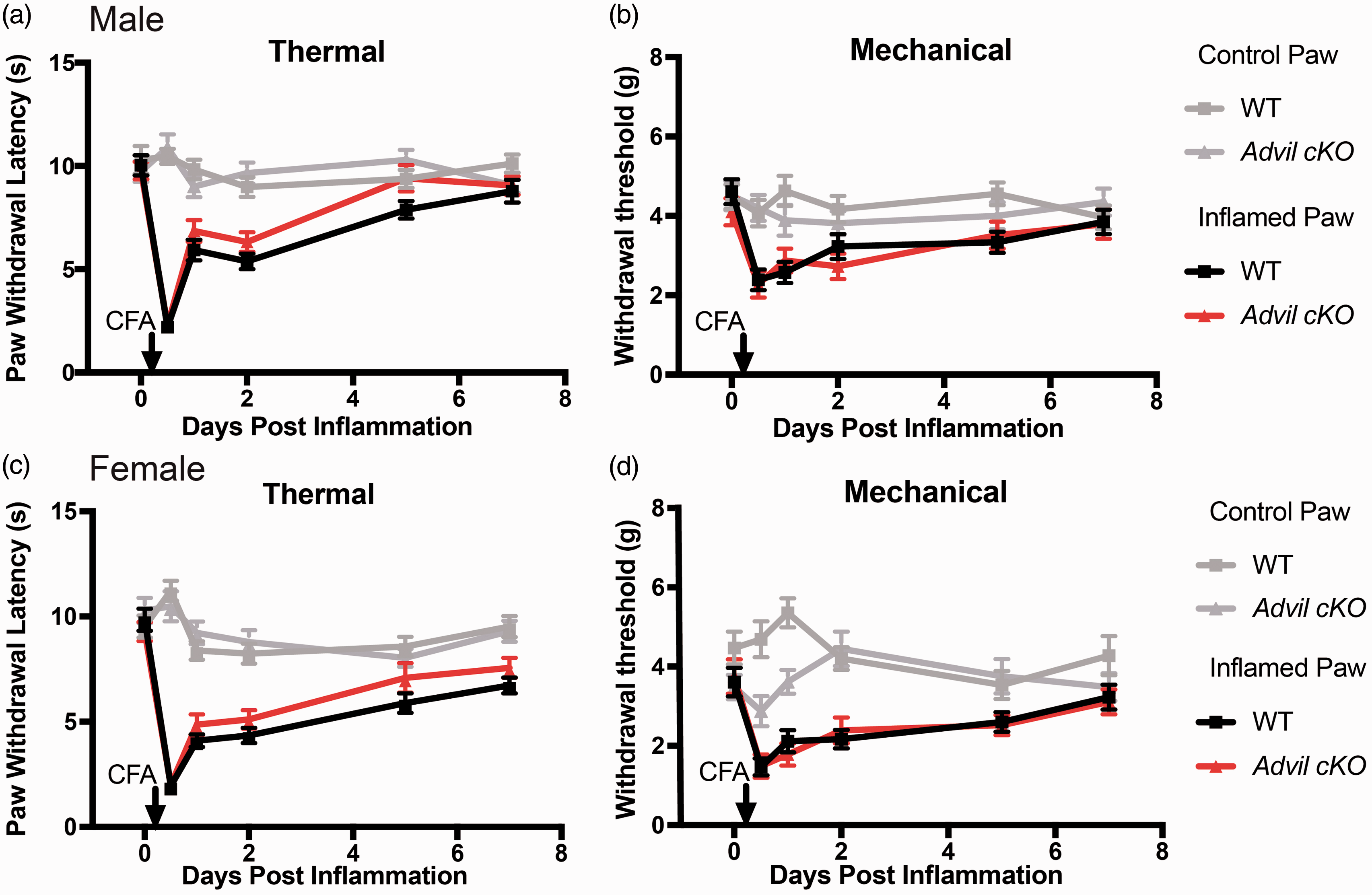
Early recovery of inflammation-induced thermal and mechanical hypersensitivity in Brn3a cKO mice. One hind paw of male (a and b) and female (c and d) mice was injected with the inflammatory agent CFA. The uninjected paw served as a control. (a) and (c) Thermal sensitivity measured before and several days after injection using the Hargreaves assay. (b) and (d) Mechanical sensitivity measured using an electronic von Frey apparatus. n = 10 WT and 14 cKO for males. n = 14 WT and 15 cKO for females. All data are mean ± SEM. Asterisks indicate significant difference between WT inflamed and cKO inflamed by two-way ANOVA, Dunnett’s post hoc test. *p < 0.05, **p < 0.005, and ***p < 0.0005.
Since tamoxifen-treated Brn3a cKO mice showed a small but significant reduction of PIP5K1C in cerebral cortex, we sought to more selectively delete Pip5k1c in sensory ganglia of adults. To accomplish this, we crossed Pip5k1cfl/fl mice with Advil-Cre-ERT2 mice (Advil cKO mice).
14
Tamoxifen administration significantly reduced PIP5K1C protein levels in the DRG of male and female Advil cKO mice but did not alter levels of PIP5K1C in spinal cord or cerebral cortex (Figure 3). The Advil cKO line thus more selectively deleted PIP5K1C in adult DRG than the Brn3a cKO line. No significant differences were observed between tamoxifen-treated WT mice and tamoxifen-treated Advil cKO mice in assays of noxious thermal or mechanical sensitivity (Table 2), nor were significant differences observed following CFA-inflammation (Figure 4). The discrepancy between Advil cKO and Brn3a cKO mice in the CFA inflammatory pain model could relate to less complete deletion of PIP5K1C in DRG of Advil cKO mice and/or partial deletion of PIP5K1C in cerebral cortex of Brn3a cKO mice (compare Figures 1 and 3).
Deletion of PIP5K1C in DRG but not in the spinal cord or cerebral cortex using Advil-Cre-ERT2 line. (a)–(c) PIP5K1C protein levels in the indicated tissues of male mice and (d) Western blot quantification relative to β-actin. Tissue harvested 18 to 20 days post tamoxifen injections. n = 13 WT and 13 cKO male mice. (e)–(g) PIP5K1C protein levels in the indicated tissues of female mice and (h) Western blot quantification relative to β-actin. Tissue harvested 18 to 20 days post tamoxifen injections. n = 12 WT and 9 cKO female mice. All data are mean ± SEM. Asterisks indicates significant difference between WT and cKO mice by t test. ***p < 0.0005. Noxious heat, mechanical, and cold behavioral assays with Advil cKO mice. WT: wild type. Male and female Advil cKO mice display no significant changes in noxious thermal or mechanical sensitivity relative to WT mice, as determined by t test. n = 9 to 13 mice per group. All data are mean ± SEM. Inflammation-induced thermal and mechanical hypersensitivity in Advil cKO mice is equivalent to WT mice. (a) and (b) One hind paw of male and (c) and (d) female mice was injected with the inflammatory agent CFA. The uninjected paw served as a control. (a) and (c) Thermal sensitivity measured before and several days after injection using the Hargreaves assay. (b) and (d) Mechanical sensitivity measured using an electronic von Frey apparatus. n = 13 WT and 13 cKO for males. n = 12 WT and 9 cKO for females. All data are mean ± SEM. No significant difference between WT inflamed and cKO inflamed by two-way ANOVA, Dunnett’s post hoc test.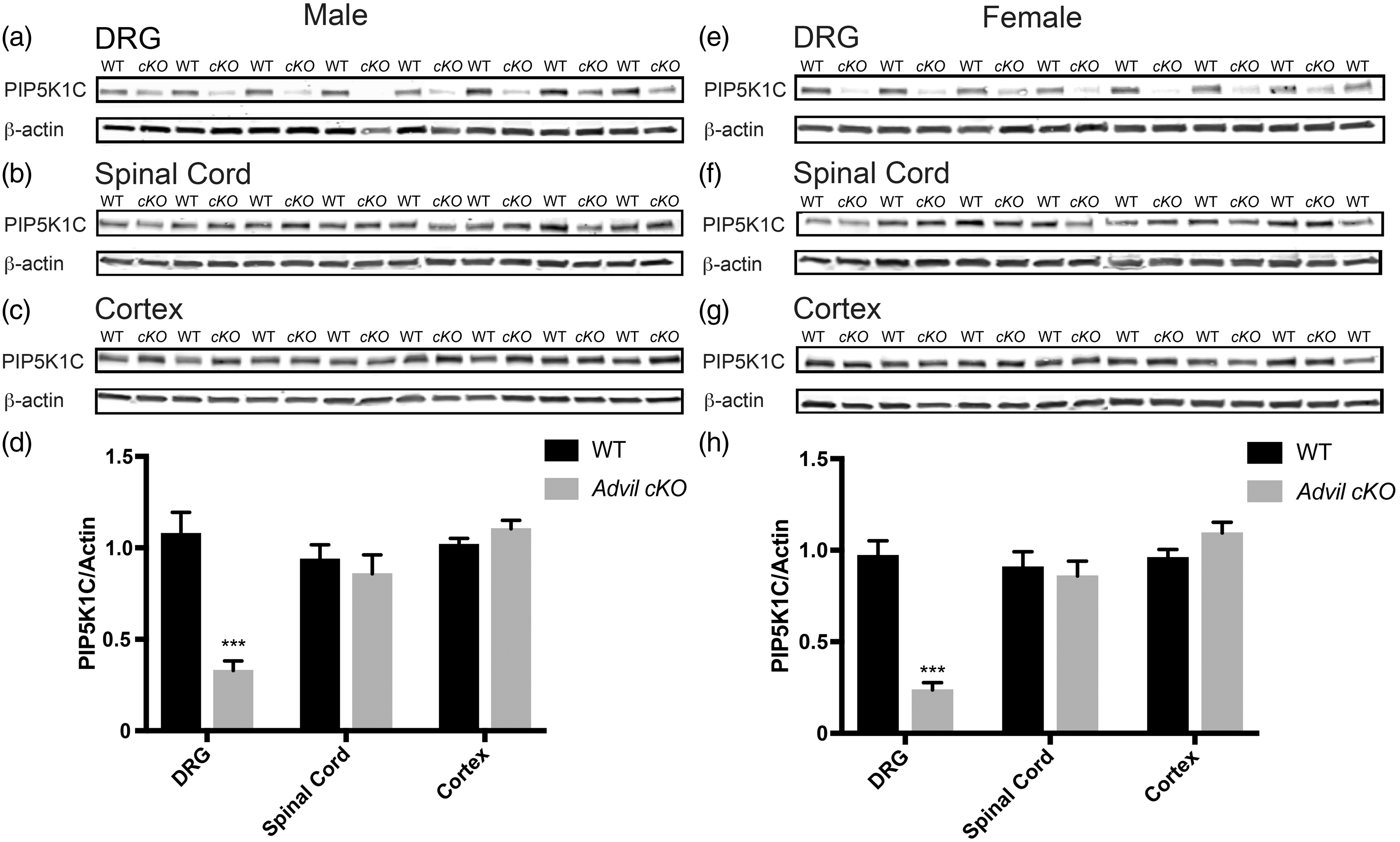

Discussion
Our study is the first to suggest that sensory-neuron selective loss of Pip5k1c causes an early-onset proprioceptive deficit in mice. Global loss of PIP5K1C in humans causes lethal congenital contractural syndrome type 3, 23 a syndrome featuring severe arthrogryposis (joint contractures) and death shortly before or immediately after birth. Joint contractures are seen in other human disorders that impair proprioception, 24 including biallelic loss of PIEZO2.25–27 The proprioceptive deficits that we observed in Advil-Cre × Pip5k1cfl/fl mice might relate to developmental loss of Pip5k1c in DRG, depletion of PIP2, and/or impaired neurotrophin-mediated neurite outgrowth. Neurotrophin-mediated RTK signaling regulates many aspects of neurodevelopment. 28 One of the kinases involved downstream of RTK signaling is phosphoinositide-3 kinase (PI3K). PI3K phosphorylates PIP2 to generate PIP3 to allow for the binding of Akt (protein kinase B). This leads to disinhibition of mTOR (mammalian target of Rapamycin), and this signaling axis regulates formation of axonal filopodia and dendritic arborization.29–31 Furthermore, PIP5K1C regulates adhesion junction formation and neuronal cell migration. 32 Future studies will be needed to resolve whether early loss of PIP5K1C impairs proprioceptive neuron development and/or maintenance.
The primary focus of our research was to evaluate how sensory neuron selective loss of PIP5K1C affects nociception. Thus, to bypass this early-onset proprioceptive phenotype, we conditionally deleted Pip5k1c in adult sensory ganglia using two different tamoxifen-inducible Cre lines. We generated tamoxifen-inducible Brn3a cKO and Advil cKO mice, neither of which displayed evidence of motor impairment or hind limb clasping over the course of our studies (data not shown).
We initially evaluated nociceptive phenotypes in tamoxifen-treated WT and Brn3a cKO mice. Brn3a cKO mice exhibited no difference in thermal or mechanical hypersensitivity following CFA inflammation but showed a faster recovery to baseline levels. We cannot conclude that this more rapid recovery was exclusively caused by the deletion of Pip5k1c in sensory neurons, as these Brn3a cKO mice also showed a significant reduction of PIP5K1C in the cerebral cortex. Initially, we tried a tamoxifen regiment (1 mg of intraperitoneal injections for five consecutive days) as reported in a previous Brn3a-Cre-ERT2 study, 21 but observed very low deletion efficiency (10%–50% deletion in DRG, data not shown). To further increase the deletion efficiency in DRG, we tried a higher tamoxifen dose (120 µg/g body weight, which is approximately 3 mg for a 25 g mouse, for seven consecutive days). However, this dosing schedule led to deletion of PIP5K1C in the spinal cord and cerebral cortex.
Brn3a is expressed in the dorsal horn of the spinal cord. 33 Additionally, conditional reporter gene targeting revealed Brn3a-positive neuronal projections from the superior colliculus into the cerebral cortex but no expression within the cerebral cortex.34,35 The slight decrease in PIP5K1C observed in our cortical Western blots may reflect low-level recombination in the cerebral cortex or, less likely, unintentional sampling of the superior colliculus, which is located just below the cerebral cortex. Interestingly, a LacZ reporter study showed laminar expression of Brn3a in the superior colliculus and mesencephalic central gray, also known as the periaqueductal gray (PAG). 35 The PAG is known to regulate ascending and descending pain pathways.36–38 It is therefore possible that PIP5K1C was also deleted in the PAG of Brn3a cKO mice. The more rapid recovery of Brn3a cKO mice following CFA-induced inflammation might thus relate to a loss of Pip5k1c in more regions of the nervous system than DRG alone. In support of this possibility, Advil cKO mice showed a more selective loss of PIP5K1C in DRG but did not show a more rapid recovery following CFA-induced inflammation. Homozygous Pip5k1c knockout mice (Pip5k1c−/−) showed major reduction of PIP2 (∼50%) in the brain, and Pip5k1c−/− cortical neurons have impaired synaptic transmission, although these animals die at birth. 39 It was thus not possible to study how global loss of Pip5k1c affected nociceptive behaviors in adults.
Collectively, our findings suggest that many of the nociceptive sensory phenotypes that we identified in Pip5k1c+/− mice are likely due to PIP5K1C haploinsufficiency in more regions of the body and nervous system than DRG alone. Support for this conclusion comes from our observation that Advil cKO mice show a >50% loss of PIP5K1C protein in DRG (which is greater than the ∼50% loss we reported in Pip5k1c+/− mice) but showed no sensory phenotypes. Moreover, Brn3a cKO mice showed faster recovery following CFA-induced inflammation, mirroring one of the phenotypes we identified in Pip5k1c+/− mice, but PIP5K1C was reduced in other regions of the nervous system besides DRG. Our study highlights the importance of evaluating multiple mutant lines (global, conditional, and adult-specific conditional) before drawing strong conclusions as to whether a gene regulates nociception exclusively via effects in sensory ganglia or via effects in other parts of the nervous system. Moreover, future work with additional Cre driver lines, such as spinal cord and/or brain specific, could be used to further validate the findings in this study.
Footnotes
Acknowledgments
The authors thank Brittany Schuck for her work in generating and characterizing the phenotypes of Advillin-Cre × Pip5k1cfl/fl mice, Hiroyuki Sakagami for providing PIP5K1C antibody, William Snider for providing the Brn3a-Cre-ERT2 mice, Ardem Patapoutian for providing the Advillin-Cre-ERT2 mice, and Eric McCoy for performing CFA injections.
Authors’ Contributions
LL performed the experiments. LL and MJZ designed the experiments, interpreted the data, and wrote the manuscript. Both authors read and approved the final manuscript.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by a grant to MJZ from NINDS (R01NS081127).