
Abstract
Background
Gastrin-releasing peptide (GRP) is thought to play a role in the itch evoked by intradermal injection of chloroquine. Although some early studies suggested that GRP was expressed in pruriceptive primary afferents, it is now thought that GRP in the spinal cord is derived mainly from a population of excitatory interneurons in lamina II, and it has been suggested that these are involved in the itch pathway. To test this hypothesis, we used the transcription factor Fos and phosphorylation of extracellular signal-regulated kinases (ERK) to look for evidence that interneurons expressing GRP were activated following intradermal injection of chloroquine into the calf, in mice that express enhanced green fluorescent protein (EGFP) in these cells.
Results
Injection of chloroquine resulted in numerous Fos- or phospho-ERK (pERK) positive cells in the somatotopically appropriate part of the superficial dorsal horn. The proportion of all neurons in this region that showed Fos or pERK was 18% and 21%, respectively. However, among the GRP–EGFP, only 7% were Fos-positive and 3% were pERK-positive. As such, GRP–EGFP cells were significantly less likely than other neurons to express Fos or to phosphorylate ERK.
Conclusions
Both expression of Fos and phosphorylation of ERK can be used to identify dorsal horn neurons activated by chloroquine injection. However, these results do not support the hypothesis that interneurons expressing GRP are critical components in the itch pathway.
Keywords
Background
Itch has been defined as an unpleasant sensation that causes a desire to scratch. Chronic itch (pruritus) can result from several dermatological and systemic diseases and represents a major unmet clinical need. Despite this, we still know relatively little about the neuronal circuits that are responsible for the sensation of itch.1–4 A major distinction can be made between those forms of itch that are relieved by antihistamines (histamine-dependent) and those that are not (histamine-independent). Many substances (pruritogens) can evoke itch, when injected into the skin, and these can operate through either histamine-dependent or histamine-independent mechanisms.5–7
An early insight into the peripheral and spinal pathways responsible for itch came from the observation that mice lacking the receptor for gastrin-releasing peptide (GRP) showed significantly reduced responses to certain pruritogens, but normal responses to a variety of painful stimuli. 8 Three lines of evidence suggested that GRP signalling at the spinal level was required for itch: (a) intrathecal administration of GRP receptor (GRPR) agonists evoked scratching, while antagonists reduced scratching in response to injected pruritogens; (b) in situ hybridisation revealed that the GRPR was expressed by neurons in lamina I of the dorsal horn; (c) a subsequent study from the same group showed that ablation of spinal GRPR-expressing neurons with saporin conjugated to bombesin (an amphibian homologue of GRP) resulted in reduced responsiveness to a variety of pruritogens. 9 This study also demonstrated that itch behaviours following administration of histamine-independent pruritogens such as chloroquine were substantially reduced in GRPR knockout mice, whereas histamine-dependent responses were much less affected. 9 Further evidence for a role of spinal GRP signalling in histamine-independent itch came from the finding that a GRPR antagonist delivered directly to the spinal cord significantly reduced the responses of superficial dorsal horn neurons to intradermal chloroquine but not to intradermal histamine. 10
There has been considerable debate concerning the source of GRP in the spinal cord. A number of studies have provided evidence that GRP is expressed by a specific subset of peptidergic primary afferents,8,11–13 and it has been suggested that the GRP released by these afferents acts through spinal GRPR to mediate itch. 8 However, several other groups have failed to detect GRP mRNA in primary afferents, using a variety of methods, including in situ hybridisation, RT and real-time polymerase chain reaction, and RNA seq.14–18 In addition, it has been reported that the antibodies that had been used to reveal GRP in the dorsal root ganglion8,13 can cross react with substance P,14,19 which is expressed by many peptidergic primary afferents.20,21
mRNA for GRP has been identified in the dorsal horn14–16,18,22,23 and the GRP-expressing cells can be identified in a mouse line (Tg-GRP-EGFP) in which enhanced green fluorescent protein (EGFP) is expressed under control of the GRP promoter.14,19,24 It has recently been shown that these cells represent a specific population of excitatory interneurons in lamina II.19,25 Taken together with the evidence against primary afferent expression of GRP, these findings have led to the alternative suggestion that GRP released by itch-activated spinal interneurons plays an important role in histamine-independent itch.14,24,26 To test this hypothesis, we have looked for evidence that chloroquine can activate GRP-expressing dorsal horn interneurons. As GRP cannot be detected in the cell bodies of these neurons with immunocytochemistry, 19 we used the Tg(GRP-EGFP) line. EGFP+ cells in this mouse are mainly present in lamina II, 19 and it has been shown that >90% of these possess GRP mRNA. 14 To reveal activated neurons, we stained for the transcription factor Fos, 27 which has been used in several previous studies,28–32 and for phosphorylated extracellular signal-regulated kinases (pERK).33,34
Methods
All animal experiments were approved by the Ethical Review Process Applications Panel of the University of Glasgow and were performed in accordance with the European Community directive 86/609/EC and the UK Animals (Scientific Procedures) Act 1986.
Fos induction
Six adult Tg(GRP-EGFP) mice of either sex (16–25 g; Gene Expression Nervous System Atlas [GENSAT]) were used to investigate Fos expression after intradermal injection of chloroquine or vehicle. The skin on the lateral aspect of the hindlimb was shaved on the day before stimulation, and in order to prevent scratching or biting of the injected area during the postinjection survival time (which would result in nociception-activated Fos), an Elizabethan collar (Harvard Apparatus, #72-0056) was applied at the time of shaving. Animals were briefly anaesthetised with isoflurane, and injections of either chloroquine (40 µg dissolved in 10 μl of phospate-buffered saline [PBS], n = 3 mice) or vehicle (10 μl PBS, n = 3 mice) were made into the lateral aspect of the left calf, after which the mice were allowed to recover from anaesthesia. The success of the intradermal injection was assessed by the formation of a small bleb6,7 in the calf skin. They were reanaesthetised with pentobarbitone (20 mg i.p.) and perfused through the left ventricle with fixative that contained 4% freshly depolymerised formaldehyde 2 h after the stimulus.
The L3 spinal segment (which contains the great majority of cells activated by these stimuli) was removed and postfixed for 2 h in the same fixative. The contralateral (right) side was notched to allow identification, and the tissue was cut into 60 µm thick transverse sections with a vibrating blade microtome (Leica VT1200). These were immersed in 50% ethanol for 30 min to enhance antibody penetration and then multiple-labelling immunofluorescence staining was performed as described previously.19,35 The sections were incubated for three days in the following combination of primary antibodies: mouse monoclonal antibody NeuN (Millipore; MAB377; diluted 1:500), chicken anti-EGFP (Abcam, ab13970; diluted 1:1000), and rabbit anti-Fos (Santa Cruz Biotechnology, sc-52; diluted 1:5,000). They were then incubated overnight in species-specific secondary antibodies that were raised in donkey and conjugated to Alexa 647, Alexa 488, or Rhodamine Red (Jackson Immunoresearch, West Grove, PA, USA). Secondary antibodies were diluted 1:500 (Alexa 647 and Alexa 488) or 1:100 (Rhodamine Red). All antibodies were diluted in PBS that contained 0.3% Triton X-100 and incubations were at 4°C. Following the immunocytochemical reaction, sections were stained with 4′,6-diamidino-2-phenylindole (DAPI) to reveal nuclei, mounted in antifade medium and stored at −20°C.
Sections were scanned with a Zeiss LSM 710 confocal microscope equipped with Argon multiline, 405 nm diode, 561 nm solid state, and 633 nm HeNe lasers. They were initially viewed with epifluorescence optics, and three sections from the chloroquine-injected mice that contained numerous Fos+ cells were selected from each animal, before EGFP was observed. Z-series (2 µm spacing) were then scanned through the full thickness of each section with the 40× oil-immersion lens (numerical aperture 1.3), with the confocal aperture set to 1 Airy unit. These scans included the central part of the dorsal horn, which contained the activated cells. The z-stacks from chloroquine-injected mice were analysed with Neurolucida for Confocal (MBF Bioscience, Williston, VT, USA). The outline of the grey matter was drawn, and the ventral border of the GRP plexus (which corresponds approximately to the boundary between the inner and outer parts of lamina II
19
) was determined from a maximum intensity projection and plotted onto the drawing. The mediolateral extent of the region that contained a high density of Fos+ cells was delineated by drawing two parallel lines that were orthogonal to the laminar boundaries (see Figure 1).
Fos in the dorsal horn following intradermal injection of chloroquine.
Initially, only the channels corresponding to NeuN and DAPI were viewed, and the locations of all neurons (NeuN+ cells) that lay within this region were plotted onto the drawing. To avoid overcounting cells that were split during sectioning, 36 we included cells if at least part of the nucleus (stained with DAPI) was present in the first optical section of the z-series and excluded them if part of the nucleus was present in the last optical section. 37 The channel corresponding to Fos was then viewed, and the presence or absence of staining in each of the neurons in the sample was recorded. Finally, the EGFP channel was viewed and all neurons that were EGFP+ were identified on the drawing. As Fos+ cells were present at highest density in lamina I and the outer part of lamina II (lamina IIo), we determined the proportion of all neurons that were located within this region and between the two parallel lines that were Fos-immunoreactive. We then determined the proportion of GFP+ neurons within this volume that were Fos-immunoreactive. Sections from the PBS-injected mice were also examined with the confocal microscope to test for the presence of Fos-immunoreactive neurons.
Phosphorylation of ERK
In initial studies, we performed intradermal injections of chloroquine or PBS and allowed a 5-min survival time before perfusion fixation, as phosphorylation of ERK occurs rapidly following stimulation.33–35,38 However, although we observed numerous pERK-positive cells in the superficial dorsal horn of the ipsilateral L3 segment in the chloroquine-injected mice, there were also many pERK+ cells in the corresponding region in PBS-injected mice. This is likely to have been caused by the mechanical noxious stimulus that results from needle insertion and distension of the skin during the intradermal injection. 6 In subsequent experiments, we therefore allowed a longer postoperative survival time (30 min). Six adult Tg(GRP-EGFP) mice of either sex (16–23 g; GENSAT) were anaesthetised with isoflurane and received intradermal injections of chloroquine (40 µg in 10 μl, n = 3 mice) or PBS (10 μl, n = 3 mice) into the left calf, which had been shaved the day before, as described earlier. The mice were maintained under isoflurane anaesthesia throughout the survival period. They then received pentobarbitone (20 mg i.p.) prior to perfusion fixation (a described earlier), which was carried out 30 min after the intradermal injection. The tissue was processed exactly as described for the Fos experiments, except that the Fos antibody was replaced with rabbit anti-pERK (Cell Signaling Technology, 9101; diluted 1:500). Confocal scanning and analysis were performed as described for the Fos experiments.
Characterisation of antibodies
The EGFP antibody was raised against recombinant full-length EGFP, and the distribution of staining matches that of native EGFP. The mouse monoclonal antibody NeuN was raised against cell nuclei extracted from mouse brain 39 and apparently labels all neurons but no glial cells in the rat spinal dorsal horn. 40 The Fos antibody was raised against a peptide corresponding to the N-terminus of human Fos. The pERK antibody detects p44 and p42 MAP kinase (Erk1 and Erk2) when these are phosphorylated either individually or dually at Thr202 and Tyr204 of Erk1 or Thr185 and Tyr187 of Erk2. It does not cross-react with the corresponding phosphorylated residues of JNK/SAPK or of p38 MAP kinase or with nonphosphorylated Erk1/2 (manufacturer’s specification). Specificity of both Fos and pERK antibodies was demonstrated by the lack of staining in nonstimulated areas (e.g. the contralateral dorsal horn).
Statistics
Data were formatted into 2 × 2 contingency tables for each animal, with rows corresponding to presence or absence of EGFP and columns to presence or absence of Fos or pERK. To determine whether there was a consistent difference in the proportions across the tables for the different cell types, we used the Mantel–Haenszel analysis. 41 Breslow–Day testing for homogeneity of the odds ratio was conducted prior to computation of the Mantel–Haenszel odds ratio and 95% confidence intervals.
Results
Fos
pERK and Fos following intradermal chloroquine.
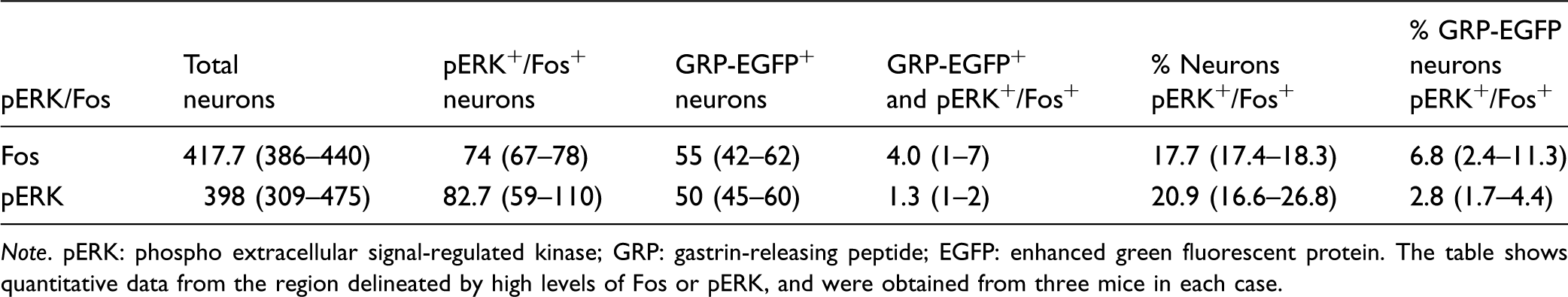
Note. pERK: phospho extracellular signal-regulated kinase; GRP: gastrin-releasing peptide; EGFP: enhanced green fluorescent protein. The table shows quantitative data from the region delineated by high levels of Fos or pERK, and were obtained from three mice in each case.
As reported previously,14,19,24,25 GRP-EGFP neurons were particularly numerous in lamina II, but were occasionally seen in lamina I, and were also scattered through the deeper parts of the dorsal horn (Figures 1c,e and 2). Although there was considerable overlap in the distribution of GRP-EGFP and Fos, relatively few of the GRP-EGFP cells were Fos-immunoreactive (Figure 1). The mean number of GRP-EGFP cells that were sampled in lamina I-IIo in the delineated area was 55, and of these, 7% were Fos-immunoreactive (Table 1).
pERK in the dorsal horn following intradermal injection of chloroquine.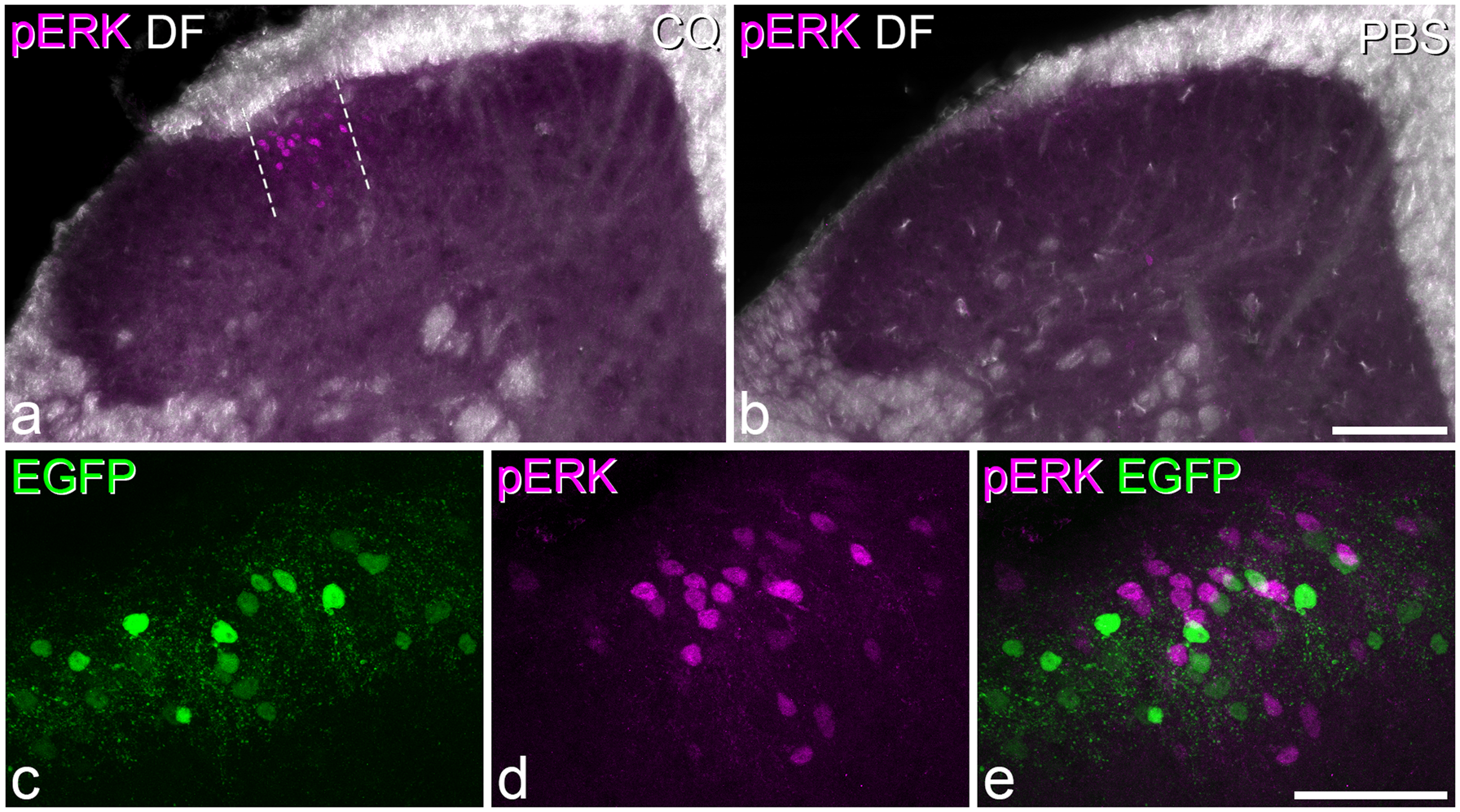
Analysis of odds ratios for coexpression of EGFP and activity-dependent markers.

pERK
Thirty minutes after chloroquine injection, numerous pERK-positive cells were seen in the ipsilateral L3 segment, while in contrast, very few pERK cells were seen in the mice that had received injection of PBS (Figure 2). The distribution of pERK cells in the chloroquine-injected mice was similar to that of Fos-positive neurons described earlier, forming a narrow mediolateral band in the middle part of the dorsal horn throughout the L3 segment. The total number of neurons in laminae I-IIo within the region delineated by pERK-immunoreactivity varied from 309 to 475 (n = 3 mice), and the proportion of these that were pERK-immunoreactive was 21% (Table 1). Again, although numerous GRP+ cells were present in this region (mean 50 cells), relatively few of these (3%) were pERK-immunoreactive (Figure 2c–e). The 95% confidence interval for the common odds ratio was below 1, indicating that GRP-EGFP cells were significantly less likely than other neurons to have phosphorylated ERK (Table 2).
Discussion
The main findings of this study are that following intradermal injection of 40 µg chloroquine into the calf, around 20% of neurons in laminae I-IIo in the somatotopically appropriate region of L3 upregulate Fos and phosphorylate ERK. However, although the activated cells showed an overlapping distribution with GRP-EGFP neurons, the latter were seldom Fos- or pERK-positive.
Fos and pERK as markers for itch activation
Several previous studies have used expression of Fos to identify neurons in the spinal dorsal horn that were activated by various pruritogens, including chloroquine, or in models of chronic itch.30,32,34,43–50 Between them, these studies have involved intradermal injections into several body regions: the cheek, neck, back, calf, and hindpaw. In each case, Fos+ neurons have been identified in laminae I–II of the corresponding spinal cord segments or spinal trigeminal nucleus. The distribution of Fos+ cells seen in the present study was therefore entirely consistent with these reports. As very few Fos cells were seen following an equivalent injection of the vehicle (PBS), it is highly likely that the Fos was induced as a result of the chloroquine and therefore represents the response to a pruritic stimulus.
There have apparently been very few reports of ERK phosphorylation in itch models. Zhang et al. 34 showed that histamine injected intradermally in the neck or hindpaw caused phosphorylation of ERK, which could be detected in the superficial dorsal horn, peaking 30 min after the stimulus. However, they found that intradermal injections of chloroquine that were sufficient to induce scratching did not evoke pERK. It is difficult to explain the discrepancy between their findings and those reported here, although in their study, the mice were not anaesthetised, and it is therefore possible that scratching of the affected area or activity in descending systems that are inactive during general anaesthesia suppressed ERK phosphorylation. It is unlikely that differences in the site of injection (neck in Zhang et al. and calf in the present study) were responsible for the different results, as Fos studies have shown very similar patterns of expression when pruritogens were injected into different sites. We found very little pERK in animals that had survived 30 min after PBS injection, which suggests that the phosphorylation was evoked by the chloroquine and therefore represents a pruritic response. However, our preliminary experiments with 5-min survival times indicated that the injection itself could cause significant phosphorylation of ERK. This was clearly very short lived, as it had completely subsided by 30 min.
Comparing our findings with Fos and pERK shows that there was a very similar distribution of labelled cells and that a comparable proportion of lamina I-IIo neurons within the somatotopically appropriate region was affected (∼20% in each case). Since ERK phosphorylation is an upstream regulator of Fos in the dorsal horn, 51 it is likely that the two markers were labelling equivalent populations of neurons, and the finding that GRP-EGFP cells were underrepresented with both markers is consistent with this suggestion.
Interestingly, Zhang et al. 34 reported that blocking phosphorylation of ERK with a MEK inhibitor reduced scratching behaviour in response to histamine but not to chloroquine. Our findings indicate that ERK is phosphorylated following intradermal injection of chloroquine but presumably it is not required for the resulting behaviour.
The identity of the itch-activated neurons in laminae I-IIo is not yet known, although it is likely that the great majority are interneurons, 28 and it will therefore be important for future studies to determine which neurochemical classes of interneuron show Fos or pERK following pruritogen injections. We have recently provided evidence that four nonoverlapping populations, defined by expression of GRP, neurotensin, neurokinin B, and substance P, can be identified among the excitatory interneurons in laminae I–III.19,25 However, the neurotensin and neurokinin B populations are both concentrated in the inner part of lamina II and lamina III and are therefore unlikely to be involved in itch. At present, the substance P-expressing cells are difficult to identify with immunocytochemistry due to the low level of peptide present in their cell bodies.
A role for GRP-expressing interneurons in chloroquine-evoked itch?
There is considerable evidence that the GRPR plays an important role in several forms of itch, 1 including that evoked by chloroquine.8,10 As it has been suggested that GRP-expressing dorsal horn interneurons are part of the itch pathway,14,24,26 we were surprised to find that GRP-EGFP cells were underrepresented among those that showed either Fos or pERK. It is likely that the dose of chloroquine we used was sufficient to cause itching, as similar doses have evoked scratching or biting in other studies in the mouse.52,53 In addition, we have found that intradermal injection of 40 µg of chloroquine into the calf in mice without Elizabethan collars leads to biting of the injected area (EP unpublished data), and this dose was sufficient to evoke Fos or pERK in a substantial proportion (∼20%) of the neurons in laminae I–IIo in this study.
This leaves several possible explanations for this paradox. (1) A significant number of GRP-EGFP cells may have been activated without expressing Fos or phosphorylating ERK, and the risk of false-negative results should always be considered with studies involving these activity-dependent markers. It is also possible that a relatively low level of activation is required to induce release of neuropeptides, including GRP. (2) It may be that sufficient GRP is released from the relatively few GRP-EGFP cells that were activated. (3) Solorzano et al. 14 reported that only ∼70% of neurons with GRP mRNA were EGFP+ in this mouse line and therefore EGFP-negative GRP-expressing neurons may have been activated. However, this explanation would require that there was a specific subpopulation of GRP-expressing neurons that lacked EGFP and that these were selectively activated by chloroquine.
Alternatively, the GRP-expressing dorsal horn interneurons may not be critically involved in itch pathways. There continues to be considerable debate about whether GRP is released from primary afferents,12,24 and if it is then this would be consistent with our findings. However, the majority view now appears to be against primary afferents as a source of GRP,15–18 at least in naïve animals. 14
Finally, it is possible that the GRPR on lamina I neurons is activated by a different peptide. Although the other main bombesin-like peptide, neuromedin B, is expressed in primary afferents,15–18 it has a very low affinity for the GRPR 54 and is therefore unlikely to mediate its activation following pruritic stimulation. However, there may be another, as yet unknown, peptide that forms the link between pruriceptive primary afferents and the GRPRs that are expressed by lamina I neurons, and the fact that there is an orphan receptor (BB3) would be consistent with this suggestion. 54
Conclusions
The results of this study show that both Fos and pERK can be used to identify cells that have been activated by intradermal injections of the pruritogen chloroquine and that similar numbers of cells are labelled for each marker. However, they do not support the suggestion that GRP-expressing interneurons in the superficial dorsal horn are preferentially activated by chloroquine.
Footnotes
Authors’ contributions
AMB, MGM, and AJT designed the study. AMB, MGM, and EP performed the anatomical experiments and analysed the resulting data. All authors contributed to the writing of the manuscript and approved the final version.
Acknowledgements
The authors are grateful to Mr R Kerr and Ms C Watt for excellent technical assistance and to Dr R Ganley for helpful discussion. The mouse strain Tg(GRP-EGFP) was originally obtained from the Mutant Mouse Regional Resource Centre (MMRRC), a National Centre for Research Resources- and NIH-funded strain repository and was donated to the MMRRC by the NINDS-funded GENSAT BAC transgenic project.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The work was supported by the Wellcome Trust (grant 102645) and the MRC (grant L003430).