
Abstract
Background
Chronic pain associated with inflammation is an important clinical problem, and the underlying mechanisms remain poorly understood. 4-Nitrophenylphosphatase domain and nonneuronal SNAP25-like protein homolog (NIPSNAP) 1, an interacting protein with neuropeptide nocistatin, is implicated in the inhibition of tactile pain allodynia. Although nocistatin inhibits some inflammatory pain responses, whether NIPSNAP1 affects inflammatory pain appears to be unclear. Here, we examined the nociceptive behavioral response of NIPSNAP1-deficient mice and the expression of NIPSNAP1 following peripheral inflammation to determine the contribution of NIPSNAP1 to inflammatory pain.
Results
Nociceptive behavioral response increased in phase II of the formalin test, particularly during the later stage (26–50 min) in NIPSNAP1-deficient mice, although the response during phase I (0–15 min) was not significantly different between the deficient and wild-type mice. Moreover, phosphorylation of extracellular signal-related kinase was enhanced in the spinal dorsal horn of the deficient mice. The prolonged inflammatory pain induced by carrageenan and complete Freund’s adjuvant was exacerbated in NIPSNAP1-deficient mice. NIPSNAP1 mRNA was expressed in small- and medium-sized neurons of the dorsal root ganglion and motor neurons of the spinal cord. In the formalin test, NIPSNAP1 mRNA was slightly increased in dorsal root ganglion but not in the spinal cord. In contrast, NIPSNAP1 mRNA levels in dorsal root ganglion were significantly decreased during 24–48 h after carrageenan injection. Prostaglandin E2, a major mediator of inflammation, stimulated NIPSNAP1 mRNA expression via the cAMP-protein kinase A signaling pathway in isolated dorsal root ganglion cells.
Conclusions
These results suggest that changes in NIPSNAP1 expression may contribute to the pathogenesis of inflammatory pain.
Background
4-Nitrophenylphosphatase domain and nonneuronal SNAP25-like protein homolog (NIPSNAP) 1 is highly expressed in the brain, spinal cord, kidney, and liver.1–3 NIPSNAP1 is exclusively expressed in neurons 2 and is localized in the postsynaptic density fraction of synapses. 4 NIPSNAP1 is associated with several neuronal diseases, including seizures, 4 prepulse inhibition, 5 phenylketonuria, 6 and Alzheimer disease. 1 Furthermore, we recently reported that NIPSNAP1 interacts with the neuropeptide nocistatin (NST) in vitro and implicates in the inhibition of tactile pain allodynia by NST in vivo. 3 NST, which is produced from the same precursor as the opioid peptide nociceptin/orphanin FQ (N/OFQ), blocks tactile allodynia evoked by N/OFQ. 7 The inhibitory effect of NST on the N/OFQ-induced allodynia is abolished in NIPSNAP1-deficient (NIPSNAP1−/−) mice. 3 Moreover, NST inhibits some inflammatory pain responses.8–12 Intraplantar administration of nmol level of NST reduces pain responses induced by peripheral injection of carrageenan. 9 Intrathecal (i.t.) administration of NST at low doses (fmol–pmol range) blocks the pain behavior induced by peripheral injection of formalin and carrageenan.10–12 Intracerebroventricular administration of pmol level of NST reduces inflammatory hyperalgesia induced by carrageenan. 8 However, whether NIPSNAP1 mediates inflammatory pain is unknown.
In the present study, we used NIPSNAP1−/− mice to determine the role of NIPSNAP1 in mouse models of inflammatory pain induced by formalin, carrageenan, and complete Freund’s adjuvant (CFA). We also analyzed the changes of NIPSNAP1 mRNA expression in dorsal root ganglion (DRG) and spinal cord following peripheral inflammation and the regulation of NIPSNAP1 mRNA expression induced by a major mediator of inflammation prostaglandin (PG) E2.
Methods
Animals
NIPSNAP1−/− mice (Accession No. CDB0535K: http://www.clst.riken.jp/arg/mutant%20mice%20list.html) were generated as previously reported. 3 All experiments were conducted in accordance with the ethical guidelines of the Osaka Institute of Technology Animal Research Committee and Kansai Medical University Animal Research Committee. NIPSNAP1−/− mice and their cognate wild-type mice were generated by breeding the respective heterozygous pairs. Eight- to 12-week-old male mice were used for behavioral analyses. Animal experiments were conducted in accordance with the National Institutes of Health guide for the care and use of laboratory animals and the ethical guideline of the Ethics Committee of the International Association for the Study of Pain.
Chemicals
PGE2 and selective PGE2 receptor subtype agonists, ONO-DI-004 (EP1 agonist), ONO-AE1-259 (EP2 agonist), ONO-AE-248 (EP3 agonist), and ONO-AE1-329 (EP4 agonist), were kindly supplied by Ono Pharmaceutical Co, Ltd. We purchased H-89 from Tocris Bioscience. Aspirin (Wako Pure Chemical Industries, Ltd.) was suspended in 0.5% methylcellulose solution (vehicle).
Pain behavioral analysis
Thermal nociception was assessed as the hindpaw withdrawal latency from a radiant heat stimulus determined by using a plantar test. 13 Noxious thermal nociception was examined in the hot plate test at 50℃ and 55℃.7,13 The cutoff time was 30 s to prevent tissue damage. For immunohistochemistry, heat stimulus was produced by immersion of the hindpaw into a water bath for 1 min (repeat six times of 10-s immersion and 10-s off interval). Mechanical allodynia was assessed by the first response method 13 and the up-down method using von Frey filaments. 14 In the first response method, calibrated von Frey filaments were applied in an ascending order through the mesh floor to the plantar surface of the hind paw five times at intervals of a few seconds. The withdrawal threshold was defined as the lowest force required to elicit a withdrawal reflex of the paw. In the up-down method, the mechanical sensitivity was evaluated using a set of eight von Frey filaments (0.04, 0.07, 0.16, 0.4, 0.6, 1.0, 1.4, and 2.0 g). The first stimulus was always the 0.4 g filament. When a withdrawal reflex of the paw was elicited, the next lower rated filament was applied, and when there was no response, the next higher rated filament was used. After the first change in direction of responses, additional four measurements were carried out, and the 50% withdrawal threshold was calculated. 14 No response to stimulus with the 2.0 g filament assigned the value of 2.0 g.
To analyze inflammatory pain, 20 µl of 1% carrageenan (Sigma-Aldrich) or CFA (Sigma-Aldrich) was subcutaneously injected into the right-dorsal surface of the hind paw. The paw diameter was measured using a micrometer caliper (Shinwa). Mechanical allodynia was assessed by the first response method using von Frey filaments.
The formalin test was essentially performed according to the procedure of Nakano et al. 11 Mice were acclimatized to individual observation cages for 30 min and were then subcutaneously injected with 20 µl of 2% formalin in 0.9% saline into the dorsal surface of the right hind paw. The time mice were engaged in licking and biting behaviors was then recorded at 5 min intervals for 50 min. The acute phase (approximately 15 min after formalin injection) was defined as phase I (0–15 min post injection). The persistent inflammatory phase, which increased after 15 min and persisted up to 50 min, comprised an early phase II (16–25 min postinjection) and a late phase II (26–50 min postinjection).
Immunohistochemistry
Immunohistochemistry was essentially performed as previously described. 7 Briefly, mice were deeply anesthetized using sodium pentobarbital and then intracardially perfused with phosphate-buffered saline (PBS), followed by 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4). The lumber spinal cord was dissected, fixed again in 4% PFA overnight, and cryoprotected in 30% sucrose overnight. Tissues sections (40 -µm thick) were prepared using a sliding microtome. The sections were blocked with PBS containing 10% normal goat serum and 0.2% Triton X-100 and then incubated with the primary antibody overnight at 4℃. The primary antibodies were rabbit anti-phospho-ERK (1:1000, Cell Signaling Technology), rat anti-F4/80 (1:250, AbD Serotec), and rat anti-Gr-1 (1:250, R&D systems). Immune complexes were visualized using Alexa546-labeled secondary antibodies (1:500, Invitrogen). Digital images were captured using a Nikon A1 laser-scanning confocal microscope equipped with an argon HeNe1 laser and appropriate filter. Control and experimental tissues were concurrently analyzed, and the images were captured using the same conditions.
Measurement of MCP-1
The hindpaws were homogenized with a Polytron homogenizer with ice-cold PBS containing 0.1% Triton X-100 and protease inhibitor (Roche). The homogenate was centrifuged at 20,000 g for 15 min at 4℃, and the resulting supernatant was subjected to mouse MCP-1 ELISA kit (BioLegend) according to the manufacturer’s protocol. Protein concentration of the supernatant was quantified by using a DC protein assay (Bio-Rad).
DRG cells
DRG cells were dissected from ddY mice at four to eight weeks of age. DRGs were digested with 0.7 mg/ml collagenase type II (Sigma-Aldrich) in minimum essential medium (MEM) for 50 min at 37℃ followed by incubation in 0.01% trypsin-EDTA (Life Technologies) for 5 min at 37℃. After the addition of 0.2 mg/ml of trypsin inhibitor (Sigma-Aldrich), the cells were dispersed by pipetting and then filtered through a Falcon 70 -µm cell strainer. The cells were plated onto poly-
Reverse-transcription polymerase chain reaction and real-time polymerase chain reaction
Total RNAs were extracted from mouse tissues using TRIzol reagent (Life Technologies) and from the DRG cells using ReliaPrep RNA cell miniprep system (Promega), and reverse transcribed. First-strand cDNA was amplified in a buffer containing Gene Taq DNA polymerase (Nippon Gene) and an anti-Taq antibody (anti-Taq high; Toyobo) with primers as follows: 5′-AACATGGCTCCGCGGTTGTGC-3′ and 5′-AGTCCATGAGGGCCGGGTAGC-3′ for NIPSNAP1; 5′-AATGTGTCCGTCGTGGATCTG-3′ and 5′-TGGTCCAGGGTTTCTTACTCC-3′ for glyceraldehyde-3-phosphate dehydrogenase (GAPDH); 5′-GGAGCCAAGTTCCAGGCCAT-3′ and 5′-TGTGCCATTATCGCCTGTTG-3′ for EP1; 5′-GACTTCGATGGCAGAGGAGA-3′ and 5′-AGGACTGAGCGCATTAACCT-3′ for EP2; 5′-CGGTTGAGCAATGCAAGACA-3′ and 5′-CAAGGAGGTGGAGCTGGAAG-3′ for EP3α; 5′-GGCAGAATCACCACGGAGAC-3′ and 5′-CTCAGTCATCCTTCCCTGGG-3′ for EP3β; 5′-AGATGTCGGTTGAGCAATGC-3′ and 5′-TGCCCTTTCTGTCCATCACT-3′ for EP3γ; and 5′-TCTCTGGTGGTGGTGCTCATCTG-3′ and 5′-TGTAAATCCAGGGGTCCAGG-3′ for EP4). Reverse-transcription polymerase chain reaction (RT-PCR) amplifications of NIPSNAP1 and EPs were performed as follows: 1 cycle at 94℃ for 1 min followed by 10 cycles at 94℃ for 1 min and at 72℃ for 3 min, 15 cycles at 94℃ for 1 min, 65℃ for 2 min, and at 72℃ for 30 s, and 10 cycles at 94℃ for 1 min, 62℃ for 2 min, and at 72℃ for 30 s, and 1 cycle at 72℃ for 2 min. RT-PCR amplification of GAPDH was performed as follows: 1 cycle at 94℃ for 1 min followed by 35 cycles at 94℃ for 1 min, 60℃ for 1.5 min, and at 72℃ for 30 s, and finally 1 cycle at 72℃ for 7 min. mRNA levels were measured using quantitative real-time PCR with a StepOne System (Applied Biosystem) and GoTaq qPCR master mix (Promega). The real-time PCR conditions were as follows: 1 cycle at 95℃ for 2 min followed by 40 cycles at 95℃ for 15 s and at 60℃ for 1 min. mRNA levels were normalized to those of GAPDH.
In situ hybridization
Sense or antisense NIPSNAP1 riboprobe RNAs were synthesized in vitro using digoxigenin-11-UTP (Roche) with a 700-bp coding region of NIPSNAP1 as template. A sliding microtome was used to prepare spinal cord sections (40 µm) from paraformaldehyde-perfused mice. DRG sections (10 µm) prepared using a cryostat were mounted on APS-coated glass slides (Matsunami). The free-floating spinal cord and DRG tissue sections on glass slides were hybridized overnight with digoxigenin-labeled riboprobes at 65℃ in a solution containing 50% formamide, 4% dextran sulfate, 250 µg/ml salmon sperm DNA, 250 µg/ml yeast tRNA, 1× Denhardt’s solution, 0.2% SDS, 25 mM EDTA, 750 mM NaCl, and 25 mM PIPES. Hybridized sections were then washed with 0.1× SCC after treatment with 20 µg/ml RNase A for 30 min at 37℃. The sections were then reacted with an alkaline phosphate-conjugated anti-digoxigenin antibody, and hybrids were detected using 2-hydroxy-3-naphtoic acid-2′-phenylanilide phosphate (Roche), 4-chloro-2-metylbenzenediazonium hemi-zinc chloride salt (Fast Red TR, Roche), and levamisole (Sigma-Aldrich). Thereafter, the free-floating spinal cord was mounted on glass slides. Digital images were captured using an LSM510 Zeiss laser-scanning confocal microscope equipped with an argon He–Ne laser and the appropriate filter.
Statistical analysis
Data are expressed as the mean ± SEM. We evaluated the statistical significance of the differences between mean values using the Student t-test or the Mann-Whitney U-test.
Results
Physiological pain in NIPSNAP1−/− mice
To address whether NIPSNAP1 deficiency affects the behavioral characteristics of physiological pain, we subjected NIPSNAP1−/− and wild-type mice to thermal and mechanical stimuli. When NIPSNAP1−/− mice were assessed on the hotplate at 50℃ and 55℃, these mice showed no significant difference in thermal nociception assessed by withdrawal latency (Figure 1(a)). In the Hargreaves test, NIPSNAP1−/− mice responded to the noxious thermal stimulus with latency comparable to that of wild-type mice (Figure 1(b)). We next carried out von Frey filament tests using two methods as follows: the first response method and the up-down method. NIPSNAP1−/− mice demonstrated a slight, but not statistically significant decrease in the threshold of nociceptive responses, in as assessed either by the lowest force required to elicit a withdrawal reflex of the first response method (Figure 1(c)) or by the 50% withdrawal threshold of the up-down method (Figure 1(d)). In addition, extracellular signal-regulated kinase (ERK) activation in the spinal dorsal horn is involved in generating pain hypersensitivity.15,16 Phosphorylation of ERK (p-ERK) in spinal dorsal horn were induced by heat stimulus at 55℃ for 1 min (Figure 1(e)) but not by mechanical stimulus at 4 g for 1 min (data not shown). There was no difference in the heat-stimulus induced p-ERK positive cell number between wild-type and NIPSNAP1−/− mice (Figure 1(f)). These results suggest that NIPSNAP1−/− mice showed normal pain sensitivity to thermal and mechanical stimuli.
Effects of NIPSNAP1 deficiency on pain sensitivity. Response latencies of wild-type (+/+) and NIPSNAP1−/− (−/−) mice on the hot plate at 50℃ and 55℃ (a) and their response to a radiant heat stimulus (b). Data are expressed as the mean ± SEM (n = 9–12). The paw threshold of wild-type and NIPSNAP1−/− mice by the first response method (c) and the 50% withdrawal threshold by the up-down method (d) induced by mechanical stimuli using the von Frey filaments (n = 7). Heat stimulus (55℃, 1 min) induced ERK phosphorylation (red) in the spinal dorsal horn of wild-type and NIPSNAP1−/− mice. Representative results (e) and number of p-ERK-positive neurons (f) in the spinal dorsal horn at 5 min after heat stimulation. p-ERK-positive neurons were quantified in four slices prepared from the three separated mice (mean ± SEM).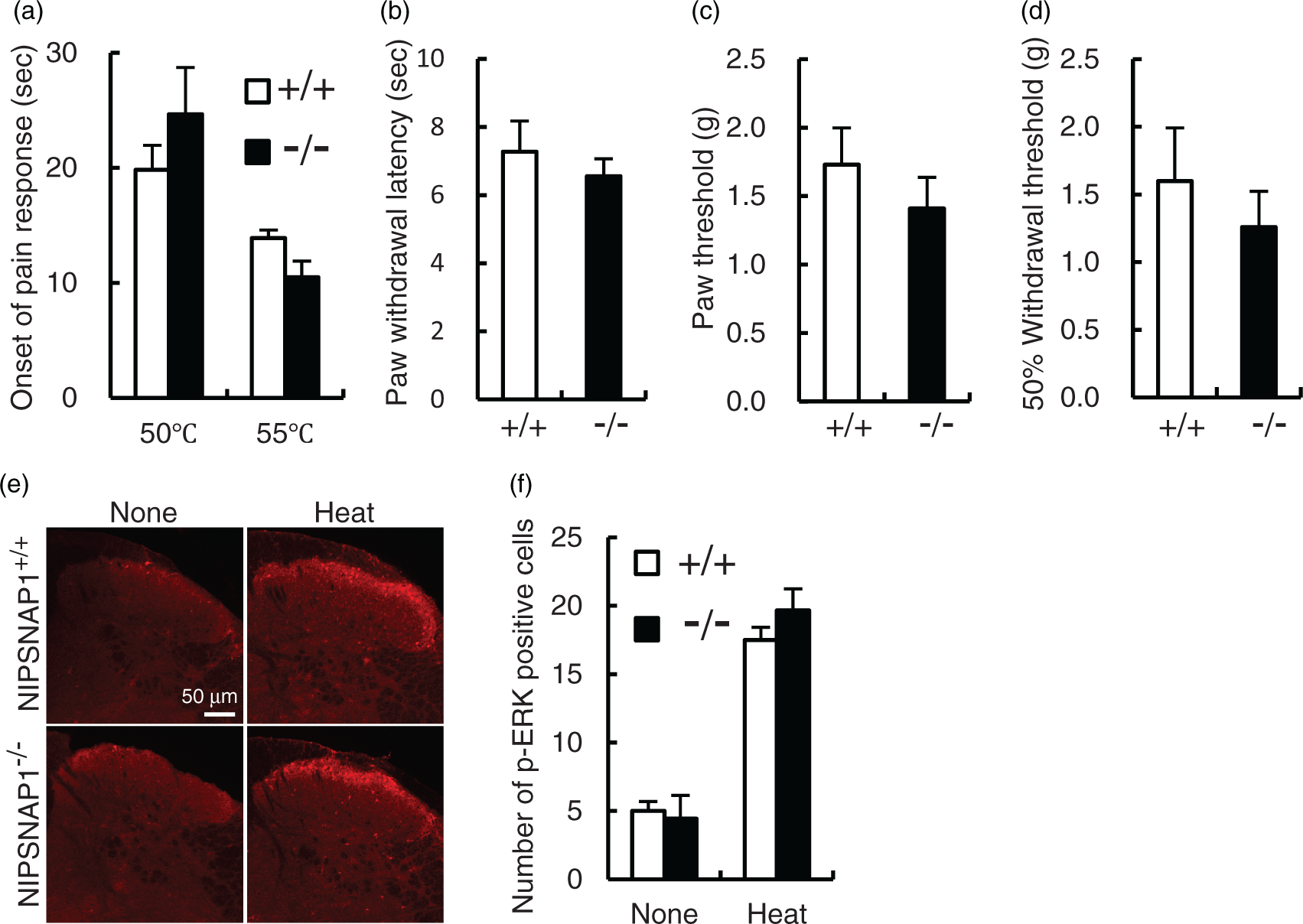
Induction of inflammatory pain in NIPSNAP1−/− mice
To determine the involvement of NIPSNAP1 in inflammatory pain, we subjected NIPSNAP1−/− mice and their wild-type littermates using three models as follows: formalin test and carrageenan- and CFA-induced inflammation. Formalin activates transient receptor potential A1 in sensory neurons to induce pain behaviors.
17
Formalin injection into the hind paw evokes the nociceptive behaviors of licking and biting the injected paw in a biphasic manner.18,19 The first transient phase (phase I) of the formalin response involves direct activation and acute sensitization of nociceptors, that is followed by a prolonged phase (phase II) induced by inflammatory responses mediated by the continuous activation of nociceptors and the central sensitization of neurons within the spinal dorsal horn triggered by noxious inputs. Wild-type and NIPSNAP1−/− mice exhibited biphasic pain responses in formalin test (Figure 2(a)). There were no significant differences in the intensity of the pain responses between wild-type and NIPSNAP1−/− mice during the phase I (0–15 min), as shown in Figure 2(b). The duration of pain behavior in the later (26–50 min, phase II-l), but not early (16–25 min, phase II-e), stage of phase II significantly increased in NIPSNAP1−/− mice (wild type, 295.4 ± 24.7 s vs. NIPSNAP1−/−, 459.9 ± 55.3 s, P < 0.05). Pain sensitization during the phase II response in the formalin test requires ERK activation in the spinal dorsal horn.15,16,20 To investigate the role of NIPSNAP1 in pain activity-dependent changes, we determined the level of p-ERK in the spinal cords of NIPSNAP1−/− and wild-type mice. The number of p-ERK-positive cells in the dorsal horn of spinal cord dramatically increased at 5 min after formalin injection into the dorsal surface of the hind paw and then decreased during the next 30–60 min in wild-type and NIPSNAP1−/− mice (Figure 2(c) and (d)). The number of p-ERK positive cells significantly increased at 5 min and 30 min in formalin-injected NIPSNAP1−/− mice (5 min, wild type 7.1 ± 1.1 cells vs. NIPSNAP1−/− 13.0 ± 2.4 cells, P < 0.05; 30 min, wild type 3.8 ± 0.4 cells vs. NIPSNAP1−/− 6.3 ± 0.6 cells, P < 0.01). These results indicate that NIPSNAP1 deficiency induced the aggravation of inflammatory pain through central sensitization elicited by the formalin test.
Pain sensitization induced in NIPSNAP1−/− mice by peripheral injection of formalin. Time spent licking and biting per 5 min is plotted versus time after peripheral injection of 2% formalin into the right hindpaw of wild-type and NIPSNAP1−/− mice. Data are expressed as the mean ± SEM (n = 11–13). **p < 0.01, *p < 0.05, versus wild-type value. (b) Cumulative time spent licking and biting during phase I (0–15 min), phase II-e (16–25 min), and phase II-l (26–50 min). *p < 0.05, versus wild-type value. (c) Formalin injected into the hind paw of wild-type and NIPSNAP1−/− mice induced ERK phosphorylation (red) in the ipsilateral dorsal horn of the spinal cord. Representative immunohistochemical data at the times indicated after formalin injection. (d) Number of p-ERK-positive neurons in the dorsal horn of spinal cord induced by formalin. p-ERK-positive neurons were quantified in four to eight slices prepared from the three to six separated mice (mean ± SEM). **p < 0.01, *p < 0.05, versus wild-type value.
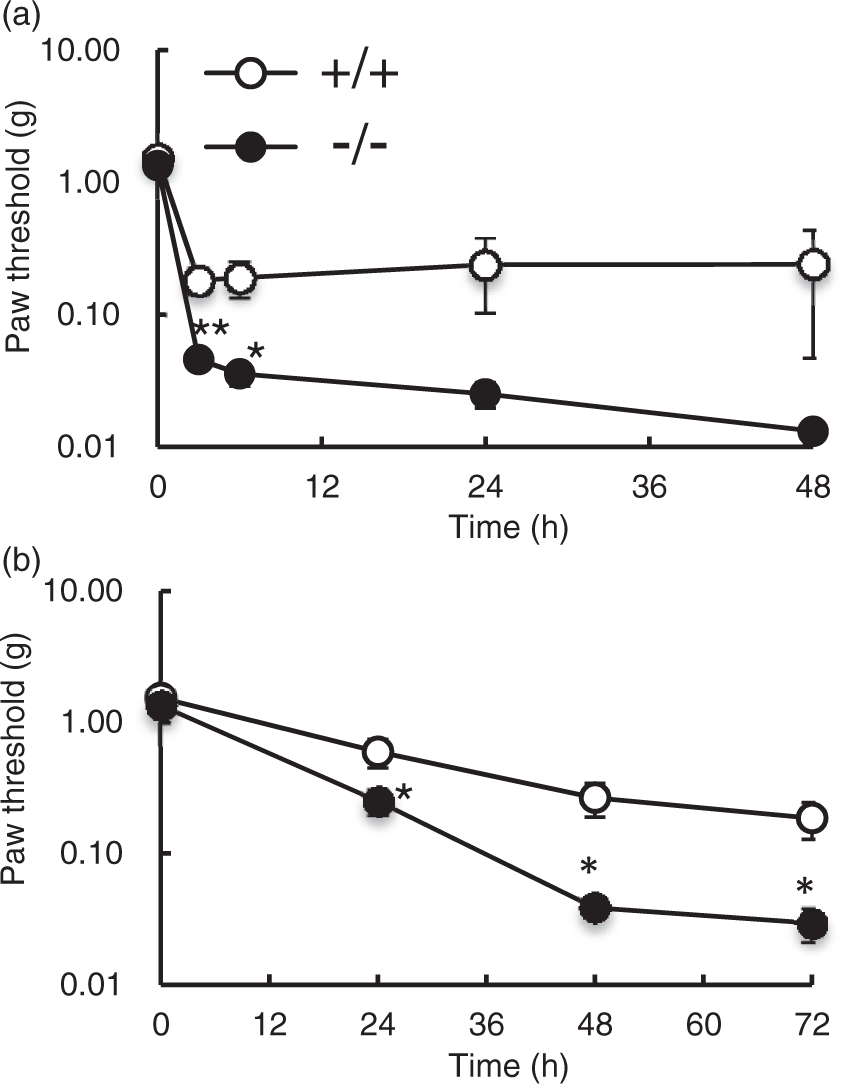
To further investigate the involvement of NIPSNAP1 in the induction of prolonged inflammatory pain, we analyzed hyperalgesia effects after injecting mice with 1% carrageenan and CFA. Intraplanter injection of carrageenan produces the paw edema, the infiltration of inflammatory cells such as macrophage and neutrophil, and the production of cytokines and chemokines. The diameter of paw at 5 h after carrageenan injection was 3.03 ± 0.22 mm and 2.98 ± 0.06 mm in wild-type and NIPSNAP1−/− mice, respectively, and these mice showed no significant difference in the paw edema (Figure 3(a)). We next compared the production of monocyte chemotactic protein-1 (MCP-1), a major pronociceptive chemokine, in the carrageenan-injected paw of wild-type and NIPSNAP1−/− mice. At 5 h after carrageenan injection, the MCP-1 contents of paw were increased from 62.3 ± 7.1 pmol/mg to 593.6 ± 26.4 pmol/mg in wild-type mice, and similar results were obtained in NIPSNAP1−/− mice from 54.7 ± 5.3 pmol/mg to 509.6 ± 33.3 pmol/mg (Figure 3(b)). Furthermore, the infiltration of inflammatory cells such as macrophage-specific marker F4/80-positive cells and neutrophil-specific marker Gr-1-positive cells were markedly increased in the carrageenan-injected paw of wild-type and NIPSNAP1−/− mice; however, these mice showed no significant difference (Figure 3(c) and (d)). These results indicated that the deficiency of NIPSNAP1 did not affect the carrageenan-induced inflammation at peripheral level. On the other hand, carrageenan-induced mechanical hyperalgesia applied by von Frey filaments during 3–48 h after the injection of NIPSNAP1−/− mice was exacerbated compared with wild-type mice and hyperalgesia significantly increased at 3 h and 6 h after carrageenan injection (3 h, wild type 0.18 ± 0.04 g vs. NIPSNAP1−/− 0.04 ± 0.01 g, P < 0.01; 6 h, wild type 0.19 ± 0.06 g vs. NIPSNAP1−/− 0.04 ± 0.01 g, P < 0.05; Figure 4(a)). The results for CFA-injected NIPSNAP1−/− mice were similar to that of carrageenan injection (Figure 4(b)). CFA-induced mechanical hyperalgesia applied by von Frey filaments was significantly enhanced in NIPSNAP1−/− mice compared with that in wild-type mice during 24–72 h after injection (24 h, wild type 0.59 ± 0.15 g vs. NIPSNAP1−/− 0.25 ± 0.05 g, P < 0.05; 48 h, wild type 0.27 ± 0.08 g vs. NIPSNAP1−/− 0.04 ± 0.01 g, P < 0.05; 72 h, wild type 0.19 ± 0.06 g vs. NIPSNAP1−/− 0.03 ± 0.01 g, P < 0.05). These results suggest that NIPSNAP1 deficiency induced mechanical hyperalgesia during prolonged inflammatory.
Effect of NIPSNAP1 deficiency on the carrageenan-induced inflammatory responses in the paws. In the paws at 5 h after the peripheral carrageenan injection of wild-type and NIPSNAP1−/− mice, the diameter (a), the level of MCP-1 (b), and the numbers of F4/80-positive macrophages, and Gr-1-positive neutrophils (c, d) were measured. Representative results (c) and number of F4/80 - and Gr-1-positive cells were quantified in two slices prepared from the three separated mice (d). Data are expressed as the mean ± SEM (a, n = 8; b, n = 4; c, n = 6). Pain sensitization induced by peripheral injection of NIPSNAP1−/− mice with carrageenan and CFA. Paw withdrawal latencies of mice injected with 1% carrageenan (a) or CFA (b) using von Frey filaments. Data are expressed as the mean ± SEM (n = 7). **p < 0.01, *p < 0.05, versus wild-type value.
Analysis of NIPSNAP1 expression following the induction of inflammatory pain
NIPSNAP1 is expressed in the brain, spinal cord, liver, and kidneys.1–3 RT-PCR analysis detected NIPSNAP1 mRNA in the spinal cord and DRG but not in a peripheral tissue paw (Figure 5(a)). In situ hybridization analysis mainly detected NIPSNAP1 mRNA in the large motor neurons of the ventral spinal cord (Figure 5(b)). NIPSNAP1 mRNA was expressed from small- to medium-sized DRG cells (240–780 µm2). In contrast, NIPSNAP1 mRNA was not detected in the spinal cord or DRG of NIPSNAP1−/− mice.
Distribution of NIPSNAP1 mRNA. (a) RT-PCR analysis of NIPSNAP1 mRNA expression in the spinal cord, paw, and DRG. GAPDH served as the control. (b) In situ hybridization analysis of NIPSNAP1 mRNA expression in the spinal cords and dorsal root ganglia of wild-type and NIPSNAP1−/− mice.
Because NIPSNAP1-deficiency affected inflammatory pain, we next analyzed NIPSNAP1 mRNA expression during the inflammatory response. In the formalin test, NIPSNAP1 mRNA level increased from 5 min to 60 min after formalin injection in DRG and 1.7-fold increase at 60 min compared with that in the control (Figure 6(a)). In contrast, NIPSNAP1 mRNA level was significantly decreased from 24 h to 48 h after carrageenan injection of DRG (24 h, 0.60 ± 0.09-fold, p < 0.01; 28 h, 0.67 ± 0.11-fold, p < 0.05; Figure 6(b)). Significant changes of NIPSNAP1 mRNA level were not observed in the spinal cord after formalin or carrageenan injection.
Analysis of NIPSNAP1 mRNA levels following the induction of inflammation. Levels of NIPSNAP1 mRNA in DRG and spinal cord induced by injection into the dorsal surface of hind paw of 2% formalin (a) or 1% carrageenan (n). The upper and lower panels show representative results of RT-PCR and real-time PCR analysis, respectively. Data are expressed as the mean ± SEM (n = 3). **p < 0.01, *p < 0.05, injected versus controls.
Oral administration of aspirin (300 µg/g), an inhibitor of PG synthesis, completely inhibited nociceptive responses induced by formalin in NIPSNAP1−/− and wild-type mice (data not shown). Intraperitoneal and i.t. administrations of cyclooxygenase inhibitor ibuprofen reduced the phase II response of formalin test and spinal release of PGE2.
21
Further, carrageenan administration induced PGE2 synthesis in paw and spinal cord.
22
Therefore, we investigated whether PGE2, a key mediator of pain transmission, regulated NIPSNAP1 mRNA expression in isolated DRG cells. PGE2 (5 µM) significantly increased 1.2-fold NIPSNAP1 mRNA level at 1 h and returned to the basal level at 6 h (Figure 7(a)). Decreased NIPSNAP1 mRNA expression was not detected in DRG cells treated with PGE2. RT-PCR analysis revealed that DRG cells expressed mRNAs encoding the PGE2 receptor subtypes, EP1, EP2, EP3α, EP3β, EP3γ, and EP4 (Figure 7(b)). Specific agonists of EP2 (ONO-AE1-259), EP3 (ONO-AE-248), and EP4 (ONO-AE1-329) but not of an EP1-specific agonist (ONO-DI-004), increased NIPSNAP1 expression at 1 h in DRG cells. The increases induced by EP2, EP3, and EP4 agonists were statistically significant (EP2, 1.74 ± 0.01-fold, p < 0.01; EP3, 1.77 ± 0.24-fold, p < 0.05; EP4, 1.78 ± 0.24-fold, p < 0.05; Figure 7(c)). The protein kinase A (PKA) inhibitor H-89 (20 µM) inhibited the PGE2-induced increase in NIPSNAP1 mRNA level in the DRG cells (Figure 7(d)). Although EP2 and EP4 receptors couple to Gs proteins to increase cAMP levels,
23
EP3α and EP3β receptors are coupled to Gi proteins followed by inhibition of cAMP synthesis, and EP3γ receptor is coupled to a Gs protein.24,25 H-89 also decreased the level of NIPSNAP1 mRNA expression induced by EP3 agonist in the DRG cells, with the similar inhibition level of EP4 agonist (Supplementary Figure 1). These results demonstrate that increased NIPSNAP1 mRNA expression was mediated through the cAMP-PKA signaling pathway.
Analysis of NIPSNAP1 mRNA expression induced by PGE2. (a) RT-PCR and real-time PCR analysis of NIPSNAP1 expression induced by 5 µM PGE2 in DRG cells. (b) RT-PCR analysis of the expression of mRNA encoding EP subtypes in DRG cells. White arrowhead shows EP3β. (c) Effect of EP agonists on the expression of NIPSNAP1 mRNA. DRG cells were incubated for 1 h in the presence of each EP agonist (1 µM), and subjected to RT-PCR and real-time PCR analysis. (d) Effect of the protein kinase A inhibitor H-89 on the PGE2-induced expression of NIPSNAP1 mRNA. DRG cells incubated with 5 µM PGE2 for 1 h in the absence and presence of 20 µM H-89 were subjected to RT-PCR and real-time PCR analysis. The upper and lower panels show representative results of RT-PCR and real-time PCR analysis, respectively. Data are expressed as the mean ± SEM (n = 3–4). **p < 0.01, *p < 0.05, treated versus untreated cells. ##p < 0.01, treated versus PGE2-treated cells.
Finally, we examined whether PGE2 affected on the expression of NIPSNAP1 mRNA following the inflammatory pain using a cyclooxygenase inhibitor aspirin. When aspirin (300 µg/g) was administrated orally at 1 h before the formalin injection, the formalin-induced increase of NIPSNAP1 mRNA in the DRG was reduced (Figure 8). This result suggests that the increase of NIPNSAP1 mRNA was mediated by PG, probably PGE2, in the inflammatory pain model induced by formalin.
Effect of cyclooxygenase inhibitor on the formalin-induced NIPSNAP1 mRNA increase in DRG. Aspirin (300 µg/g) was administered orally to mice for 1 h before the peripheral formalin injection. Data are expressed as the mean ± SEM (n = 3–6). *p < 0.05, versus vehicle-administered mice.
Discussion
In this study, we provided novel evidence that NIPSNAP1 modulates pathological inflammatory pain. Specifically, we demonstrated that peripheral inflammation elicited by formalin was significantly enhanced during central sensitization to pain in spinal dorsal horn of NIPSNAP1−/− mice compared with that of wild-type mice. Moreover, carrageenan and CFA exacerbated the prolonged pain responses of NIPSNAP1−/− mice. NIPSNAP1 mRNA levels expressed in DRG significantly increased at 60 min in formalin-injected mice and decreased from 24 h to 48 h in the carrageenan-injected mice. Further, the major mediator of inflammation PGE2 increased NIPSNAP1 mRNA levels in DRG cells through the cAMP-PKA pathway.
The duration of the response during the later stage of the formalin test (26–50 min) of NIPSNAP1−/− mice was significantly longer compared with that of wild-type mice, although the responses of either were similar during phase I (0–15 min) and the earlier stage (16–25 min) of the phase II (Figure 2(a) and (b)). We found that formalin injection significantly increased the number of p-ERK positive cells in the lumbar spinal cord of NIPSNAP1−/− mice compared with that in wild-type mice (Figure 2(c) and (d)). Deficiency of NIPSNAP1 induced increased p-ERK positive cells of dorsal spinal cord occurred by 5–30 min post-formalin injection and significant aggravation of pain behavioral response by 31–40 min (Figure 2(a)). Inhibitors of ERK-mediated signaling reduce the pain behavioral response in phase II without significant effect on the phase I of the formalin test, suggesting that nociceptive sensitization in phase II is dependent on ERK activation.15,16,20 It has been reported that i.t. administration of PD98059, an ERK inhibitor, inhibits the pain responses of phase II with maximal inhibitory effect at the later stage (30–40 min), although the level of ERK phosphorylation peaked at 3 min in the superficial spinal cord. 15 In addition, the deficiency of ERK2 reduces the 5-min ERK phosphorylation and the pain responses of phase II in formalin test. 20 Taken together, we conclude from these findings that NIPSNAP1 implicates in modulating central pain sensitization evoked by formalin through ERK activation.
I.t. administration of cyclooxygenase inhibitor reduces pain behavioral response in phase II of formalin test, and peripheral formalin evokes the increase of PGE2 release from the spinal cord in the 0–10 min and 20–30 min. 21 Treatment with PGE2 increased NIPSNAP1 mRNA levels at 1 h in primary DRG cells (Figure 7(a)), and NIPSNAP1 mRNA expression was significantly upregulated by formalin injection at 60 min in DRG (Figure 6(a)). Peripheral or spinal PGE2 generated by formalin injection probably stimulate NIPSNAP1 mRNA expression. In fact, administration of cyclooxygenase inhibitor inhibited the increase of NIPSNAP1 mRNA expression induced by formalin (Figure 8). In contrast, the changes of NIPSNAP1 mRNA expression were not observed in spinal cord (Figure 6), which may be due to the localization of NIPSNAP1 mRNA in motor neurons of ventral spinal cord (Figure 5(b)). The upregulation of NIPSNAP1 mRNA expression observed at 60 min was event after ERK activation and aggravation of pain behavioral response. Therefore, the increased NIPSNAP1 mRNA expression is likely involved in the attenuation of inflammatory pain.
We demonstrated that NIPSNAP1 influenced the duration of inflammatory pain. Prolonged nociceptive behaviors induced by carrageenan and CFA were exacerbated in NIPSNAP1−/− mice (Figure 4), and NIPSNAP1 mRNA expression in DRG was reduced at 24–48 h after administration of carrageenan (Figure 6(b)). Although intracerebroventricular or i.t. administrations of NST blocks the pain behavior induced by peripheral injection of carrageenan,8,11 the mRNA encoding the NST precursor preproN/OFQ mRNA is hardly expressed in DRG neurons under physiological conditions, transiently increases during carrageenan-induced peripheral inflammation, peaks at 30 min later, and returns to basal level within 6 h.26,27 Therefore, decreased NIPSNAP1 expression is likely involved in the exacerbation of prolonged inflammatory pain independent of NST generation. Furthermore, carrageenan reduced the NIPSNAP1 mRNA expression, although the peripheral carrageenan-induced PGE2 release is observed at 1–6 h and 12–24 h in the paw and 3–24 h in the spinal cord. 22 It seems likely that the carrageenan-induced PGE2 release is later than the formalin-induced one. The unknown reduction mechanism of NIPSANP1 mRNA expression may greatly exceed the increase of NIPSNAP1 mRNA induced by PGE2 in the later inflammatory pain, resulting in decreased the NIPSNAP1 mRNA induced by peripheral carrageenan injection.
PGE2 exerts its effects through four different G-protein-coupled EP receptors. We demonstrated that NIPSANP1 mRNA levels were significantly increased by EP2, EP3 (probably EP3γ), and EP4 through the activation of adenylate cyclase and subsequent activation of PKA. Moreover, the PKA inhibitor H-89 inhibited NIPSNAP1 mRNA expression induced by PGE2 in DRG cells (Figure 7(d)). PKA phosphorylates the transcription factor cAMP response element-binding protein. Analysis of transcription factor bindings sites available in the TFBIND database 28 revealed at least eight cAMP response element-binding protein binding sites in 1-kb upstream of mouse NIPSNAP1 gene (data not shown). Taken together, the data support the conclusion that PGE2 increased NIPSNAP1 mRNA expression through the cAMP-PKA pathway, although the mechanisms that downregulates NIPSNAP1 expression remains to be determined.
Conclusions
In conclusion, NIPSNAP1 contributes to the pathogenesis of inflammatory pain by regulating central sensitization via ERK activation. The upregulation of NIPSNAP1 mRNA via PGE2 correlates with the decreased central sensitization during convalescence, as well as the downregulation of NIPSNAP1 mRNA expression the exacerbation of prolonged behavioral hypersensitivities. Thus, our findings suggest that NIPSNAP1 will serve as a novel therapeutic target for pathological inflammatory pain.
Footnotes
Author note
Kazuya Okamoto is currently at Nomura Micro Science Co. Ltd., Atsugi, Japan; Masaki Ohashi is currently at Oisis Corporation, Itami, Japan; and Kyohei Fujisato is currently at Yao Tokushukai General Hospital, Yao, Japan.
Author contributions
KOk conducted most experiments, analyzed data, and produced the figure. MO, KOh, EM, and KF participated in the experiments using DRG cells. KOh and AT participated in the experiments using mouse pain model. TM supervised behavioral analysis of pain models. SI supervised the project and edited the manuscript. EO-A participated in the design of experiments and manuscript preparation. All authors read and approved the final manuscript.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by Grants-in-Aid for Scientific Research (C) from the Japan Society for the Promotion of Science to E O-A (Grant Numbers 23590726, 26460709).